烤烟品种K326及其耐低温抗早花变异系基因表达谱分析
2015-04-15陈建军邓世媛邱妙文赵伟才
杨 静,陈 杰,陈建军,邓世媛,邱妙文,赵伟才,王 维*
(1.黔东南州烟草公司镇远县分公司,贵州镇远 557700;2.华南农业大学农学院,广州 510642;3.黔东南州烟草公司,贵州凯里 556000;4.广东省烟草南雄科学研究所,广东南雄 512400)
烤烟品种K326及其耐低温抗早花变异系基因表达谱分析
杨静1,2,陈杰3,陈建军2,邓世媛2,邱妙文4,赵伟才4,王维2*
(1.黔东南州烟草公司镇远县分公司,贵州镇远 557700;2.华南农业大学农学院,广州 510642;3.黔东南州烟草公司,贵州凯里 556000;4.广东省烟草南雄科学研究所,广东南雄 512400)
摘要:为了解烤烟早花与抗早花的形成及调控机制,本研究利用安捷伦烟草全基因组芯片分析了烤烟品种K326及其抗早花品系华烟06茎尖端花芽分化过程中基因的表达。结果表明,K326与华烟06距离各自花芽分化相同时间的茎尖端未花芽分化时的差异探针数最多,有5295个;K326分化当天各品种(品系)之间相比较差异探针数量最少,有2116个。与分子功能相关的差异基因主要与催化性能、转运性能、结合性能和转录调节性能有关;与细胞组成相关的差异基因主要是与细胞、细胞组分、细胞器相关的基因;与生物学过程相关的差异基因种类最多最复杂,涉及到植株抗逆性、生长、发育等方面,这对进一步研究与从中搜寻与烟草开花相关的基因奠定了基础。
关键词:烤烟;早花;抗早花;基因芯片;表达谱
烟草早花指植株未能达到正常生长应具有的高度和叶数就现蕾开花。国内外烤烟均存在烟草早花问题[1-2],严重影响着烟叶的生产。近百年来,人们在开花生理生化方面进行了大量的探索[3],直到1990年,Yanofsky等[4]成功地分离了花器官发育控制基因AGAMOUS,植物发育生物学的机理研究由以生理生化为主转变到以分子生物学及生理遗传学为主。拟南芥开花基因的研究开启了植物开花基因研究的大门,水稻、谷物、果树中与开花相关的基因的研究也已取得了很大的进展,与开花相关的

(http://www.shanghaibiotech.com),利用在线SAS系统进行数据分析。通过GO(Gene Ontology)分析对两倍以上的差异表达基因进行功能注释和富集分析。
1.5荧光定量PCR验证
利用烟草Actin基因(AB158612)为内参,从基因芯片分析结果中随机挑选出5个与开花相关的基因,进行实时荧光定量PCR,荧光定量PCR所用材料与基因芯片的材料及处理方式完全一样。3次重复,反应条件为95℃,10min;接着进行40个循环:95℃,15秒;60℃,30 s。算法为log2 2-ΔΔ Ct。
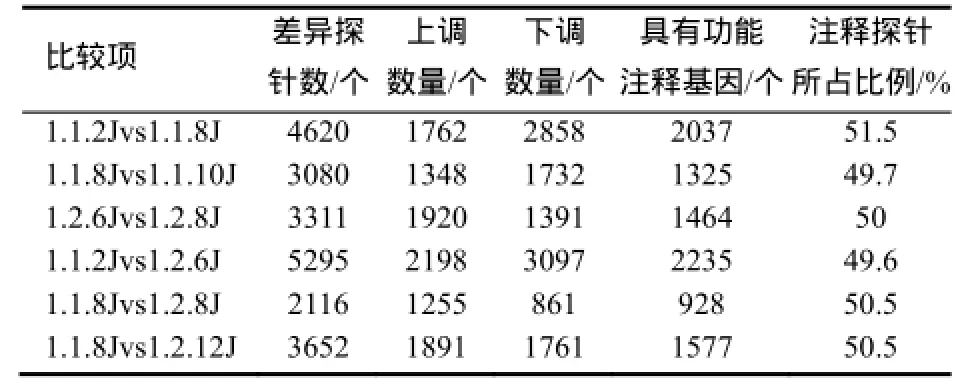
表1 差异探针分析Table1 Analysis of differential probes
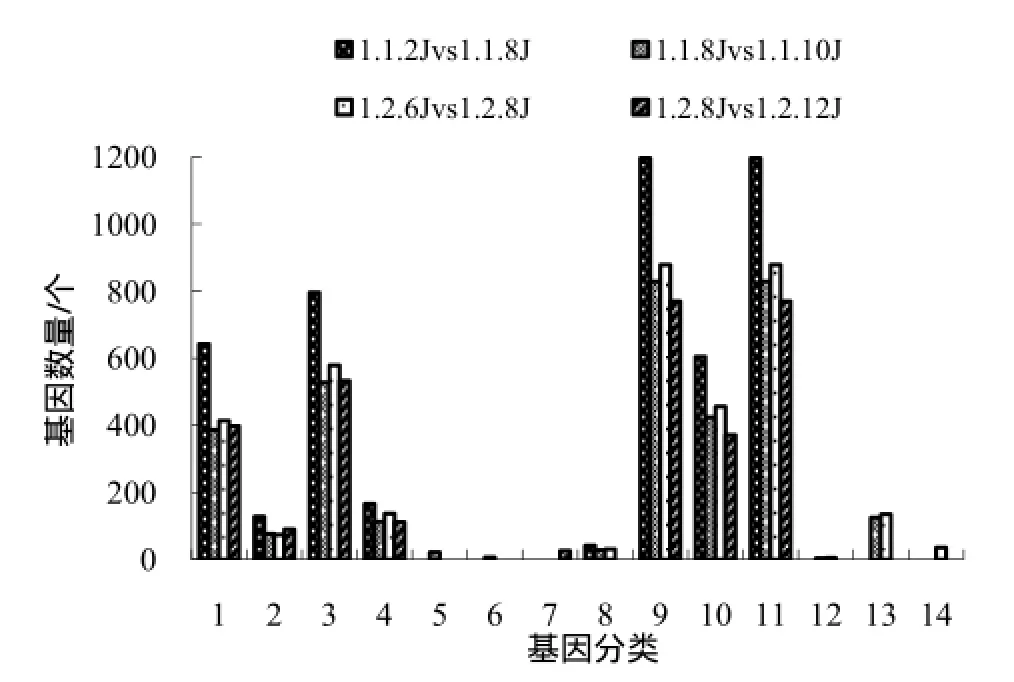
图2 与分子功能及细胞组成相关的显著富集基因Fig.2 Molecular function related and cellular component related significantly enriched genes
2 结果
2.1 芯片数据验证
试验中K326于4月11日现蕾,华烟06于4月24日现蕾,二者现蕾时间相差13 d。为了验证芯片分析结果,随机挑选了5个与开花有关并具有代表性的差异基因(CCR2,ELF4,AGL20,CBF1,COL9)进行荧光定量PCR验证,结果表示芯片表达谱信息具有较高的可靠性。
2.2芯片扫描结果中差异探针数的基本特征
按照两倍以上(FC>2或FC<0.5)差异为有效差异的标准筛选差异探针,将基因表达情况分别进行纵向比较和横向比较。纵向比较为K326和华烟06各自花芽分化过程中茎尖端基因表达情况的比较;横向比较包括:K326和华烟06距离各自花芽分化19 d时茎尖端基因表达情况的比较;K326花芽分化当天,K326与华烟06的比较;花芽分化时K326茎尖端基因表达情况与华烟06的比较(表1)。其中K326与华烟06距离各自花芽分化相同时间的茎尖端未花芽分化时的差异探针数最多,有5295个;K326分化当天各品种(品系)之间相比较差异探针数量最少,有2116个。
2.3纵向比较中差异基因的功能分类及富集分析
华烟06中与分子功能相关的差异表达基因可分为13类,而K326只有12类,其中少了蛋白标签(protein tag);华烟06中与细胞组分相关的差异基因有9类,K326有10类,多了与共质体(symplast)相关的基因(表2)。
K326花芽分化过程中,未分化与分化相比,分子功能中显著富集的差异基因主要与催化性能、转运性能、结合性能和转录调节性能有关,这与华烟06一致;K326花芽分化与分化后5 d相比,除了与催化性能、转运性能、结合性能和转录调节性能有关的基因在茎尖端显著富集之外,与酶调节活性、营养贮存活性有关的基因也显著富集;与细胞组成相关的显著富集的基因有胞外区、细胞、细胞器及细胞组分相关的基因等(图2)。
K326和华烟06花芽分化过程中与生物学过程相关的基因分类共有24种,通过极显著富集筛选出16种,其中包括发育过程、刺激应答、生殖过程等(表3)。
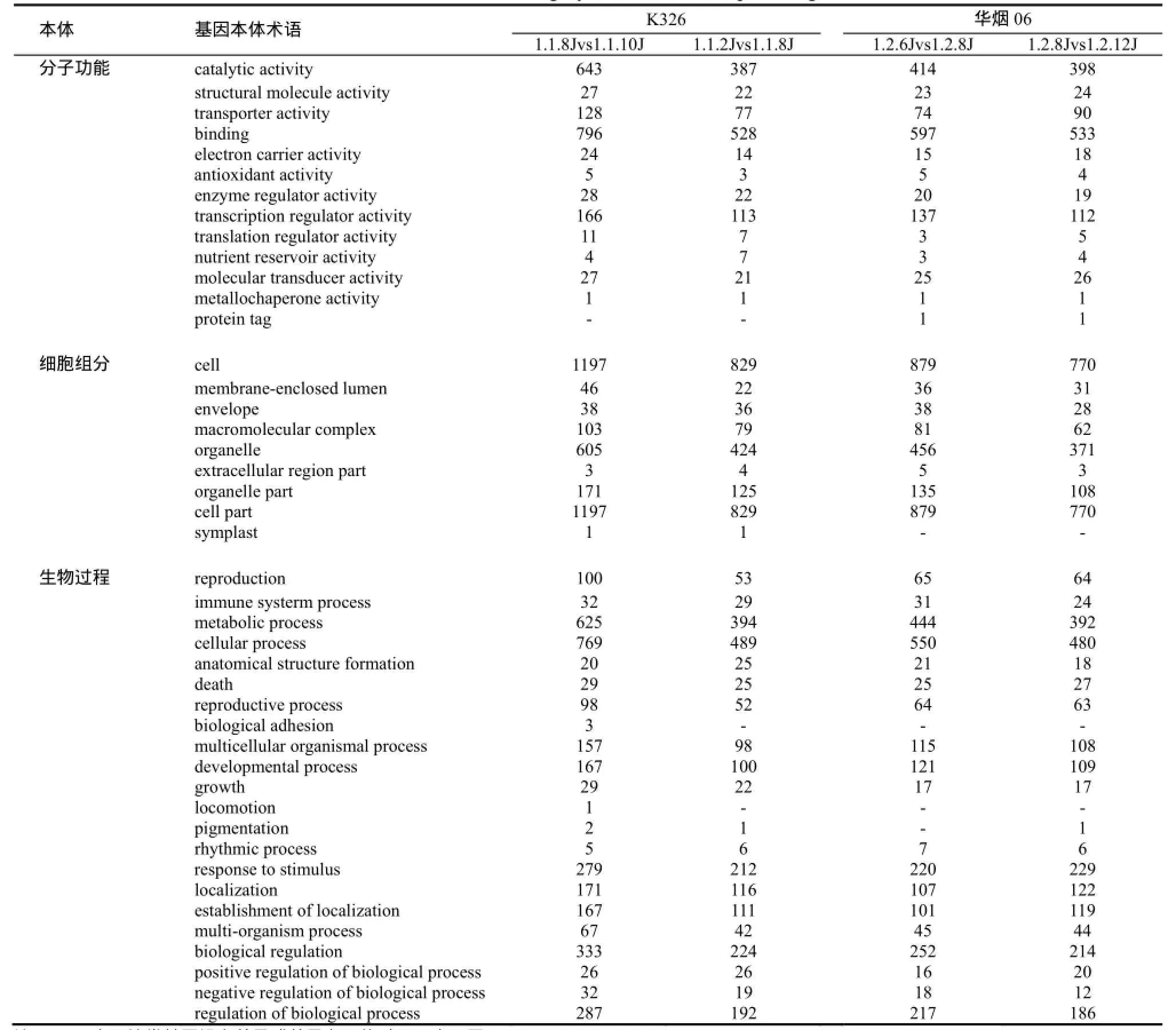
表2 差异表达基因的GO功能分类Table2 GO functional category of differential expressed genes
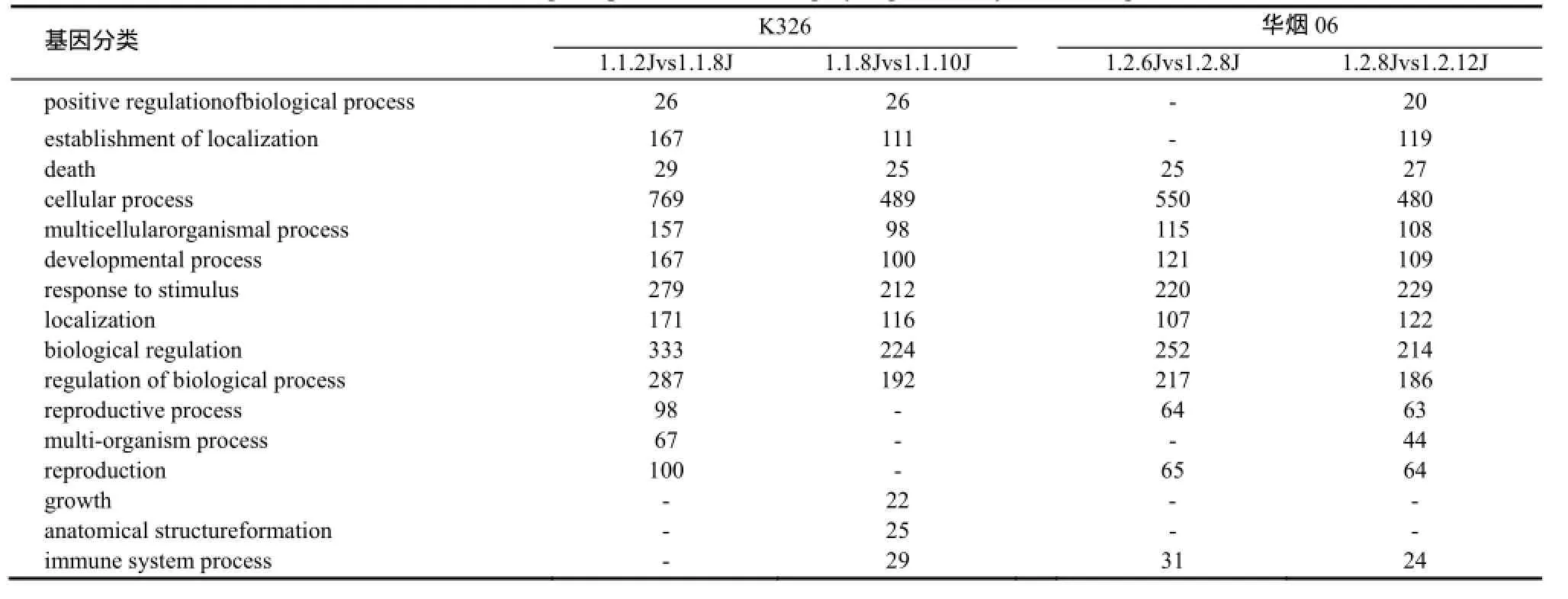
表3 与生物学过程相关的极显著富集基因Table3 Biological process related highly significantly enriched genes
2.4横向比较中差异基因的功能分类与富集分析
K326和华烟06距离各自花芽分化天数相同、花芽未分化的茎尖端基因表达情况相比,与分子功能相关的显著富集的基因主要与催化性能、转运性能、结合性能和转录调节性能相关,K326分化当天与华烟06相比,与催化性能、转运性能、结合性能和转录调节性能相关的差异基因显著富集,K326、华烟06花芽分化时的差异表达基因中,与催化性能、转运活性、结合性能、转录调节性能、分子转导活性有关的基因显著富集;K326和华烟06距离各自花芽分化相同天数的差异表达基因中,与细胞组成相关的显著富集的基因主要与细胞、细胞器、细胞组分、胞外区组分、胞外区相关,K326分化当天,与胞外区、细胞、细胞组分相关的基因显著富集(图3)。
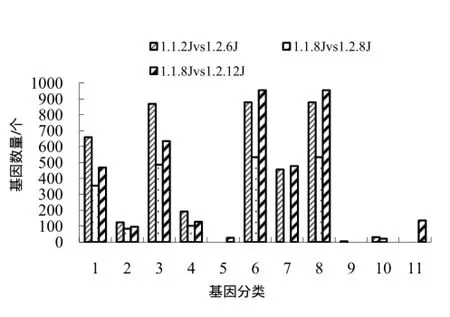
图3 横向比较中与分子功能及细胞组成相关的显著富集基因Fig.3 Molecular function related and cellular component related significantly enriched genes by horizontal comparison
K326分化当天差异表达基因最少,其中对死亡、刺激反应、定位、生物调节及生物调节进程相关、生物进程正向调节相关的差异基因都有极显著富集(图4)。
3 讨论
试验结果表明,在低温诱导下,K326移栽后65 d即开始现蕾,而华烟06则表现出了明显的抗早花特性,移栽后60 d才开始花芽分化。植物对环境刺激高度敏感,Lee等[12]的研究结果表明,即便是短时期的触碰和黑暗处理也会导致植物基因芯片检测结果存在较大差异。从本试验的比较结果看,K326与华烟06花芽均未分化且距离各自花芽分化时间相同点相比的差异探针数最多,达到5295个,而K326分化当天,二者之间相比差异探针数量最少,差异探针数为2116个,这说明基因表达差异不仅与植株的生长发育时期及遗传因素有关,更多的表达可能与气温及日照长短有关。
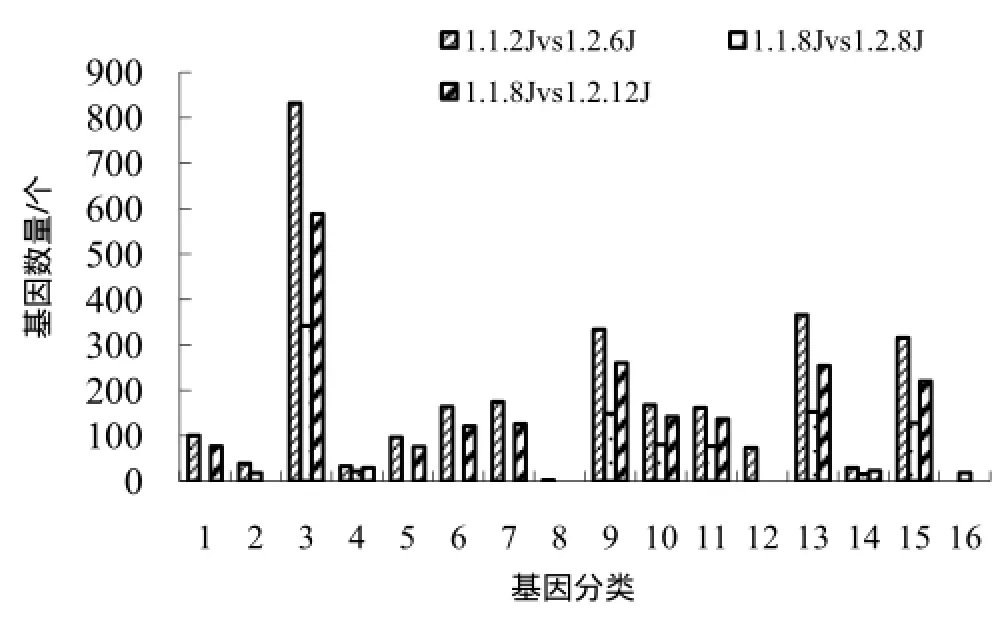
图4 横向比较中与生物学过程相关的极显著富集基因Fig.4 Biological process related highly significantly enriched genes by horizontal comparison
各品种(品系)生长发育过程中,分子功能、细胞组成、生物学过程相关的差异基因数量虽有一定差异,但分布比例是一致的,这说明K326与华烟06生长发育过程中基因总体的调控机制是相似的,这说明调控不同烤烟品种(品系)生长的基本机理相近,是否具有耐低温抗早花特性取决于少量特殊基因的表达。
为了进一步了解花芽分化过程中基因的表达变化及调控机制,有必要对差异基因进行显著和极显著富集分析,以确定差异表达基因行使的主要生物功能及其对植株花芽形成所具有的影响[13]。虽然K326花芽分化当天的差异基因数量最少,但通过显著富集分析,与营养储蓄相关的差异基因显著富集,这可能与植株的生长发育状态有密切关系。从与细胞组成相关的差异基因的显著富集分析看,K326花芽分化与分化后5 d相比的差异基因的分类要多于其未分化与分化时相比的差异基因分类;而华烟06花芽未分化1与花芽未分化2相比,与细胞组成相关的差异基因种类要比未分化2与分化时相比的多,调控细胞生长的基因的表达有待进一步研究。
4 结论
本试验通过比较分析,初步了解了K326及其抗早花品系花芽分化过程中基因的调控及表达机制。与分子功能相关的差异基因主要与催化性能、转运性能、结合性能和转录调节性能有关;与细胞组成相关显著富集的差异基因主要是与细胞、细胞组分、细胞器相关的基因;与生物学过程相关的差异基因种类最多最复杂,涉及到植株抗逆性、生长、发育等方面,这可为进一步研究并从中搜寻与烟草开花相关的基因提供参考依据。
参考文献
[1]SteinbergRA. Premature blossoming :effects of vernalization,seedling age and environment on subsequent growth and flowering of transplanted tobacco[J]. Plant Physiology ,1952,27(4):745-753.
[2] 张国,朱列书,王奎武,等. 烟草早花研究进展[J]. 作物研究,2005(5):383-385.
[3] 种康,雍伟东,谭克辉. 高等植物春化作用研究进展[J].植物学通报,1999,16(5):481-487.
[4] Yanofsky M F, Ma H, Bowman J L, et al. The protein encoded by the Arabidopsis homeotic gene agamous resembles transcription factors. Nature, 1990, 346:35-39.
[5] HorvathDP. Common mechanisms regulate flowering and dormancy[J]. Plant Science,2009,177(6) :523-531.
[6] Matsuda N, Ikeda K, Kurosaka M, et al. Early flowering phenotype in transgenic pears (Pyrus communis L.)expressing the CiFT gene[J].Japan.Soc.Hort.Sci,2009,78 (4): 410-416.
[7] 许志茹,李玉花. 基因芯片技术在植物研究中的应用[J].生物技术,2004,14(6):70-72.
[8] 朱素琴,季本华,陈名蔚. 高盐低温胁迫下水稻转录因子基因表达谱分析[J]. 科技通报,2010,26(6):844-852.
[9] 宋雯雯,李文滨,韩雪. 干旱胁迫下大豆幼苗根系基因的表达谱分析[J]. 中国农业科学,2010,43(22):4579-4586.
[10] 许州达,景瑞莲,甘强,等. 用水稻基因芯片筛选小麦耐旱相关基因[J]. 农业生物技术学报,2007,15(5):821-827.
[11] 曾黎琼,张伟,段玉云,等. 利用基因芯片分析非洲菊花序发育相关基因的表达[J]. 西南农业学报,2009,22(3):759-763.
[12] Lee D, Polisensky D H,Braam J.Genome-wide identification of touch and darkness-regulated arabidopsisgenes:a focus on calmodulin-like andXTHgenes[J]. New Phytologist,2005,165(2):429-444.
[13] 刘明,王米渠,丁维俊,等. 表达谱芯片数据的基因功能富集分析[J]. 生物医学工程学杂志,2010,27(5):1166-1168,1196.
中图分类号:S572.03
文章编号:1007-5119(2015)02-0093-06
DOI:10.13496/j.issn.1007-5119.2015.02.017
基金项目:国家烟草专卖局重点科技项目“耐低温抗早花烤烟新品种选育及遗传生理机理研究”(110200902043);广东省烟草专卖局(公司)科技项目“耐低温抗早花烤烟新品种选育”(200816);国家自然科学基金项目“抗早花烟草成花生理信号物质运转及其抵御低温的生理机理研究”(31101108)
作者简介:杨静,女,硕士研究生,研究方向为烟草遗传育种。E-mail:yangjinggzchina@163.com。*通信作者,E-mail:wangwei@scau.edu.cn
收稿日期:2014-07-08 修回日期:2015-03-09
Gene Expression Profile Analysis of Flue-cured Tobacco K326and thePremature Flowering Resistance Varieties
YANG Jing1,2, CHEN Jie3, CHEN Jianjun2, DENG Shiyuan2, QIU Miaowen4, ZHAO Weicai4, WANG Wei2*
(1.Zhenyuan County Tobacco Company of Guizhou Province, Zhenyuan, Guizhou 557700, China; 2.College of Agriculture, South China Agricultural University, Guangzhou 510642, China;3. Qiandongnan Tobacco Company of Guizhou Province, Kaili,Guizhou 556000, China;4.Nanxiong Institute of Tobacco Science, Guangdong Provincial Tobacco Company, Nanxiong, Guangdong 512400,China)
Abstract:To study the regulation and formation mechanismof premature flowering and premature flowering resistance of flue-cured tobacco, we used the Agilent Tobacco Oligo Microarray to deduce the expression profiling of flue-cured tobacco K326 and its premature flowering resistance variety Huayan06 under the flower bud differentiation process. The result showed that the same days from their flower bud differentiation the number of differential probes were the maximum amount of the comparative item between K326 and Huayan06. The number of the probe was 5295. On the day of K326 flower bud differentiation, the number of differential probe was the minimum amount. The number of the probe was 2116. The molecular function mainly included catalytic activity,transporter activity, binding and transcription regulator activity. The cellular component mainly included cell, cell part and organelle. The biological process was the most complex, involved plant resistance, growth, development, etc. It would establish a basis for the further study and search for genes associated with tobacco flowering.
Keywords:flue-cured tobacco; premature flowering; premature flowering resistance; gene chip; expression profile
