艾丁湖盐角草种子异型性及萌发特性
2015-04-11魏梦浩黄俊华
魏梦浩,黄俊华
(新疆农业大学林学与园艺学院,新疆 乌鲁木齐830052)
种子异型性(Seed Polymorphism)或种子二型性(Seed Dimorphism)是指在同一植株的不同部位产生不同形态或行为的种子的现象[1]。异型性种子之间往往在形状、大小、颜色和外部结构等形态学特征以及散布、休眠、萌发和幼苗生长等生态行为上存在明显差异[2]。这些异型性种子的萌发在时间上交替出现,降低死亡风险[3-7]。该现象多集中在菊科(Asteraceae)、藜科(Chenopodiaceae)和禾本科(Graminea)中,常出现在那些生长于荒漠和盐漠等环境条件高度多变生境中的植物上[1,8]。因此,种子异型性被认为是许多一年生植物对异质性环境的独特适应方式,在植物生活史对策和生态适应机制方面具有重要的研究价值[9]。藜科种子异型性的研究主要集中在滨藜属(Atriplex)[10]、盐角草属(Salicornia)[11]、猪毛菜属(Salsola)[12]和碱蓬属(Suaeda)[3]。
盐角草是藜科一年生盐生植物[13],株高10 ~40 cm,植株常常发红色。茎直立,多分枝;枝对生,肉质,通常长1 ~10 cm。叶鳞片状,顶端尖,基部连合成鞘状。穗状花序,花腋生,每1 个苞片内有3 朵花,集成一簇,陷入肉质地花序轴内;肉质花被,倒圆锥状,上部扁平成菱形;雄蕊伸出花被之外;花药矩圆形,子房卵形。已有研究发现,盐角草具有明显的种子异型性[9],且花序中央花产生的种子总是比两个边花产生的种子大,边花的种子比中央花种子更不耐盐且多休眠[14]。常分布于海拔400 ~700 m 的盐湖边、盐化沼泽边、潮湿盐土及重盐土等平原地区[15]。
前人做了大量藜科植物种子异型性的研究,但对于盐角草属的报道则很少,在国内更是寥寥无几[16]。本研究以新疆境内吐鲁番盆地艾丁湖的盐角草种群为研究对象,充分考虑新疆特殊的地理环境产生的影响,通过比较种子形态、扩散方式、萌发特征等,分析该特殊地理环境中盐角草种子异型性特征以及对该物种在逆境中生存发展的意义。
1 材料与方法
1.1 研究区概况
艾丁湖位于新疆维吾尔自治区吐鲁番市东南30 km,吐鲁番盆地最低洼处,湖面海拔-154 m。盐沼湖水为氯化钠型水,含盐量达200 g·L-1,盐壳地的盐壳厚0.2 ~1.5 m,含盐50% ~95%。冬季气温在0 ~15℃,春秋季节温度在19 ~38 ℃。艾丁湖为“季节性湖泊”。湖区年平均降水量别为6 ~25 mm,而年蒸发量却高达2 727 ~3 720 mm[17],地表由砂粘土和盐壳组成。湖泊周边地区植物枯死,生态明显恶化。夏季炎热,地势闭塞,形成了独特的生态环境[18]。
1.2 种子采集与保存
2013 年9 月底,于盐角草开花后期在新疆吐鲁番盆 地 艾 丁 湖 南 岸(42° 38' 26″ - 42° 38' 45″ N、89°15'52″-89°15'07″ E、海拔-154 m),根据盐角草种群空间分布差异确定的4 块样地进行盐角草的生态-生物学特性调查。
2014 年10 月中旬于盐角草种子成熟期在4 块样地的自然群落中采样,随机采集自然成熟的盐角草40株。取样时,对每株盐角草小心挖取(包括根、茎、果),并单株放置于密封袋内保存,注意保持植株各构件的完整性,采收后置于通风条件良好的室内风干,收集种子,在室温下保存。
1.3 种子的类型确定
随机选取盐角草20 株,观察并记录种子着生部位、翅的发育状况,记录不同类型种子的重量、形状、颜色和着生方式。
种子重量的测定以100 粒×4 组的平均值,用千分之一天平(上海菁海仪器有限公司,JA1003)称重。
1.4 异型种子形态及微形态特征观察
形态学观察:用Nikon SMZ25 型体式显微镜记录种子长度、直径、形状、外皮附属物,颜色主要参照“CMYK 印刷专用精选色谱”,大小、形状参照刘长江的《中国植物种子形态学研究方法和术语》[18]。种子长度和直径的测量以30 粒种子的平均值为准。
微形态观察:选取成熟饱满的种子,除去外部包裹物,每个样品选择3 ~4 粒,经过超声波75%酒精清洗,然后干燥,在解剖镜下粘贴于粘有双面胶的样品台上,JFC-1100 型真空喷镀机黄金离子溅射,JSM-6360LU 型扫描电镜观察和拍摄照片。
1.5 异型种子的吸水特性
种子吸水试验前用1%的高锰酸钾(KMnO4)浸泡种子5 min 进行表面灭菌处理,后经蒸馏水冲洗,在室内环境(气温20 ~25 ℃,相对湿度45% ~50%)中进行,将种子置于直径为90 mm 并且垫有两层滤纸的培养皿中(在整个过程中保持滤纸湿润),并开始计时。分别在吸水0.5、1、2、4、8、24、48 h 时将种子取出,迅速擦干、称重,后再放回培养皿中。每组20 粒3 个重复。吸水特性结果用种子吸水重量增加率(Wr)表示。计算公式为:
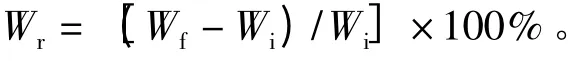
式中,Wr表示种子吸水重量增加率;Wf表示不同时间段种子吸水后的重量;Wi表示种子原重量。
1.6 不同变温条件异型种子的萌发特性
根据吐鲁番艾丁湖1959 -2007 年气候资料统计春季(3-5 月)日平均最高气温和日平均最低气温,共设定光暗交替的(12 h 光照/12 h 黑暗下)15/2 ℃、20/10℃、25/15 ℃共3 个变温条件进行萌发培养。
试验前各选取颗粒饱满的成熟种子用1%的次氯酸钠(NaClO)消毒20 min 进行表面灭菌处理,蒸馏水充分冲洗,将种子置于直径为90 mm 培养皿中培养,以两层滤纸为基质,加入10 mL 蒸馏水(在整个萌发过程中保持滤纸湿润),每组25 粒、4 个重复,放入RTOP-260Y 光照培养箱内萌发。种子的萌发以胚根的出现为标志,萌发过程中每24 h 统计1 次种子萌发数。以连续3 d 不萌发为结束,萌发结果以萌发率表示。计算公式为:
萌发率=(萌发种子数/参试种子总数)×100%。
2 结果分析
2.1 盐角草的多型性种子类型
根据种子的颜色、着生方式、大小、重量、形状和有无翅等特性,将盐角草的种子分为A 型、B 型和C 型3种类型(表1)。可以看出,百粒重大的种子体积也较大。
2.2 异性种子形态结构
2.2.1 种子形态特征 A 型种子为棕黄色,呈椭圆形(图1),主要分布于穗状花序的中央花位上,宿存花被的背部有红褐色翅状附属物,散布单位的直径为(2.496 ±0. 265)mm,单位质量最大,百粒重为0.109 g(表1),容易从母体上脱落,且翅较长易被风传播到较远的地方。
B 型种子为姜黄色,呈椭圆形(图1),主要分布于中上部穗状花序的旁边花位上,宿存花被的背部有红褐色翅状附属物,较为短小,散布单位的直径为(1.709 ±0.181)mm(表1);较容易从母体脱落,翅短随风传递到较近的地方或散落于植株附近。
C 型种子为棕色,呈椭圆形(图1),随机分布于植株中下部穗状花序的旁边花位上,种皮外部没有附属物,种子直径仅为(1.147 ±0.085)mm(表1),单位质量最小,百粒重仅为0.02 g;不易从母体脱落,脱落后散落于母本植株附近。
2.2.2 种皮表面结构 扫描电镜观察结果表明:A、B型种子表面粗糙,有棱状纹饰,C 型种子表面较粗糙,有稀疏棱状纹饰(图2);A、B 型种子种皮无附属物,一侧生长有钩刺,长度分别约为60、80 μm(图2);C 型种子种皮表面均分布有刺,长度约为20 μm,但主要集中分布于种脊处,部分刺前段卷曲形成钩刺(图2k),并且发现表面钩刺均为中空结构(图2l)。
2.3 异型种子吸水特性
异型种子的吸水特征存在着明显的差异(图3)。A 型种子吸水0. 5 h 后重量增加了(38. 9% ±1.8%),随后种子重量增加速率快速增高,4 h 后增加(89.39% ±3.7%),种子开始萌发。B 型种子吸水0.5 h 后重量增加了(26.7% ±1.9%),随后种子重量增加速率快速增高,8 h 后增加了(92.0% ±4.1%),随后种子开始萌发。C 型种子吸水0.5 h 后重量仅增(2.5% ±0.3%),1 h 后重量增加(22.5% ±1.3%),随后种子重量增加速率放缓,24 h 后吸水接近停滞,48 h 后开始萌发。
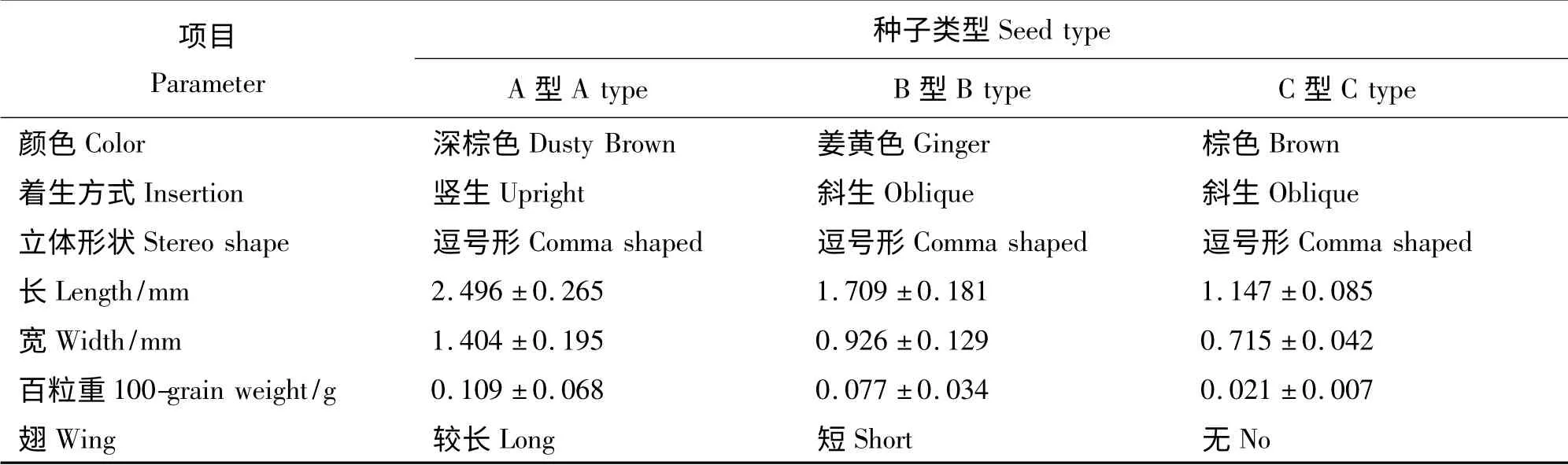
表1 盐角草3 种类型种子的特征Table 1 Characteristics of types A、B、C utricles of S. europaea

图1 盐角草多型种子类型Fig.1 The four dispersal units and fruit types of S. europaea
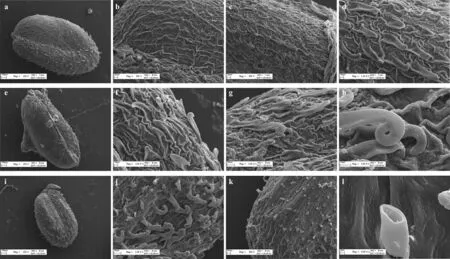
图2 盐角草多型种子表面纹饰Fig.2 The four dispersal units and exine ornamentation of S. europaea
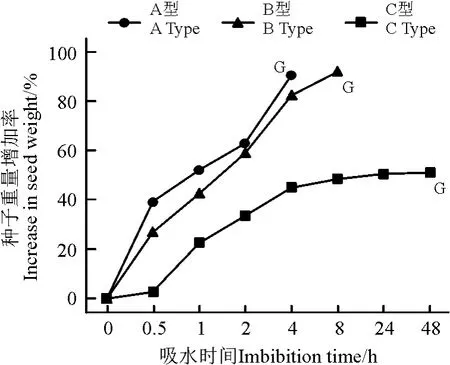
图3 盐角草异型种子吸水曲线Fig.3 Imbibitions curves for dimorphic seeds of S. europaea
变温萌发试验结果表明(图4),A 型种子在变温条件下萌发率在3 种类型种子种均是最高(>74.67%),这表明了A 型种子具有相对较宽的萌发温度范围;B 型种子在变温条件下萌发率略低于A 型种子(>53.33%);C 型种子在3 种变温条件下的萌发率都很低(<10%),表明其存在休眠现象。
3 讨论与结论
盐角草是一年生的盐生植物[9],广布于全世界各种生境,在新疆半干旱区和盐碱地也有分布,根据种子大小和着生位置划分种子类型已有报道[20]。早在20世纪60 年代初期Konig[9]和Dalby[14]的研究发现,一个花序中央花的种子总是比两个边花种子大。20 世纪70 年代后期Ungar[19],Jeyarany 和Ungar[21]根据萌发试验发现,盐角草有两种类型的种子。但是对于综合着生位置、种子大小、扩散方式的划分类型从未报道。或许是由于物种在长期的生存发展过程中为了更好地适应不良环境的冲击产生了新型种子,或是新疆本 地 特 殊 的 极 端 地 理 条 件、 环境条件所致,本研究通过综合着生位置、种子大小、扩散方式、萌发特性等特点发现,位于新疆吐鲁番艾丁湖盆地的盐角草产生中央花位、边花位和边花位小型3 种类型的种子,且他们在种皮颜色、大小、质量、种子着生方式、表面附属物、萌发、休眠特性、吸水能力等方面均存在显著差异。这对于进一步细化盐角草对抗严重异质化环境所作出的生活史对策,以及异型性种子是如何增强物种应对严酷生境的灵活性,提高了该物种对异质化环境的适应性的研究起到了承前启后的作用。
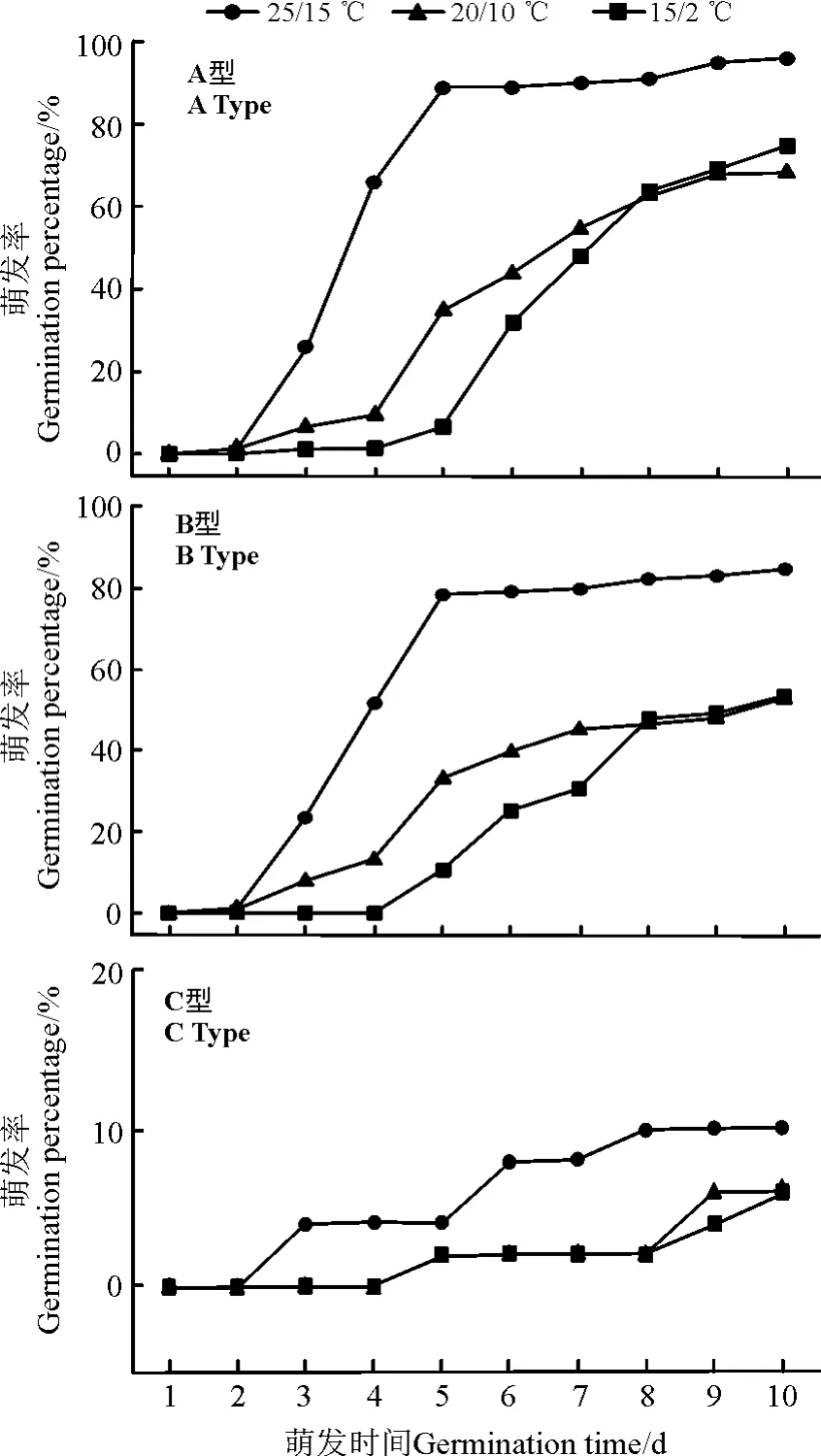
图4 盐角草3 种类型种子在不同变温条件下的发芽率Fig.4 Germination percentage of 3 seed types of S. europaea at different alternating temperatures
盐角草3 种类型的种子在大小和质量上有着显著差异。一般大粒种子有较大的胚,萌发和生长一般较快;小粒种子存在休眠,可形成种子库,确保物种延续。盐角草3 种类型的种子同时具备了大、小种子的优势,从而可以更加适应异质化严重的荒漠环境。
根据各种类型种子的特征可得出:A 型种子主要分布于中间花序部位,最为成熟,最易从母体上脱离,可随风传播到较远的距离,可作为“进攻者“扩大自己的生存范围,从而扩大种群面积;B 型种子分布在植株的边花序部位,成熟后较易从母体上脱离,脱落后散布于母体附近,可作为“防守者”来延续种群在原生境下的繁衍,或可随风传播至较远的距离,可作为A 型种子的“进攻辅助者”协助A 型种子扩大生存范围;C 型种子主要随机分布于植株的中下部的边花序,存在休眠现象,不易脱落,且花被片无翅状附属物,只能散落在母体底部或是附近,可在土壤中形成种子库,再经低温等刺激破除休眠后具备萌发的能力,作为植株抵御逆境的“后备力量”存在,确保物种的延续。这种不同类型种子拥有不同类型扩散方式及作用的现象在藜科植物紫翅猪毛菜(Salsola affinis)中也有报道[22]。
种子异型性通常与种子萌发多样性联系在一起[11],在盐角草中也存在这种现象。而温度是种子萌发和出苗的基本条件之一,它会影响种子萌发的一系列过程,不同类型的种子由于其生理特性与物理特性的不同,其温度对种子萌发的影响也不同[23],温度过高或者过低都会影响种子的活力造成萌发和出苗不良[24]。
总结前人研究得出的盐角草异型种子的描述,较小种子出现在花序的边花,由于种子有初期休眠现象,对光敏感,因此是主要的种子库成分,种子库中的种子发芽最高值出现在4 -5 月;较大的种子在花序的中间,对环境要素不敏感,种子库中的比例很小[25]。但是,对于综合环境因素对该物种种子萌发时所产生的影响,以及种子萌发时在物种繁衍过程中所承担的作用在已有报道中并未提及。
本研究中,盐角草A、B 类型种子在15 ℃/2 ℃环境下,都有萌发但受到不同程度的抑制;而25 ℃/15℃环境下时,A、B 型种子萌发速率加快且萌发率都达到最高;这表明该物种的A、B 型种子萌发范围较宽但在较高温度下萌发效果最好。这对于在异质化严重的环境中繁衍十分有利。在春季融雪温度回升后A、B型种子开始陆续萌发,在温度可能达到最适条件时迅速大批量萌发,以保证该物种一年中个体数量的充足为来年提供足够的种子用以繁衍,但在野外环境中早期萌发的种子很容易受到环境波动的影响造成大批量的死亡。
C 型种子在3 种变温条件下的萌发率都很低,并且萌发速度最慢,表明其存在休眠现象,可在土壤中形成种子库。该类型的种子可在环境恶劣时不进行萌发,等到条件适宜时再经低温等刺激破除休眠后具备萌发的能力,作为植株抵御逆境的“后备力量”存在,确保物种的延续。
Jeyarany 和Ungar[21]在1984 年发表的文章中写道,盐角草小种子的萌发需要光照。自然环境下60%的大种萌发,但只有4%的小种子萌发[23]。春季土壤水分和温度的升高有助于种子萌发。然而,春季(5 月)降水和温度的显著波动会影响早期种子的萌发或是早期萌发的幼苗死亡。在这种情况下,推迟萌发的小种子萌发后则可补充地上生物量。盐角草通过生产不同萌发特征的种子是分配资源的一部分,大种子采取K-策略抢占资源。小种子存在休眠,可作为在种子库确保种群在一个地方长期存储单位。
通过研究盐角草异型性种子萌发特性,发现盐角草异型种子属于“两头下注”策略,都具有不同的萌发格局。该物种异型种子的萌发具有明显的时空差异,A、B 型种子采取“机会主义”策略,他们在较宽的温度范围内都可以萌发,温度条件合适时,将迅速萌发;在荒漠或干旱条件下,适合萌发和生长的时节是短暂并且不可预测的,这种在较宽温度范围内可快速萌发的能力使盐角草在早春利用任何可用的降水、升温等有利环境条件迅速萌发[16]。
吐鲁番艾丁湖春秋季节温度在19 ~38 ℃,昼夜温差极大,湖区年平均降水量为6 ~25 mm,而年蒸发量却高达2 727 ~3 720 mm。盐角草在艾丁湖自然环境下,当年种子成熟散落后,会经过冬季低温,待来年回温后,才能萌发。但是,如果植物的种子由于某一次升温,有活力的种子同时萌发,之后的降温可能会导致全部个体的死亡[26]。干旱或半干旱地区的植物可能通过保持长时间连续萌发,外种皮附着翅或其他多种因素阻碍其萌发,从而降低风险。
C 型种子采取“保守主义”的策略,在各种温度下萌发均很低,具有休眠现象,经过外界刺激后才能萌发。C 型种子无翅,且质量轻、不易脱落,脱落后也分散在母体附近,大部分会进入种子库,占领最适生存生境,是作为避免种群灭亡的储存力量。种子库对于在时间和空间上的不可预测环境中的物种具有重要意义,可降低或避免盐角草种子完全的响应一次降雨而带来的奉献。
在异质化严重的环境下对植物的生长、生存极为不利,盐角草3 种类型的种子,使得该物种可以扩散到不同距离范围,避免了密度过高后的种内竞争。3 种类型种子的不同萌发特性,使得盐角草种子在面对强烈多变的环境时不至于完全灭亡。盐角草种子既具备“机会主义”又具备“保守主义”,种群的扩张即具备“进攻型”、“进攻辅助型”和“防御性”,这些对策的相互协调合作扩大了盐角草种子萌发的时间范围,增加对环境条件的适应性,有利于其在严酷的环境中成功定居和繁衍。
[1] Venable D L.The evolutionary ecology of seed heteromorphism[J].The American Naturalist,1985,12(6):577-595.
[2] Baskin C C,Baskin J M.Seeds:Ecology,Biogeography and Evolution of Dormancy and Germination[M].San Diego:Academic Press,1998.
[3] Khan M A,Gul B,Weber D J.Germination of dimorphic seeds of Suaeda moquinii under high salinity stress[J].Australian Journal of Botany,2001,49:185-192.
[4] Venable D L.Bet hedging in a guild of desert annuals[J]Ecology,2007,88:1086-1090.
[5] Wei Y,Dong M,Huang Z Y.Seed polymorphism,dormancy and germination of Salsola affinis(Chenopodiaceae),a dominant desert annual inhabiting the Junggar Basin of Xinjiang,China[J].Australian Journal of Botany,2007,55:464-470.
[6] Wang L,Huang Z Y,Baskin C C,Baskin J M,Dong M.Germination of dimorphic seeds of the desert annual halophyte Suaeda aralocaspica (Chenopodiaceae),a C4plant without Kranz anatomy[J].Annals of Botany,2008,102:757-769.
[7] Yao S X,Lan H Y,Zhang F C.Variation of seed heteromorphism in Chenopodium album and the effect of salinity stress on the descendants[J].Annals of Botany,2010,105:1015-1025.
[8] Ungar I A.Population ecology of halophyte seeds[J].Botanical Review,1987,53:301-334.
[9] Konig D.Beitrage zur Kenntnis der deutschen Salicornien[J].Mitt Florist-Soziol.Arbeitsgem,1960,8:5-58.
[10] 李伟强,刘小京,毛任钊,安萍,乔海龙,黄玮,李志刚.植物种子二形性(多形性)研究进展[J].生态学报,2006,26(4):1234-1242.
[11] Berger A.Seed dimorphism and germination behavior in Salicornia patula[J].Plant Ecology,1985,61(1-3):137-143.
[12] Ellison A M.Effect of seed dimorphism on the density-dependent dynamics of experimental populations of Atriplex triangularis(Chenopodiaceae)[J].American Journal of Botany,1987,74(8):1280-1288.
[13] Christy T G,Ungar I A.Germination response of dimorphic seeds of two halophyte species to environmentally controlled and natural conditions[J].Canadian Journal of Botany,2003,81(9):918-926.
[14] Dalby D H.Chromosome number,morphology and breeding behavior in the British Salicorniae[J].Watsonia,1962(5):150-162.
[15] 李疆,尹景原,卡得尔,李新.吐鲁番盆地的降水分布特征[J].新疆地理,1984,7(2):22-30.
[16] 王宗灵,徐雨清,王刚.沙区有限降水制约下一年生植物种子萌发与生存对策研究[J].兰州大学学报:自然科学版,1998,34(2):98-103.
[17] 谢宏琴,贾国东,彭平安,穆桂金.艾丁湖二千余年来环境演变的地球化学记录[J].干旱区地理,2005,28(2):205-209.
[18] 刘长江,林祁,贺建秀.中国植物种子形态学研究方法和术语[J].西北植物学报.2004,24(1):178-188.
[19] Ungar I A.Halophyte seed germination[J].Botanical Review,1978,44:233-264.
[20] Bewley D J,Black M.Seeds:Physiology of Development and Germination[M].New York:Plenum Press,1994:1-10.
[21] Jeyarany P,Ungar I A.The effect of seed dimorphism on the germination and survival of Salicornia Europaea L. populations[J].American Journal of Botany,1984,71(4):542-549.
[22] 王宏飞,魏岩.紫翅猪毛菜的种子多型性及其结实格局[J].生物多样性,2007,15(4):419-424.
[23] Ajmal K M,Bilquees G,Darrell J W.Influence of salinity and temperature on the germination of Kochia scoparia[J].Wetlands Ecology and Management,2001,9:483-489.
[24] 常水晶,左兵,王晓炜,黄俊华.光照、温度及盐分对钠猪毛菜种子萌发的影响[J].干旱区地理,2008,31(6):897-903.
[25] Ungar I A.Seed dimorphism in Saliconia europaea L.[J].Botanical Gazette,1979,140:102-108.
[26] Wei Y,Dong M,Huang Z Y,Tan D Y.Factors influencing seed germination of Salsola affinis(Chenopodzaceae),a dominant annual halophyte inhabiting the deserts of Xinjiang,China[J].Flora,2008,20(3):134-140.
