二倍体棱叶蒜花粉活性的变化特征
2015-04-11席延坡林辰壹刘俊霞孙新领
席延坡,林辰壹,刘俊霞,孙新领,鲍 亮,薛 梅,吴 慧
(1.新疆农业大学林学与园艺学院,新疆 乌鲁木齐830052;2.新疆农业大学设施农业研究所,新疆 乌鲁木齐830052)
花粉作为父本重要的基因流载体,与花粉育性一起决定了雄性亲本的遗传信息是否能够传递给下一代[1]。花粉的寿命和生活力是植物长期适应生态环境变化、保障自身在自然条件下繁殖成功的进化结果,与花粉的竞争力、自交选择、花粉传播距离、花粉移出、花粉呈现以及柱头可授期等密切相关[2],因此,花粉的寿命和生活力一直是植物繁殖生物学重要的组成部分[3]。棱叶蒜(Allium caeruleum),又称棱叶薤[4]、兰花山蒜[5],在中国仅分布于新疆天山一带的低山带[4-6],国外分布在俄罗斯伏尔加河下游到西伯利亚,是重要的野生葱属植物资源,具较高的食用、饲用、观赏和药用价值[6]。新疆不同地理位置分布的棱叶蒜居群间存在着二倍体、三倍体和四倍体的倍性分化[7],然而不同倍性棱叶蒜繁殖特征尤其是花粉寿命以及活力特征尚未见报道。本研究以乌鲁木齐市雅玛里克山原生境下分布的二倍体棱叶蒜不同开花物候期的花粉为研究对象,比较并选择二倍体棱叶蒜花粉适宜的培养方法及生活力检测的方法,检测其花粉活力和花粉萌发率变化规律。该研究结果将有助于克服杂交育种中亲本花期不遇和远距离杂交的困难,提高植物育种效率,为该野生植物资源的迁地保护、引种驯化和杂交育种提供科学依据,同时,为其他倍性分化的野生棱叶蒜植物资源繁育系统研究和种质资源开发利用奠定基础。
1 材料与方法
1.1 材料
研究材料棱叶蒜(2n =2x)分布于乌鲁木齐市雅玛里 克 山 阴 坡 面,地 理 位 置 为44°07'26. 6″ N,87°34'11.8″ E,海拔983 m,为温带大陆性气候,年平均降水量为208.4 mm,年平均日照数为2 813.5 h,年均无霜期179 d[8]。
1.2 花粉采集
基于棱叶蒜开花物候特征,在原生境下分别于棱叶蒜的开花始期、开花盛期和开花末期标记完全伸出花被片还未散粉的、饱满的、健康的花药,基于前期研究成果根据开花动态,采集当日即将自然散粉的小花,置于无菌培养皿中带回实验室后观察,当花药开裂后立即测定花粉活力[9]。
1.3 花粉活力测定
分别采用联苯胺α -萘酚法、0.5% 氯化三苯基四氮唑(TTC)法、醋酸洋红法、碘-碘化钾法(I2-KI)、孢粉染色法、红墨水染色法和四甲基偶氮唑(MTT)法,共计7 种染色方法测定不同开花物候期的花粉活力[9-12]。将新鲜花粉置于载玻片上,分别滴入上述溶液,混合均匀后于25 ℃的恒温箱内静置,联苯胺α -萘酚处理15 min;0.5% TTC 处理20 min;醋酸洋红处理2 ~3 min;I2-KI 处理5 min;孢粉染色处理15 min;红墨水染色处理15 ~20 min 以及MTT 处理5 min。以经过高温杀死后的无活力花粉为对照。以花粉的着色率代表花粉活力,计算花粉活力百分率,花粉活力=(有活力的花粉数/花粉总数)×100%。
1.4 花粉萌发率测定
采用二因素完全随机的离体萌发法,进行花粉萌发率测定的配方选择与优化[13]。蔗糖浓度分别为0、20、50、100、150 g·L-1,硼酸浓度分别为0、50、80、100、150 mg·L-1,各处理均添加3 g·L-1的琼脂,用0.1 mol·L-1的NaOH 和1 mol·L-1的HCl 调节pH至5.8。取居群开花盛期的花粉进行优化配方检测;取居群不同开花物候期的花粉进行花粉萌发率变化特征测定。在25 ℃的恒温培养箱内培养4 h。以经过高温杀死的花粉为对照,以花粉管伸长长度超过或等于花粉粒直径的2 倍记为萌发花粉。计算花粉活力和萌发率,花粉萌发率=(萌发的花粉数目/花粉总数)×100%。
1.5 数据统计与分析
花粉活力和花粉萌发率试验每个处理重复3 次,在10 ×10 倍光学显微镜(Motic 2000)下每张制片随机选取5 个视野观察并统计着色花粉或萌发花粉,每视野不少于30 粒花粉。试验所得数据采用“平均值±标准差”表示,并采用SPSS 17.0 软件进行差异显著性分析和相关性分析,用Duncan 法进行多重比较。
2 结果
2.1 二倍体棱叶蒜花粉活力检测方法的比较选择
通过对不同开花物候期的花粉活力检测表明,经联苯胺α -萘酚处理,有活力和无活力花粉均被染成淡黄色或淡红色;经TTC 处理,有活力和无活力花粉均无着色;经醋酸洋红处理,有活力和无活力花粉均呈现红色;经I2-KI 处理,花粉在极短的时间内由黄色变成金黄色或黄褐色,着色和变色的速度都非常快,肉眼无法观察区分有活力花粉和无活力花粉;经红墨水染色,有活力和无活力花粉被染成淡红色或无着色。研究结果表明,联苯胺α - 萘酚法、TTC法、醋酸洋红法、I2-KI、孢粉染色法和红墨水法在测定棱叶蒜花粉活力过程中,各处理与对照之间无显著差异(P >0.05),没有获得可以鉴定花粉活性的有效特征,无法区分和判断花粉是否存在活力,因而不适宜于二倍体棱叶蒜的花粉活力检测。
经MTT 处理后,有活力的花粉被染成蓝色或淡蓝色,而无活力的花粉与对照均未被染色。雅玛里克山居群的棱叶蒜在开花始期、开花盛期和开花末期花粉的平均寿命均在60 h 之内。不同开花物候期的花粉活力变化规律一致,居群开花始期、盛期和末期的平均花粉活力分别为6.25%、6.60%和6.50%,方差分析结果表明,不同开花物候期相同散粉时间的花粉活力无显著差异(P >0.05)。因此,相比其他6 种方法,MTT 花粉活力检测方法可以显著区分和判断花粉是否存在活力,适宜于棱叶蒜的花粉活力检测(图1)。
2.2 二倍体棱叶蒜不同开花物候期的花粉活力变化特征
采用MTT 法检测雅玛里克山居群棱叶蒜的花粉活力(表1)。方差分析结果表明,随着散粉后时间的延长,花粉活力下降,花粉着色由深蓝色至淡蓝色到无色,花粉活力存在显著差异(P <0.05)。
在居群开花始期0 -4 h 花粉活力逐渐升高,散粉1 和4 h 的花粉着色率分别为51.66%和80.17%。平均花粉活力超过50%的时间为花药散粉后的1 -8 h,平均花粉活力为58.80%。散粉24 h,有活力花粉被染成淡蓝色,着色率为12.66%,与散粉36、48 和60 h的花粉着色率之间不存在显著差异(P >0.05),由此可知,二倍体棱叶蒜开花始期的有效花粉活力在24 h之内。
在居群开花盛期0 -4 h 花粉活力逐渐升高,散粉1 和4 h 的花粉着色率分别为37.03%和84.64%。平均花粉活力超过50%的时间为花药散粉后的0 -10 h,平均花粉活力为53.69%。散粉48 h,有活力花粉被染成淡蓝色,着色率为1.27%,与60 h 的花粉着色率之间不存在显著差异(P >0.05),由此可知,二倍体棱叶蒜开花盛期的有效花粉活力在48 h 之内。
在居群开花末期0 -4 h 花粉活力逐渐升高,散粉1 和4 h 的花粉着色率分别为32.83%和83.56%。平均花粉活力超过50%的时间为花药散粉后的0 -10 h,平均花粉活力为52.36%。散粉36 h,有活力花粉被染成淡蓝色,着色率为5.86%,与48 和60 h 的花粉着色率之间不存在显著差异(P >0.05),由此可知,二倍体棱叶蒜开花盛期的有效花粉活力在36 h 之内。

表1 MTT 法测定棱叶蒜花粉活力Table 1 Pollen vitality determined by MTT method %
2.3 离体萌发法测定二倍体棱叶蒜花粉萌发率与配方优化
离体萌发法的配方选择与优化试验结果表明(表2),不同浓度的硼酸和蔗糖对花粉萌发影响差异显著(P <0.05),在不含蔗糖和硼酸的培养基中,二倍体棱叶蒜的花粉萌发率仅为3.49%,在仅含有蔗糖成分的培养基中,随着蔗糖浓度的升高,花粉萌发率呈现出先升高后下降的变化趋势;在仅含有硼酸成分的培养基中,随着硼酸浓度的升高,花粉萌发率也呈现出先升高后下降的变化趋势,高浓度的蔗糖和硼酸均有抑制花粉萌发的作用,而当蔗糖和硼酸浓度分别达到100 g·L-1和80 mg·L-1时,花粉萌发率为72.70%,是二倍体棱叶蒜花粉萌发的适宜浓度,其次,配方为50 g·L-1蔗糖加80 mg·L-1硼酸以及100 g·L-1蔗糖加100 mg·L-1硼酸下的花粉萌发率超过50%。方差分析结果表明,不同浓度的硼酸和蔗糖对花粉萌发影响差异显著,蔗糖和花粉对花粉的萌发有互作效应(表3)。
2.4 离体萌发法检测二倍体棱叶蒜不同开花物候期的花粉萌发率
采用优化的培养基配方法检测雅玛里克山居群棱叶蒜的花粉萌发率(表4)。方差分析结果表明,随着散粉后时间的延长,花粉活力下降,花粉管萌发长度由长到短,花粉萌发率由高到低,花粉活力存在显著差异(P <0.05)。
在居群开花始期0 -4 h 花粉萌发率逐渐升高,花粉萌发率由散粉1 h 的32.81%至4 h 的76.97%。平均花粉萌发率超过50%的时间为花药散粉后的0 -8 h,平均花粉活力为51.64%。散粉24 h 后的萌发率下降至10.23%,且花粉管短;与散粉36、48 和60 h 的花粉萌发率不存在显著差异(P >0.05),由此可知,二倍体棱叶蒜开花始期的有效花粉萌发率在24 h 之内。
在居群开花盛期0 -4 h 花粉萌发率逐渐升高,花粉萌发率由散粉1 h 的58.03%至4 h 的81.39%。平均花粉萌发率超过50%的时间为花药散粉后的0 -10 h,平均花粉活力为51.74%。散粉24 h 后的萌发率下降至11.63%,且花粉管短;与散粉36、48 和60 h 的花粉萌发率不存在显著差异(P >0.05),由此可知,二倍体棱叶蒜开花盛期的有效花粉萌发率在24 h 之内。

表2 不同浓度蔗糖和硼酸处理下的花粉萌发率Table 2 Pollen germination percentage under different concentrations of sucrose and boric acid treatment %
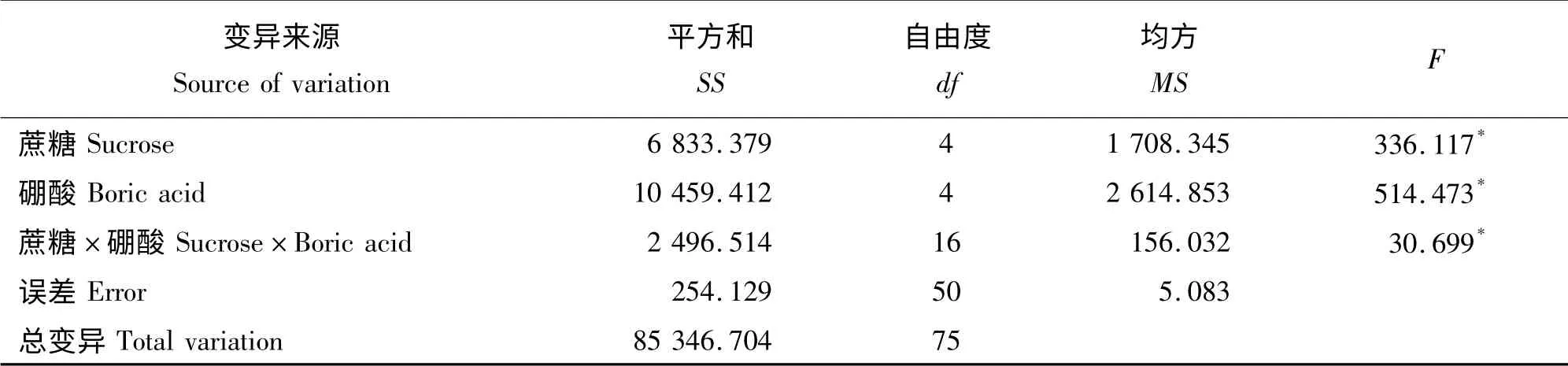
表3 花粉萌发正交试验中培养基成分互作作用的方差分析Table 3 Analysis of variance of the results of the orthogonal test of germination percentage
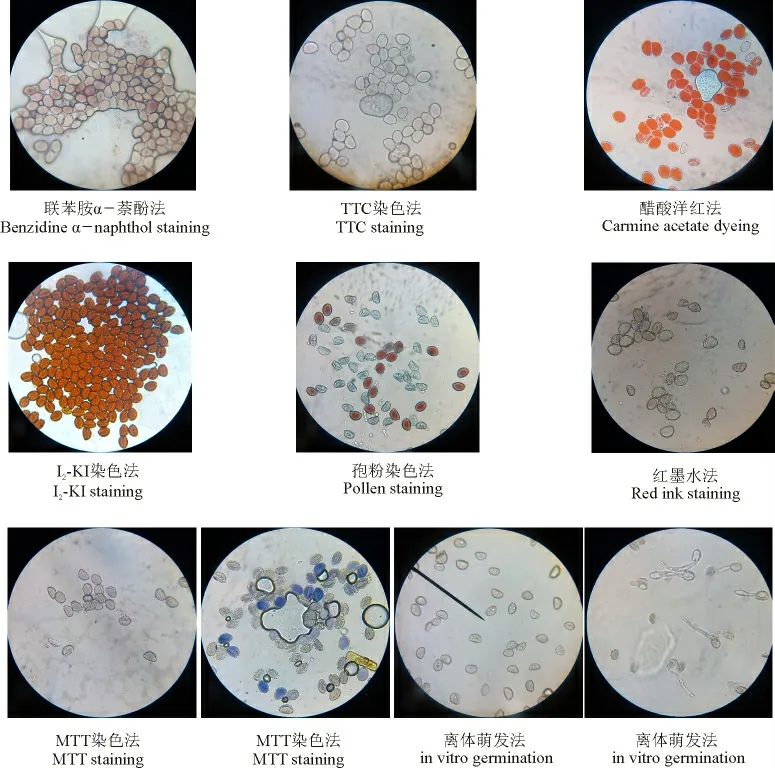
图1 二倍体棱叶蒜花粉活性的变化特征Fig.1 The variation characteristics of A. caeruleum pollen activity

表4 离体萌发法测定棱叶蒜花粉萌发率Table 4 Pollen germination percentage determined in vitro %
在居群开花末期0 -4 h 花粉萌发率逐渐升高,花粉萌发率由散粉1 h 的55.25%至4 h 的77.94%。平均花粉萌发率超过50%的时间为花药散粉后的0 -8 h,平均花粉活力为52.24%。散粉24 h 后的萌发率下降至12.50%,且花粉管短;与散粉36、48 和60 h 的花粉萌发率不存在显著差异(P >0.05),由此可知,二倍体棱叶蒜开花末期的有效花粉萌发率在24 h 之内。
2.5 花粉活力和花粉萌发率的相关性
通过对不同开花物候期的花粉活力与花粉萌发率之间的相关性分析可知,开花始期的花粉活力与萌发率之间相关系数为0.986(P <0.01),开花盛期的花粉活力与萌发率之间相关系数为0.988(P <0.01),开花末期的花粉活力与萌发率之间相关系数为0.997(P <0.01),不同开花物候期的花粉活力与花粉萌发率之间存在极显著正相关(P <0.01)。MTT 检测法与离体萌发法均能较好地检测花粉活性。
3 讨论与结论
花粉生活力是杂交育种前评估花粉活性的一个重要手段,花粉活力测定结果的准确性往往决定了细胞学试验和杂交育种的成败。不同植物花粉的自然寿命不同,花粉生活力检测方法存在差异[14-16]。利用染色法进行花粉生活力的测定,具有快速简便的优点,但不同植物种类的花粉有其自身结构特性和花粉内含物不同等原因,不同植物的花粉选择不同的染色方法[15],有醋酸洋红法[9]、TTC 染色法或I2-KI 染色法[15]、次甲基蓝或联苯胺α-萘酚染色法等[16]。
本研究表明,二倍体棱叶蒜具有较强的花粉活力检测方法的选择性,供试的7 种花粉活力检测方法中仅有MTT 法适用于二倍体棱叶蒜花粉活力的测定,这与葱属植物中高葶韭(A. obliquum)的检测结果存在差异,高葶韭花粉检测的适合方法有TTC 染色法、MTT 染色法和联苯胺染色法[9],这可能与花粉孢壁的厚度、氧化还原酶活性强弱、花粉内含物物质种类和含量、花粉细胞原生质膜透性等有关[16]。目前,适用于大多数植物花粉生活力检测法仍然是离体萌发法[17],该方法同样适用于二倍体棱叶蒜,本研究表明,其最适配方为100 g·L-1的蔗糖+80 mg·L-1的硼酸,这与高葶韭花粉最适培养基配方100 g·L-1蔗糖+50 mg·L-1硼酸的研究结果类似[9]。
由MTT 法和离体萌发法检测的花粉生活力结果可知,相同开花物候期和散粉时间的花粉活力多数高于花粉萌发率,说明能被染色的二倍体棱叶蒜花粉只是具有萌发的潜力,但在实际培养条件下不一定都能萌发。这一结果在二倍体香蕉(Musa nanalour)[17]、苹果(Malus pumila)[18]和大花三色堇(Viola wittrockiana)[19]中也有报道。MTT 染色法与花粉线粒体中的琥珀酸脱氢酶活性有关,能够影响二倍体棱叶蒜花粉中琥珀酸脱氢酶活性的环境因子在某种程度上会影响花粉萌发,同时,蔗糖浓度、硼离子浓度以及温度和pH值等因子都会影响花粉萌发,共同决定了花粉活力和寿命,这为进一步在人工控制条件下延长二倍体棱叶蒜花粉活力提供了参考。
杂交育种过程中要充分考虑开花物候期和采用的花粉育性的检测方法。综合各开花物候期花粉活力变化特征,分布于雅玛里克山的二倍体棱叶蒜居群超过50%的花粉活力和花粉萌发率的时间在8 h 以内。通过MTT 法检测的棱叶蒜的有效花粉活力的时间在开花盛期具有48 h,长于开花始期和开花末期的36 h,而离体萌发法检测出维持正常的花粉萌发率在不同开花物候期均表现出24 h,综合比较两种测定方法,为获得田间杂交成功,建议授粉时间应控制在花药散粉后0~8 h,最佳授粉时间选择0 ~4 h,花粉离体保存时间不超过24 h。
花粉活力与萌发率的相关性分析表明,雅玛里克山居群的二倍体棱叶蒜同一花期的花粉活力与萌发率极显著相关,花粉活力与花粉萌发率变化趋势相同,都存在两个峰值,这可能是为了适应环境的变化,增加授粉成功机率的一种自我保护机制。
植物的花粉活力由遗传基因决定,不同植物种类的花粉活力存在显著差异[20],长寿命花粉的植物在开花前、整个花期至单花花期结束时花粉均具有活力[10,21],短寿命的花粉活力约几小时不等[13,22-23],二倍体棱叶蒜花粉寿命在60 h 以内,花刚一开放花粉就已具有较高的活力,花药散粉后的1 ~6 h 的花粉活力或萌发率维持在50%左右;且存在两个活力高峰,48 h后基本无活力或萌发率,属于短寿命花粉。这与野生葱属植物中的高葶韭[9]、蒙古韭(A. mongolicum)、碱韭(A. polyrhizum)和细叶韭(A. temuissimum)[10]的研究结果一致,高葶韭花粉高于50%的活力或萌发率只能维持2 ~10 h,48 h 后失去活力[9];蒙古韭、碱韭和细叶韭的花粉活力在刚开花时较强,但下降较快,8 h后花粉基本失去活力[10],花粉均可育,也属于短寿命花粉。本研究通过对雅玛里克山分布的野生二倍体棱叶蒜居群的不同开花物候期的花粉活力和萌发率测定为今后该资源的种质创新奠定了理论基础。
[1] Igic B,Kohn J R.The distribution of plant mating systems:Study bias against obligately outcrossing species[J]. Evolution,2006,60(5):1098-103.
[2] Dafni A,Firmage D.Pollen-viability and longevity:Practical,ecological and evolutionary implications[J].Plant Systematics and Evolution,2000,222(1-4):113-132.
[3] 张大勇.植物生活史进化与繁殖生态学[M].北京:科学出版社,2004:322-323.
[4] Xu J M,Kamelin R V.Allium Linnaeus[A].Wu Z Y,Raven P H.Flora of China[M].Beijing:Science Press and Missouri Botanical Garden Press,2000,24:73-202.
[5] 中国科学院植物研究所.中国高等植物.第十三卷[M].青岛:青岛出版社,2002:169-170.
[6] Krzymińska A,Gawłowska M,Wolko B,Bocianowski J. Genetic diversity of ornamental Allium species and cultivars assessed with isozymes[J].Journal of Applied Genetics,2008,49(3):213-220.
[7] 杨帆,林辰壹,席延坡,张顺,杨天灵. 葱属植物棱叶蒜的形态性状与核型特征[J]. 植物遗传资源学报,2014,15(6):1262-1269.
[8] 刘盛梅.乌鲁木齐县农作物抗旱对策研究[J].现代农业科技,2012(20):237-244.
[9] 林辰壹,韩文娟,祁晨霞,叶强.新疆高葶韭雄蕊开花动态特征与花粉育性研究[J].草业科学,2012,29(4):561-566.
[10] 赵金花,李青丰,那仁图雅,斯琴.3 种野生葱属植物花粉活力和柱头可授性研究[J].草业科学,2010,27(4):93-96.
[11] 李和帅,王承民,王波超,范海阔.我国几个主要椰子品种花粉生活力研究[J].江西农业学报,2013,25(29):11-14.
[12] Rodriguez-Riano T,Dafni A.A new procedure to assess pollen viability[J].Sexual Plant Reproduction,2000,12(4):241-244.
[13] 汪妮,张志轩,董自梅.韭菜花粉生活力测定方法的筛选研究[J].长江蔬菜,2009(2):53-55.
[14] 左丹丹,明军,刘春,王丽娜.植物花粉生活力检测技术进展[J].安徽农业科学,2007,35(16):4742-4745.
[15] 王钦丽,卢龙斗,吴小琴,陈祖铿,林金星.花粉的保存及其生活力测定[J].植物学通报,2002,19(3):365-373.
[16] 马诗钰,周兰英,蒲光兰,赖腾跃,蔡利娟,王波.贴梗海棠花粉生活力与贮藏性研究[J].北方园艺,2014(16):81-85.
[17] 邓彪,盛鸥,魏岳荣,李大志,高慧君,邝瑞彬,易干军,Agustin B.二倍体香蕉种质资源花粉活力检测方法的筛选[J].分子植物育种,2014,12(5):1011-1017.
[18] 徐臣善.苹果花粉萌发生理影响因子的研究[J].中国农学通报,2011,27(31):163-167.
[19] 穆金艳,杜晓华,贾文庆,刘会超,黄伶俐.大花三色堇花粉生活力测定及贮藏方法[J].西北农业学报,2013,22(9):158-162.
[20] 胡适宜.被子植物胚胎学[M].北京:高等教育出版社,1982:20-60.
[21] 王彩霞,田敏,李全健.白及的花部特征与繁育系统[J].园艺学报,2012,39(6):1159-1166.
[22] 马建琴,郭太君,陈少鹏,赵仁林.野生细叶百合开花生物学研究[J].北方园艺,2013(23):88-92.
[23] 郭红超,严成,魏岩.木地肤的开花动态与花粉活力及柱头可授性研究[J].草业科学,2014,23(4):87-93.
