高寒地区不同退化草地植被和土壤微生物特性及其相关性研究
2015-04-15卢虎姚拓李建宏马文彬柴晓虹
卢虎,姚拓,李建宏,马文彬,柴晓虹
(甘肃农业大学草业学院,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
高寒地区不同退化草地植被和土壤微生物特性及其相关性研究
卢虎,姚拓*,李建宏,马文彬,柴晓虹
(甘肃农业大学草业学院,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
以青藏高原东北缘天祝高寒地区不同退化草地为研究对象,通过野外调查及室内测定分析,研究了植被特征、微生物量特性、微生物熵及三者之间的相关性。结果表明,随着草地退化程度的加重,植被种类减少、优势种改变,植物群落高度、盖度、地上生物量显著降低(P<0.05),土壤微生物生物量碳、氮减少。草地植被地上生物量从轻度退化草地(LDG)到重度退化草地(SDG)减少了88.63%;LDG中微生物量碳、氮(218.90 mg/kg、44.32 mg/kg)均显著高于SDG(P<0.05),但中度退化草地(MDG)与SDG差异不显著;微生物熵在不同退化草地中的变化规律不明显。草地地上生物量与土壤微生物量呈显著正相关关系(P<0.05),与微生物熵呈负相关关系,其中微生物熵与微生物量碳的相关系数相对较高。
植被特性;微生物量碳;微生物量氮;微生物熵
天然草地在干旱、风沙、水蚀、盐碱、内涝、地下水位变化等诸多不利的自然因素影响下,或过度放牧、滥挖、滥割和樵采等不合理人为管理和利用下,植被遭到破坏,导致了草地生态环境恶化,草地利用性能降低,甚至失去利用价值。苏大学等[1]提出广义的草地退化概念包括草地退化、沙化和盐渍化等3个方面。健康草地生态系统具有极高的生态价值。谢高地等[2]估算草地生态系统对青藏高原生态系统总服务价值的贡献最大为48.3%。天然草地生态系统每年提供的生态服务价值为2571.78×108元,占全国草地生态系统每年服务价值的17.68%[3]。朴世龙和方精云[4]认为青藏高原草地植被的总NPP为0.21 Pg C (1 Pg=1015g),约占全国植被NPP总量的12.43%。闫玉春和唐海萍[5]指出草地退化是草地生态学和草地经营学中倍受关注的问题。陈全功[6]认为气候变异是江河源区高寒草甸退化的主要原因,不合理的草地利用加剧了黑土滩的蔓延和危害。高清竹等[7]利用遥感监测和评价指标体系研究发现从1981年到2004年的近24年里,藏北地区及其各个区域草地退化较为严重。刘纪远等[8]通过三期遥感影像研究发现,三江源草地退化的格局在20世纪70年代中后期已基本形成,且至今草地的退化过程一直在持续发生。上述研究从宏观角度分析草地退化的过程,涉及微观角度的研究主要集中在土壤微生物的活性方面。
土壤中生活着丰富的微生物类群,其种类、数量、分布、生命活动规律与土壤中的物质和能量转化、土壤肥力、植物生长等的关系是土壤微生物资源的价值所在。章家恩和刘文高[9]认为微生物资源与农业可持续发展的关系十分密切,在提高与保持土壤肥力、转化营养元素、净化环境与平衡生态系统等方面起着极其重要的作用。陈秀蓉和南志标[10]指出细菌在农业生态系统中不仅具有调节植物生长发育、抑制病原微生物等作用,而且在生态系统中的营养元素矿化、土壤肥力的保持和提高以及能量转化和物质循环等方面具有其他生物无法代替的作用。近年来的研究报道中,土壤微生物的作用除了生态系统的“分解者”,还增加了系统中能量、物质、信息流的链接角色。李骁和王迎春[11]阐述植物、土壤和微生物通过相互作用,构成了一个植物-土壤-微生物的有机整体。土壤微生物多样性代表着微生物群落的稳定性,是生命体在遗传、种类和生态系统层次上的变化。王新平等[12]结合土壤分形特征、植被盖度、物种特征、生物量、土壤水分、土壤微生物,以及土壤物理和土壤养分特征分析了人工固沙灌木林的演变。王晓龙等[13]系统调查了鄱阳湖典型湿地植物群落土壤微生物量,地表植被生物量及土壤养分性状。宗宁等[14]在西藏高寒草甸地区开展了外源氮素添加与刈割模拟放牧实验,测定了其对植物生物量分配、土壤微生物碳氮和生态系统呼吸的影响。目前,有关植被、土壤以及微生物三方面的单独研究已较为深入,然而综合分析比对草地生态各要素之间关系的报道仍较少,本研究选择青藏高原东北缘天祝高寒地区不同退化草地为对象,运用系统学的观点将草地植被、土壤微生物的重要指标综合分析,旨在为青藏高原草地健康及持续利用提供理论依据和技术支撑。
1 材料与方法
1.1 研究区自然概况
研究地位于甘肃省天祝县金强河甘肃农业大学高山草原试验站(N 37°11′-37°13′,E 102°29′-102°33′),海拔2960~2990 m,气候寒冷潮湿,年均温-0.1℃,1月均温-18.3℃,7月均温12.7℃,>0℃年积温1380℃,水热同期,年日照时数2600 h;年降水量416 mm,多为地形雨,集中于7-9月,年蒸发量1592 mm。无绝对无霜期,仅分冷、热2季,春季常干旱,并有暴风雪。植被以嵩草(Kobresiabellardii)、苔草(Carexspp.)、针茅(Stipacapillata)、莎草(Cyperusspp.)、珠芽蓼(Polygonumviviparum)、金露梅(Potentillafruticosa)、棘豆(Oxytropisspp.)、狼毒(Stellerachamaejasme)、委陵菜(Potentillachinensis)和杜鹃(Rhododendronspp.)等为主。土层厚40~80 cm。土壤pH值7.0~8.2,土壤水分40%~80%,有机质含量10%~16%,全氮含量0.5%~0.8%,全磷含量0.056%~0.071%。土壤以亚高山草甸土、亚高山黑钙土等为主。
1.2 样地植物特性调查
根据《天然草地退化、沙化、盐渍化的分级标准》(GB19377-2003)[1],将研究区内草地分为轻度退化草地(lightly degraded grassland, LDG)、中度退化草地(moderate degraded grassland, MDG)和重度退化草地(severe degraded grassland, SDG)(表1),于2013年7月进行采样。在每种退化草地上随机选取 3个50 cm×50 cm的样方,调查并记录每个样方内的植物种类、高度和盖度。植被盖度采用针刺法测量,高度采用样方内群落自然高度的平均值。同时将样方内的植物齐地面刈割,除去粘附的土壤、砾石等杂质后带回实验室,在(100±5)℃恒温箱内烘干至恒重,测定样方内地上生物量[15]。

表1 样地基本概况
1.3 样地土壤样品采集
于2013年7月,在1.2的刈割了地上植物的每个样方内,除去地面的枯枝落叶及砾石等杂质,按0~10 cm、10~20 cm和20~30 cm三个层面分别采集土壤样品,每个样方取1 kg左右装入无菌聚乙烯袋密封,放置在盛有冰袋的整理箱中低温运输。土壤样品带回实验室后立即进行土壤微生物生物量碳、氮及微生物熵的测定(4℃不超过24 h)。
1.4 土壤微生物特性测定
1.4.1 微生物量碳测定 微生物量碳采用氯仿熏蒸浸提法[16]。按以下公式计算土壤微生物量碳。
式中,N为FeSO4浓度;Vo为空白液消耗FeSO4(mL)数;V为土壤提取液消耗FeSO4(mL)数;F为稀释倍数;M为烘干土样重(g)。
微生物量碳=0.38×ΔEC
式中,ΔEC为熏蒸和不熏蒸0.5mol/L的K2SO4提取的碳的差值。0.38为校正系数。
1.4.2 微生物量氮的测定 微生物量氮采用氯仿熏蒸浸提法[17]。按以下公式计算土壤微生物量氮。
式中,N为HCl的当量浓度;V为土壤浸提液消耗HCl(mL)数;Vo为空白液消耗HCl(mL)数;0.014为每mg当量氮的重量(g);F为稀释倍数;M为烘干土样重(g)。
微生物量氮=0.54×ΔEN
式中,ΔEN为熏蒸和不熏蒸0.5mol/LK2SO4提取的氮的差值。0.54为校正系数。
1.4.3 微生物熵的测定 土壤微生物熵(qCO2)用碱液吸收法测定[14]。通过以下公式计算:
式中,EC为微生物量;Vo为空白液消耗HCl(mL)数;V为土壤培养24h后含CO2碱液消耗HCl(mL)数;M为烘干土样重(g)。
1.5 数据分析
采用Excel2007和SPSS17.0软件进行数据整理,运用Duncan方差分析方法及Pearson相关性分析方法分别进行统计分析。
2 结果与分析
2.1 不同退化草地植被特征
2.1.1 草地植物种类 调查表明(表2),不同退化程度草地内植物种类的组成各不相同,数量也有较大改变。植物种类以轻度退化草地(LDG)最为丰富,其次为中度退化草地(MDG),重度退化草地(SDG)最少。随着退化程度的加重,植物种类数量锐减,这是因为当草原急剧退化时,土壤质量、含水量及孔隙度等物理性质发生改变,使得原来定植的某些物种不再适宜当地的土壤环境,还有某些物种对其他植物的他感作用,如草原退化的指标物种狼毒,上述原因都导致植物种类数量减少。

表2 草地植被种类
3种退化草地(LDG、MDG和SDG)植物优势种分别为披碱草-扁蓿豆、披碱草-嵩草和披碱草-委陵菜。披碱草在3种退化草地中皆为优势种,证明其受草地退化程度影响较小,相比其他植物,披碱草耐寒抗旱、耐盐碱抗风沙的生态习性突出,适宜高寒草原的生长环境,物种频度高。轻度退化草地中的另一优势种扁蓿豆主要分布在我国高纬度寒冷地区,性耐寒、抗旱能力也较强。中度退化草地中的嵩草生长在2500~3400 m高海拔地区,适应寒冷潮湿气候,具有一定的抗旱能力,其须根相当发达,与其他莎草科植物根系交织一起,形成富有弹性的生草土,因此具有耐牧、耐践踏特性,当草地出现退化时,嵩草成为草地中的主要植物种类。重度退化草地中委陵菜是一种生境分布极广的多年生草本植物,在海拔400~3200 m的地区都可以生长,其种类繁多,耐践踏,且适口性差,家畜不喜采食,草地严重退化之后,委陵菜仍然可以生长。
2.1.2 草地群落高度和盖度 草地植物群落是各种群之间通过竞争、共生等种间关系而形成的一个具有一定群落结构的集合体。调查表明(表3),不同退化程度,草地植被高度和盖度随退化程度的加重而递减,其中植物群落高度从轻度退化草地(LDG)到中度退化草地(MDG)减少了48.84%,从中度退化草地(MDG)到重度退化草地(SDG)减少了90.91%;群落高度的大幅度减少在一定程度上表明了草地植被群落垂直空间结构的缩减,退化越严重,群落结构越简单,其直接原因是放牧压力的增加、家畜的啃食及践踏强度加深。植被群落盖度从LDG到MDG减少了13.54%,从MDG到SDG减少了7.23%;群落盖度的降低是在草地植被群落水平空间结构层面的改变,其改变没有垂直空间那么剧烈,主要原因是放牧干扰对植物种类在水平格局上的分布影响不大。
2.1.3 不同退化草地地上生物量 草地地上生物量与群落高度及盖度变化一致,随退化程度的加深而数量递减。从轻度退化草地(LDG)到中度退化草地(MDG)减少了54.02%,从中度退化草地(MDG)到重度退化草地(SDG)减少了75.00%,从轻度退化草地(LDG)到中度退化草地(SDG)减少了88.63%(表3)。在退化程度不断加重的同时,植物种类减少、群落空间结构缩小,相应的植物量也急剧减少,作为草地农业生态系统重要的产量指标,单位面积与体积内所含植物体的总量整体下滑,故此草地退化与地上生物量之间具有明显的对应关系,地上生物量的大幅度降低直接反映了草地退化程度在不断加重。

表3 不同退化草地植被特征
注:同列不同字母表示差异显著(P<0.05),下同。
Note: The different letters within the same column mean significantly different (P<0.05), the same below.
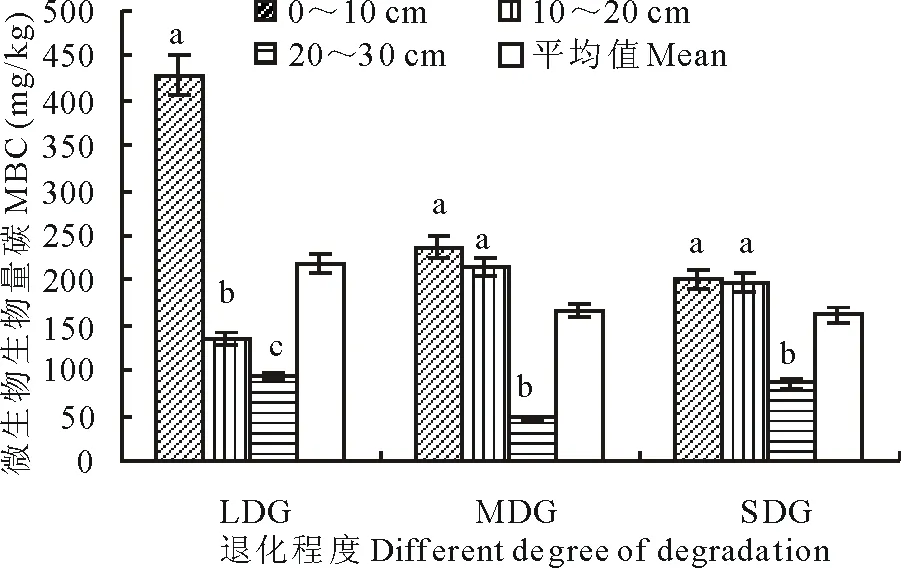
图1 不同退化草地土壤微生物量碳含量Fig.1 Content of microbial biomass carbon (MBC) in different degraded grasslands不同字母表示差异显著(P<0.05),下同 。 The different letters mean significantly different (P<0.05), the same below.
2.2 不同退化草地土壤微生物特征
2.2.1 微生物量碳 土壤微生物量碳参与了土壤以及大气的碳循环,是微生物生态中最重要的指标之一。结果表明(图1),不同退化程度的草地土壤微生物量碳随退化程度加重而减少。其中轻度退化草地(LDG)显著高于中度退化草地(MDG)(P<0.05),但是MDG和重度退化草地(SDG)中微生物量碳的差异不显著。草地退化对土壤微生物量碳的影响非常大,这是因为草地退化主要表现在植被退化上,其种类组成、空间结构以及产草量等多方面的退化引起了土壤微生物生态的变化,微生物数量、种类和微生物量首先受到影响,呈下降趋势。微生物量碳是土壤活性成分的重要生理指标,土壤有机质的重要组成部分,它的改变直接反映了微生物生理生化活性及土壤肥力的大小。草地退化之后,土壤微生物生理生化活性降低、土壤肥力降低,可提供的有机质减少,则土壤微生物所占的生物量碳的比例就随之减少。
土壤微生物量碳上层高于下层,最大值都出现在0~10 cm土层,LDG中最为明显;MDG中0~10 cm和10~20 cm土层之间差异不显著,但都和深层20~30 cm差异显著;SDG和MDG呈现出相似的特征(图1)。土壤微生物具有一定的垂直分布规律,因为0~10 cm土层植被根系发达、土壤透气性较高,这一土层为微生物提供了生长空间与绝大部分的营养物质,相比较深的土层,这一土层微生物大量繁殖,活性最高。然而随着土壤层次的加深,植物根系分布减少、土壤透气性降低,这都不利于微生物通过吸收植物营养或固定大气中的CO2来维持其生命活动所需。
2.2.2 微生物量氮 土壤微生物量氮是土壤氮素的一个重要储备库,也是土壤有机氮中最活跃的组分,在土壤氮循环与转化过程中起着重要的调节作用。不同退化程度的草地土壤微生物量氮与微生物量碳一致(图2),即随着退化程度加重而递减,轻度退化草地(LDG)显著高于重度退化草地(SDG)(P<0.05)。土壤微生物量氮相比微生物量碳数值要小很多,但其数值随着草地退化而减少的规律仍然与微生物量碳一致。这是因为草地植被大幅度退化之后,土壤中的营养结构出现较大改变,土壤养分储量指标中重要的土壤氮素以及可利用土壤氮随之下降,土壤微生物对土壤氮素的转化及利用降低,微生物量氮自然呈下降趋势。
土壤微生物量氮在不同土壤层次中的分布与微生物量碳类似,不同退化程度中表层0~10 cm与其余两层差异显著,呈上层高于下层的规律,且在LDG、SDG中变化最明显(图2)。与上述土壤微生物量碳的原因一致,土壤微生物的垂直分布规律使得表层土壤比深层土壤更适宜于微生物的生长繁殖,该层次微生物生理活性也越高。在土壤氮素循环中,地壳表面广泛分布着具固氮作用的微生物和由微生物与植物所组成的各种类型的固氮体系,形成庞大的生物固氮网。具有固氮作用的微生物大量分布在土壤表层或是与植物组成根瘤菌分布在根际,由此决定了土壤微生物量氮必定呈随土壤层次加深而降低的趋势。
2.3 不同退化草地微生物熵特征
微生物熵是指一定时期内土壤微生物量碳与土壤呼吸作用产生的CO2-C的比值。实际上是单位数量的土壤微生物的呼吸强度。不同退化程度的草地微生物熵,最大值出现在中度退化草地(MDG)中,相比其他两种样地差异不显著(P>0.05)。微生物熵值作为一个比值并没有随着草地退化程度的改变而改变,这主要是因为影响微生物熵值变化的两个因素:土壤微生物量碳和土壤呼吸作用所产生的CO2-C这两个数值都在不断地发生着变化。轻度退化草地(LDG)由于植被退化程度较轻,土壤微生物活性较高、微生物量碳含量高,但土壤呼吸作用产生的CO2-C也因植物根系发达、土壤中小型动物的多样性丰富而数值较高,则微生物熵这个比值较低。MDG中植被退化改变了土壤中微生物的活性,微生物量碳有所降低,但由于能够进行呼吸作用的植物、动物、微生物的大量减少,其呼吸作用产生的CO2-C降低速率要比微生物量碳快,则微生物熵数值偏高。重度退化草地(SDG)中植被严重退化导致土壤微生物活性和土壤呼吸作用都大幅度降低,则微生物熵值也不比MDG低。
微生物熵值在土壤层次方面与土壤微生物量相反,最大值出现在20~30 cm,下层高于上层(图3)。这是因为随着土壤层次的加深,土壤微生物的活性降低、土壤中碳储存含量下降,微生物量碳不断减少,但是土壤呼吸作用在深层几乎无法进行或是极难进行,从表层到深层其下降幅度比微生物量的下降幅度要快,微生物熵是微生物量碳与呼吸作用产生的CO2-C的比值,则土壤深层的微生物熵比土壤表层的大。

图2 不同退化草地土壤微生物量氮含量Fig.2 Content of microbial biomass nitrogen (MBN) in different degraded grasslands
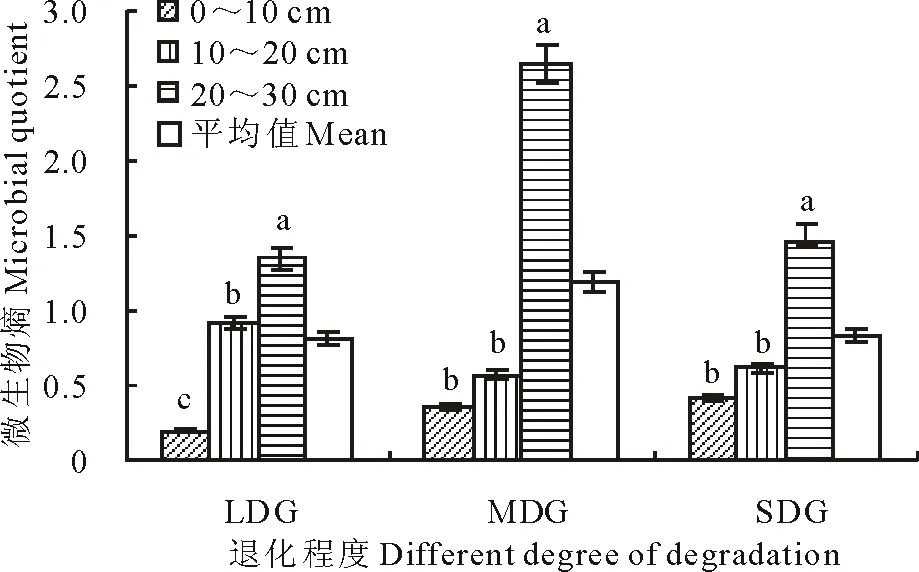
图3 不同退化草地的微生物熵Fig.3 Microbial quotient in different degraded grasslands
2.4 不同退化草地地上生物量、土壤微生物量及微生物熵之间的相关性
相关性分析表明(表4),不同退化程度的草地地上生物量与土壤微生物量碳、微生物量氮之间都呈显著正相关(P<0.05),与微生物熵呈负相关,且地上生物量与微生物量碳呈极显著正相关(P<0.01)。这是因为退化草地生态系统中,土壤活性成分微生物主要依靠植物供给营养物质,进而提供生长场所,植被退化之后,植物种类、数量各方面的改变导致微生物相应改变。地上生物量的变化直接与土壤微生物量相互关联,两者在变化上保持着高度一致。同时植被退化影响了土壤呼吸作用,地上生物量与微生物熵之间存在一定的关联,但与微生物量之间相关性小。
土壤微生物量碳、氮分别与地上生物量的相关系数为0.938和0.694,并且均达到显著正相关水平(P<0.05)。微生物量碳作为微生物的有机质,其来源主要就是地上植被生物量。微生物氮作为土壤氮素营养的重要组成部分,生物固氮等途径还依靠与植物根系的共生作用完成,所以它与地上植物量也关联较高。土壤微生物活性的改变也影响了土壤呼吸作用,则微生物量碳、氮与微生物熵三者之间也存在着不同大小的关联。
微生物熵与地上生物量及土壤微生物量都呈负相关,但是相关系数的显著性均未达到显著水平(P>0.05),其中与微生物量碳相关系数(-0.439)最高。土壤呼吸作用释放CO2量较小,且在一定程度上受土壤通透性等理化性质制约,则微生物熵与地上生物量、微生物量三者之间的关联较小,其显著性都未达到5%水平。土壤呼吸作用较大程度上是由土壤微生物完成的,土壤微生物活性的高低直接导致了呼吸作用的大小。其中微生物量碳作为微生物熵数值计算的分子成分,两者之间关联相比微生物量氮大。

表4 草地地上生物量、土壤微生物量及微生物熵之间的相关性
注:用Pearson法计算数值间相关性,**表示相关系数显著性达到1%水平,*表示相关系数显著性达到5%水平。
Note: ** indicate correlation is significant at the 0.01 level (2-tailed), * at the 0.05 level (1-tailed).
总体来看,由于植被情况与土壤微生物活性之间较强的对应关系,两者相关系数最高,达到显著水平(P<0.05)。土壤呼吸作用主要参与土壤碳循环过程中,微生物熵与地上生物量、微生物量碳相关系数相对较高,与参与土壤氮循环的微生物量氮相关系数相对较低,均未达到显著水平(P>0.05)。
3 讨论
3.1 草地退化对植被的影响
本研究通过实测得出,研究区的植被种类、优势种、植被群落高度及盖度、地上生物量随着草地退化程度的加重而不断减少的结果是草地退化在植被退化层面的具体表现形式,也是草地退化产生的直接结果,甚至在草地退化分级标准中经常使用。本研究选取的青藏高原草地属于高寒生态脆弱区,植被极易受环境因素、人为放牧及天然灾害等影响,且遭受破坏之后在一定时间尺度上很难通过生态系统自我调节完成生态恢复。学者们对此问题得出的结果较一致,如周华坤等[18]得出随着高寒草甸退化程度的加深,植被盖度和优良牧草地上生物量比例逐渐下降。地上总生物量在轻度退化阶段最高,在极度退化阶段最低,随着退化加剧,杂草生物量显著增加,而莎草和禾草生物量显著减少。公延明等[19]得出随着退化程度加深及植物群落的逆行演替,优良牧草盖度、高度、频度等呈递减趋势,杂类草表现为增加趋势,植物群落物种多样性丧失。上述学者提到草地植被中的“杂草类”增加是相对于“优良牧草”而言的,本研究中不同退化草地优势种的改变与此有一定关联。
3.2 草地退化对土壤微生物特性的影响
本研究中土壤微生物量随草地退化程度的加重而降低是草地退化在土壤退化层面的表现,土壤微生物是土壤中的活性成分,其活性强弱、数量多少与土壤肥力的高低紧密联系着。微生物量碳、氮又是衡量土壤微生物活性的重要指标,其数值的降低直接说明了土壤肥力的下降。由草地退化引起土壤微生物活性降低,在草地生态系统中两者互相作用、相互影响。草地植被接受土壤微生物的溶磷、固氮、转化土壤有机质等作用的帮助而更好地获得环境中的营养物质,土壤微生物通过吸收植物根际释放的各类化学成分获取生长所需。两者关系密不可分,草地退化对土壤微生物生物量的影响很大,植被退化必定引起土壤微生物量的减少。大多学者也得出一致的结果,如郝金娥[20]研究得出玛沁和达日不同退化天然草地土壤微生物生物量碳随着退化程度的加重逐渐减少。周翰舒等[21]研究认为,未退化草地土壤微生物碳、氮含量高于退化草地。Wu等[22]研究认为内蒙古呼伦贝尔半干旱草原围栏内轻度退化草地的土壤微生物量碳、氮比放牧地高,很可能是因为植物凋落物及根际分泌物增加了土壤有机物的量;土壤有机质和土壤微生物量之间显著的相关性也说明微生物量是土壤C、N动态的敏感指示者。Li等[23]研究土壤利用管理方式对退化草地恢复的报告中指出虽然人工干预建植可以显著增加土壤微生物量,但是在草地植被丰富度、均匀度和多样性方面重度退化草地仍然比人工草地高。Wen等[24]研究高寒草地对碳库影响得出随着土壤有机碳的减少,土壤微生物量碳在土壤碳库中所占比例上升,同时随着退化程度的加深微生物量碳含量降低。但是牛得草等[25]研究围封与放牧对土壤微生物的影响认为,退化程度低的草地与退化程度高的草地内土壤微生物量碳含量无显著差异(P>0.05),而退化程度低的草地内土壤微生物量氮含量显著高于退化程度高的草地(P<0.05),这一结果与本研究退化程度加深,微生物量碳氮减少且差异显著的结果并不一致,原因可能是不同研究中选用的试验点土壤母质、气候条件及水分条件不同,因此微生物碳氮的变化规律也不同。同样王春燕等[26]研究恢复和改良内蒙古草地时得出,长期封育使土壤微生物碳含量降低。但是牛得草等[25]还得出不同土层的土壤微生物量碳、氮含量变化规律均为表层(0~10 cm)土壤高于下层(10~20 cm)土壤,本研究与其相同。
3.3 草地退化对微生物熵的影响
微生物熵的变化在本研究中与草地退化程度的变化两者之间并无规律性的联系。微生物熵作为土壤微生物量碳与土壤呼吸所产生CO2-C量的比值,其含义体现在了两方面上。土壤呼吸强度是土壤微生物的一个重要活性指标,同样也用于评价土壤肥力。能够完成土壤呼吸作用的不仅有微生物的呼吸作用,还有一小部分土壤动物和植物根系的呼吸作用。然而土壤呼吸作用的主要来源仍然是土壤微生物的贡献。魏卫东与刘育红[27]得出随着高寒草原退化程度的加剧,土壤CO2释放速率呈下降趋势,不同退化程度间土壤CO2释放速率差异达极显著水平,不同退化程度下土壤CO2释放速率基本呈未退化(UD)>轻度退化(LD)>中度退化(MD)>高度退化(HD)>极度退化(ED)的变化趋势。熊莉等[28]得出踩踏对草地土壤呼吸产生显著影响。本研究与上述结果一致,但本研究中微生物熵还包含微生物量碳值的改变,虽然呼吸速率在下降、其所产生的碳量在下降、同时微生物量碳也在下降,微生物熵值的大小实际上反映了两者下降程度的快慢比。朱炜歆等[29]认为同一草地类型的土壤呼吸强度、微生物碳都在各自土壤深度下随深度增加而减小,代谢熵的变化趋势刚好相反。这一结果与本研究中微生物熵在不同土壤层次上的变化规律相同,随着土层深度的增加而变大。
3.4 草地退化对草地生态系统“草-土-微生物”相互作用的影响
草地退化是目前我国广大牧区所面临的一个严重问题,不仅对牧区生态环境有重大影响,还会对牧区的经济发展造成干扰,因此,草地退化及其影响问题成了学术界研究的重点问题之一。综观前人研究,可以发现,前人大多是从植物层面或土壤理化性质方面展开的。但是,本课题组其他的研究发现[30-32]:草地退化与土壤微生物之间存在紧密联系。土壤微生物在草地生态系统中扮演了非常重要的角色,它不但能促进物质循环,还能够改变土壤理化性质,影响土壤肥力。因此,在研究草地退化机理及影响时,就必须考虑微生物因素,建立“草-土-微生物”三位一体的分析模型,本研究即是对植被特征和微生物特征的综合研究,研究发现草地退化在植被和微生物两个层次上都有体现,而这两个层次之间的因果关系如何,本研究尚不能得到明确答案,胡雷等[33],朱炜歆等[29],赵哈林等[34],尚占环等[35],McSherry和Ritchie[36]也探讨了这一问题,认为草地层面的植物多样性、物种组成、地上植被盖度、草地生产力、优良牧草比例,土壤层面的土壤呼吸强度、土壤有机质、全氮、全磷含量、土壤酶活力、土壤动物与微生物都存在一定相关性。但由于缺乏更充分的研究证据,这一问题还需进一步研究讨论。
4 结论
在草地退化程度不断加深的状态下,草地生态系统中的生产者、非生物环境与物质循环3个界面都发生着相应的变化,本研究中得出植被物种改变、结构与产量大幅度下降;土壤呼吸作用与地上产量及地下微生物量的改变具有相关性;土壤微生物活性降低,其中活性成分微生物量碳与微生物量氮显著减少。反之,草地生态系统在进行内部结构、能量分配等方面的调整,适应草地退化程度的加深。
[1] Su D X, Zhang Z H, Chen Z Z,etal. GB 19377-2003 Parameters for Degradation, Sandification and Salification of Rangelands[S]. Beijing: Chinese Standards Press, 2003.
[2] Xie G D, Lu C X, Leng Y F,etal. Ecological assets valuation of the Tibetan Plateau. Journal of Natural Resources, 2003, 18(2): 189-195.
[3] Xie G D, Lu C X, Xiao Y,etal. The economic evaluation of grassland ecosystem services in Qinghai-Tibet Plateau. Journal of Mountain Science, 2003, 21(1): 50-55.
[4] Piao S L, Fang J Y. Terrestrial net primary production and its spatio-temporal patterns in Qinghai-Xizang Plateau, China during 1982-1999. Journal of Nature Resource, 2002, 17(3): 373-380.
[5] Yan Y C, Tang H P. Differentiation of related concepts of grassland degradation. Acta Prataculturae Sinica, 2008, 17(1): 93-99.
[6] Chen Q G. Grassland deterioration in the source region of the Yangtze-Yellow rivers and integrated control of the ecological environment. Acta Prataculturae Sinica, 2007, 16(1): 10-15.
[7] Gao Q Z, Li Y E, Lin E D,etal. Temporal and spatial distribution of grassland degradation in Northern Tibet. Acta Geographica Sinica, 2005, 60(6): 965-973.
[8] Liu J Y, Xu X L, Shao Q Q. The spatial and temporal characteristics of grassland degradation in the Three-River Headwaters Region in Qinghai Province. Acta Geographica Sinica, 2008, 63(4): 364-376.
[9] Zhang J E, Liu W G. Utilization of microbes resources and sustainable development of agriculture. Soil and Environmental Science, 2001, 10(2): 154-157.
[10] Chen X R, Nan Z B. Bacterial diversity and its role in agricultural ecosystems. Pratacultural Science, 2002, 39(9): 34-38.
[11] Li X, Wang Y C. The biodiversity of soil microbes and plants. Acta Scientiarum Naturalium Universitatis Neimongol, 2006, 37(6): 708-713.
[12] Wang X P, Li X R, Xiao H L,etal. Evolution characteristics of the artificially re-vegetated shrub ecosystem of arid and semi-arid sand dune area. Acta Ecologica Sinica, 2005, 25(8): 1974-1980.
[13] Wang X L, Xu L G, Yao X,etal. Analysis on the soil microbial biomass in typical hygrophilous vegetation of Poyang Lake. Acta Ecologica Sinica, 2010, 30(18): 5033-5042.
[14] Zong N, Shi P L, Jiang J,etal. Interactive effects of short-term nitrogen enrichment and simulated grazing on ecosystem respiration in an alpine meadow on the Tibetan Plateau. Acta Ecologica Sinica, 2013, 33(19): 6191-6201.
[15] Xu G H, Zheng H Y. Manual of Soil Microorganism Analysis[M]. Beijing: Agricultural Press, 1986.
[16] Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C. Soil Biology Biochemistry, 1987, 19(6): 703-707.
[17] Brookes P C, Andrea L, Pruden G,etal. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biology Biochemistry, 1985, 12(6): 837-842.
[18] Zhou H K, Zhao X Q, Zhou L,etal. A study on correlations between vegetation degradation and soil degradation in the 'Alpine Meadow' of the Qinghai-Tibetan Plateau. Acta Prataculturae Sinica, 2005, 14(3): 31-40.
[19] Gong Y M, Hu Y K, Maidi A,etal. Alpine grassland community characteristics at the different stages of degenerating succession in Bayanbulak. Journal of Arid Land Resource and Environment, 2010, 24(6): 149-152.
[20] Hao J E. Study on the Number of Soil Microbes and Microbial Biomass Carbon Under Different Successional Stages of Vegetation Community in Yangtze and Yellow Rivers Source Region[D]. Xining: Qinghai University, 2010: 8.
[21] Zhou H S, Yang G W, Liu N,etal. Plant community and soil microbial characteristics in typical grasslands of different degradation degrees. Pratacultural Science, 2014, 31(1): 30-38.
[22] Wu X, Li Z H, Fu B J,etal. Restoration of ecosystem carbon and nitrogen storage and microbial biomass after grazing exclusion in semi-arid grasslands of Inner Mongolia. Ecological Engineering, 2014, 73: 395-403.
[23] Li Y Y, Dong S K, Wen L,etal. Soil carbon and nitrogen pools and their relationship to plant and soil dynamics of degraded and artificially restored grasslands of the Qinghai-Tibetan Plateau. Geoderma, 2014, 213: 178-184.
[24] Wen L, Dong S K, Li Y Y,etal. The impact of land degradation on the C pools in alpine grasslands of the Qinghai-Tibet Plateau. Plant and Soil, 2013, 368(1-2): 329-340.
[25] Niu D C, Jiang S G, Qin Y,etal. Effects of grazing and fencing on soil microorganisms and enzymes activities. Pratacultural Science, 2013, 30(4): 528-534.
[26] Wang C Y, Zhang J J, Lu Y L,etal. Effects of long-term grazing exclusion on soil organic carbon fractions in the grasslands of Inner Mongolia. Acta Prataculturae Sinica, 2014, 23(5): 31-39.
[27] Wei W D, Liu Y H. The influence of different degraded degradation of alpine steppe in source area of Lantsang, Yellow and Yangtze River on soil respiration. Hubei Agricultural Sciences, 2014, (8): 1102-1106.
[28] Xiong L, Xu Z F, Wu F Z,etal. Effects of stepping on soil respiration ofZoysiamatrellalawn during the winter dormancy period. Acta Prataculturae Sinica, 2014, 23(2): 83-89.
[29] Zhu W X, Suo N J, Gu Z K,etal. Soil properties associated with microbe in different types of Gannan grassland. Pratacultural Science, 2012, 29(10): 1491-1496.
[30] Ma W W, Yao T, Jin P,etal. Characteristics of microorganisms and enzyme activity under two plant communities in Desert Steppe. Journal of Desert Research, 2014, 34(1): 176-183.
[31] Zhang Y X, Yao T, Wang G J,etal. Characteristics of vegetation and soil inorganic nitrogen concentrations under different disturbed habitats in a weak alpine ecosystem. Acta Prataculturae Sinica, 2014, 23(4): 245-252.
[32] Lu H, Li X G, Yao T,etal. Characteristics of vegetation and soil microorganisms of molehill grassland in an ecologically vulnerable alpine region. Acta Prataculturae Sinica, 2014, 23(5): 214-222.
[33] Hu L, Wang C T, Wang G X,etal. Changes in the activities of soil enzymes and microbial community structure at different degradation successional stages of alpine meadows in the headwater region of Three Rivers, China. Acta Prataculturae Sinica, 2014, 23(3): 8-19.
[34] Zhao H L, Zhao X Y, Zhang T H,etal. Study on bio-processes in desertification in Northern Agro-Pasture Interzone. Journal of Desert Research, 2002, 22(4): 309-315.
[35] Shang Z H, Ding L L, Long R J,etal. Relationship between soil microorganisms, above-ground vegetation, and soil environment of degraded alpine meadows in the headwater areas of the Yangtze and Yellow Rivers, Qinghai-Tibetan Plateau. Acta Prataculturae Sinica, 2007, 16(1): 34-40.
[36] McSherry M E, Ritchie M E. Effects of grazing on grassland soil carbon: a global review. Global Change Biology, 2013, 19, (5): 1347-1357.
参考文献:
[1] 苏大学, 张自和, 陈佐忠, 等. GB 19377-2003天然草地退化、沙化、盐渍化的分级指标[S]. 北京: 中国标准出版社, 2003.
[2] 谢高地, 鲁春霞, 冷允法, 等. 青藏高原生态资产的价值评估. 自然资源学报, 2003, 18(2): 189-195.
[3] 谢高地, 鲁春霞, 肖玉, 等. 青藏高原高寒草地生态系统服务价值评估. 山地学报, 2003, 21(1): 50-55.
[4] 朴世龙, 方精云. 1982~1999年青藏高原植被净第一性生产力及其时空变化. 自然资源学报, 2002, 17(3): 373-380.
[5] 闫玉春, 唐海萍. 草地退化相关概念辨析. 草业学报, 2008, 17(1): 93-99.
[6] 陈全功. 江河源区草地退化与生态环境的综合治理. 草业学报, 2007, 16(1): 10-15.
[7] 高清竹, 李玉娥, 林而达, 等. 藏北地区草地退化的时空分布特征. 地理学报, 2005, 60(6): 965-973.
[8] 刘纪远, 徐新良, 邵全琴. 近30年来青海三江源地区草地退化的时空特征. 地理学报, 2008, 63(4): 364-376.
[9] 章家恩, 刘文高. 微生物资源的开发利用与农业可持续发展. 土壤与环境, 2001, 10(2): 154-157.
[10] 陈秀蓉, 南志标. 细菌多样性及其在农业生态系统中的作用. 草业科学, 2002, 39(9): 34-38.
[11] 李骁, 王迎春. 土壤微生物多样性与植物多样性. 内蒙古大学学报(自然科学版), 2006, 37(6): 708-713.
[12] 王新平, 李新荣, 肖洪浪, 等. 干旱半干旱地区人工固沙灌木林生态系统演变特征. 生态学报, 2005, 25(8): 1974-1980.
[13] 王晓龙, 徐立刚, 姚鑫, 等. 鄱阳湖典型湿地植物群落土壤微生物量特征. 生态学报, 2010, 30(18): 5033-5042.
[14] 宗宁, 石培礼, 蒋婧, 等. 短期氮素添加和模拟放牧对青藏高原高寒草甸生态系统呼吸的影响. 生态学报, 2013, 33(19): 6191-6201.
[15] 许光辉, 郑洪元. 土壤微生物分析方法手册[M]. 北京:农业出版社, 1986.
[18] 周华坤, 赵新全, 周立, 等. 青藏高原高寒草甸的植被退化与土壤退化特征研究. 草业学报, 2005, 14(3): 31-40.
[19] 公延明, 胡玉昆, 阿德力·麦地, 等. 巴音布鲁克高寒草地退化演替阶段植物群落特性研究. 干旱区资源与环境, 2010, 24(6): 149-152.
[20] 郝金娥. 江河源区不同植被演替阶段土壤微生物数量及微生物生物量碳的特性[D]. 西宁: 青海大学, 2010. 8.
[21] 周翰舒, 杨高文, 刘楠, 等. 不同退化程度的草地植被和土壤特征. 草业科学, 2014, 31(1): 30-38.
[25] 牛得草, 江世高, 秦燕, 等. 围封与放牧对土壤微生物和酶活性的影响. 草业科学, 2013, 30(4): 528-534.
[26] 王春燕, 张晋京, 吕瑜良, 等. 长期封育对内蒙古羊草草地土壤有机碳组分的影响. 草业学报, 2014, 23(5): 31-39.
[27] 魏卫东, 刘育红. 三江源区高寒草原不同退化程度对土壤呼吸的影响. 湖北农业科学, 2014, (8): 1102-1106.
[28] 熊莉, 徐振锋, 吴福忠, 等. 踩踏对亚热带沟叶结缕草草坪冬季休眠期土壤呼吸的影响. 草业学报, 2014, 23(2): 83-89.
[29] 朱炜歆, 索南吉, 顾振宽, 等. 甘南草地土壤微生物与理化特性. 草业科学, 2012, 29(10): 1491-1496.
[30] 马文文, 姚拓, 靳鹏, 等. 荒漠草原2种植物群落土壤微生物及土壤酶特征. 中国沙漠, 2014, 34(1): 176-183.
[31] 张玉霞, 姚拓, 王国基, 等. 高寒生态脆弱区不同扰动生境草地植被及土壤无机氮变化特征. 草业学报, 2014, 23(4): 245-252.
[32] 卢虎, 李显刚, 姚拓, 等. 高寒生态脆弱区“黑土滩”草地植被与土壤微生物数量特征研究. 草业学报, 2014, 23(5): 214-222.
[33] 胡雷, 王长庭, 王根绪, 等. 三江源区不同退化演替阶段高寒草甸土壤酶活性和微生物群落结构的变化. 草业学报, 2014, 23(3): 8-19.
[34] 赵哈林, 赵学勇, 张铜会, 等. 北方农牧交错区沙漠化的生物过程研究. 中国沙漠, 2002, 22(4): 309-315.
[35] 尚占环, 丁玲玲, 龙瑞军, 等. 江河源区退化高寒草地土壤微生物与地上植被及土壤环境的关系. 草业学报, 2007, 16(1): 34-40.
Vegetation and soil microorganism characteristics of degraded grasslands
LU Hu, YAO Tuo*, LI Jian-Hong, MA Wen-Bin, CHAI Xiao-Hong
CollegeofPrataculture,GansuAgriculturalUniversity,Sino-U.S.CentersforGrazinglandEcosystemSustainability,Lanzhou730070,China
Currently,studies of grassland degradation studies are mainly focused on soil properties, vegetation or soil microbiology separately; studies which consider all of these components are limited. This paper attempted to apply the perspective of systemic theory to provide scientific evidence for grassland degradation. Various degraded grasslands, located at the northeast margin of Qinghai-Tibetan Plateau, were selected for the study. Grassland characteristics were identified and associations between vegetation and soil microorganism properties were investigated using correlation analysis. The results showed that the number of plant species reduced with increased degradation and the dominant species changed.Additionally the height, coverage and aboveground biomass of the plant community decreased significantly (P<0.05) with enhanced degradation. Soil microbial carbon and nitrogen content also decreased with enhanced degradation. Aboveground biomass decreases by 88.6% from lightly degraded grassland (LDG) to severe degraded grassland (SDG). The microbial biomass of carbon (218.90 mg/kg) and nitrogen (44.32 mg/kg) in LDG were significantly higher than those in SDG (P<0.05), but there was no difference between moderate degraded grassland (MDG) and SDG. Soil microbial quotient was not affected by degradation. There was a significant positive correlation between aboveground biomass and microbial biomass (P<0.05).
vegetation characteristic; microbial biomass of carbon; microbial biomass of nitrogen; microbial quotient
10.11686/cyxb20150505
http://cyxb.lzu.edu.cn
2014-10-30;改回日期:2014-12-26
国家自然基金(31360584)和现代农业产业技术体系(CARS-35)资助。
卢虎(1986-),男,甘肃天水人,在读博士。E-mail: luhoo_7@aliyun.com *通讯作者Corresponding author. E-mail:yaotuo@gsau.edu.cn
卢虎,姚拓,李建宏,马文彬,柴晓虹. 高寒地区不同退化草地植被和土壤微生物特性及其相关性研究. 草业学报, 2015, 24(5): 34-43.
Lu H, Yao T, Li J H, Ma W B, Chai X H. Vegetation and soil microorganism characteristics of degraded grasslands. Acta Prataculturae Sinica, 2015, 24(5): 34-43.
