沙打旺黄矮根腐病菌钙调素基因的克隆
2015-04-08刘建利李彦忠
刘建利,李彦忠
(1.草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州730020;
2.北方民族大学生物科学与工程学院,宁夏 银川750021;3.中国农业科学院草原研究所,内蒙古 呼和浩特010010)
沙打旺(Astr agal us adsur gens)是豆科(Leguminosae)黄芪属多年生优质牧草,因其抵御风沙能力强而得到“沙大王”的美誉。目前,制约沙打旺大面积种植的最主要原因是沙打旺埃里砖格孢(Embellisia astr agali)引起的黄矮根腐病,该病害是造成沙打旺草地退化最严重的病害之一[1],该病属系统性病害,严重威胁沙打旺草地的生产力、使用寿命、草品质及种子产量[2]。植物病原真菌与宿主植物之间信号识别及传导,在病害发生中发挥着重要的作用,因此,病原真菌与植物互作过程中的信号转导机制及其调控已成为分子植物病理学研究的热点。钙调素是钙离子介导的信号通路中重要的分子,研究表明,钙调素参与调控许多植物病原真菌的交配、细胞分化、有丝分裂、致病性、分生孢子形成及萌发、色素合成、毒素的产生以及侵染过程[3]。到目前为止,绿僵菌(Metar hizin m anisopliae)[4]、苜蓿炭疽菌(Colletotrichu m medicaginis)[5]、巴 西 副 球 孢 子 菌(Par acoccidioides br asiliensis)[6]、埃默森小芽枝霉(Bl astocl adiell a emersonii)[7]、构巢曲霉(Asper gill us nidul ans)[8]、白假丝酵母(Candida albieans)[9]、盘基网柄菌(Dictyosteliu m discoideu m)[10]、致病疫霉(Phytophthor a inf estans)[11]、脉 胞 霉(Neurospor a cr assa)[12]、米曲霉(Asper gill us or yzae)[13]、荚膜组织孢浆菌(Histopl asma capsul atum)[14]、三 叶 草 刺 盘 孢 菌(Colletotrichu m trif olii)[15]、盘 长 孢 状 刺 盘 孢 菌(Colletotrichum gloeosporioides)[16]、稻 瘟 病 菌(Magaporthe grisea)[17]等许多真菌钙调素编码基因已被克隆。本研究旨在从沙打旺埃里砖格孢中克隆钙调素编码基因序列,为研究其在致病中的功能奠定基础。
1 材料与方法
1.1 材料与试剂
沙打旺黄矮根腐病菌株E.astr agali NHLZU0408来自于兰州大学草地农业科技学院草地保护所。
主要试剂有E.Z.N.A Fungal DNA Mini Kit(美国Omega Bio-Tek,Inc.公 司),Pure Plasmid Mini Kit(康为世纪生物科技有限公司),Ultrapure RNA Kit超纯RNA 提取试剂盒(康为世纪生物科技有限公司),高保真酶Pri mer STAR@HS(Per-mix)[宝生物工程(大连)有限公司]TransScriptⅡOne-Step g DNA Removal and c DNA Synt hesis Super Mix,PCR Super Mix,T 载体、Blunt-T 载体(北京全式金生物技术有限公司),其他试剂购自天津市大茂化学试剂厂。
1.2 主要仪器
PCR 仪(S1000型,Bio-Rad公司),台式高速冷冻离心机(3 K30型,Sig ma公司),空气浴恒温摇床(HZ-9210 K 型,江苏太仓市实验设备厂),电子天平(FA2104B,上海精密科学仪器有限公司),电泳系统(DYY-21型,北京六一仪器厂),凝胶成像系统(Gel Doc XR 型号,美国Bio-Rad公司)。
1.3 试验方法
1.3.1 基因组DNA、总RNA 提取及反转录 将真菌接种于PDA 液体培养基中,培养7 d,过滤菌丝体后,置于研钵中加液氮研磨后,DNA 提取步骤按照E.Z.N.A Fungal DNA Mini Kit提取试剂盒说明书进行,RNA 提取步骤按照Ultrapure RNA Kit超纯RNA 提取试剂盒说明书进行,1%琼脂糖电泳检测。反转录TransScriptⅡOne-Step g DNA Removal and c DNA Synthesis Super Mix按照说明书进行。
1.3.2 钙调素基因同源片段克隆 简并引物[18]:
CAL-228F,5′-GAGTTCAAGGAGGCCTTCTCCC-3′;CAL-737R,5′-CATCTTTCTGGCCATCATGG -3′。引物由生工生物工程(上海)股份有限公司合成。PCR反应体系组成:2×PCR Super Mix(Mg2+plus)10 μL,引物(10μmol·L-1)各1μL,DNA 模板1μL,水补足使总体积为20μL。PCR 条件参考Pryor和Bigelow[18],扩增产物连接T 载体,委托生工生物工程(上海)股份有限公司测序。
1.3.3 5′端和3′端的克隆
5′Hi-TAIL PCR 特 异 性 引 物 SPCa M51:ATCTCCGGCCCACGCAATCGGCAAAG;SPCa M52:ACGATGGACTCCAGTCCGGCCCTACAACACC ACGACATGCACCGCACTC;SPCa M53:CGACAACAACGGCACCATTGACTTCCC;
3′Hi-TAIL PCR 特 异 性 引 物 SPCa M31:ATCTCCGGCCCACGCAATCGGCAAAG;SPCa M32:ACGATGGACTCCAGTCCGGCCCTACAACACCAACATGCACCGCACTC;SPCa M33:CGACAACAACGGCACCATTGACTTCCC;Hi-TAIL PCR 随机简并引物(LAD系列)和3轮PCR条件参照Liu等[19]的方法。
1.3.4 钙调素基因全长的克隆
全长引物 Ca MF:AACACGACAGCCTGCCCACC;Ca MR:CGTCACTCGTCGCCGTTA
以1μL总DNA、c DNA 为模板,PCR 反应体系组成:2×Pri mer STAR®HS(Per mix)10μL,引物(10μmol·L-1)各1μL,模板1μL,水补足使总体积为20μL。PCR 条件94℃预变性5 min,94℃变性1 min,56 ℃退火1 min,72 ℃延伸2 min,30个循环,72 ℃后延伸5 min。
2 结果与分析
2.1 保守片段克隆
以基因组DNA 为模板,采用CAL-228F/CAL-737R引物,PCR扩增获得约539 bp片段(图1),测序后BLAST比对结果显示为钙调素基因部分片段。

图1 用简并引物扩增沙打旺黄矮根腐病菌钙调素基因部分同源片段电泳图Fig.1 Result of cal modulin part conservative sequence of E.astr agali using degenerate PCR
2.2 5′端片段和3′端片段的克隆
采用Hi-TAIL PCR 技术分别克隆沙打旺黄矮根腐病菌钙调素基因5′端和3′端片段,经过3 轮PCR,LAD1~LAD5所有5个随机引物菌扩增出条带,5′端 部 分 片 段 取LAD3 的 第3 轮PCR 扩 增 的300-400 bp产物回收测序,获得344 bp序列(图2),含特异性引物位点,其前面部分序列也与同源片段中序列一致,BLAST 比对后与其他物种钙调素基因高度一致;3′端部分片段取LAD3 的第3 轮PCR 约700~800 bp产物回收测序,获得729 bp序列(图2),含特异性引物位点,其后面部分序列也与同源片段中序列一致,BLAST 比对后与Gen Bank中其他物种钙调素基因高度一致。
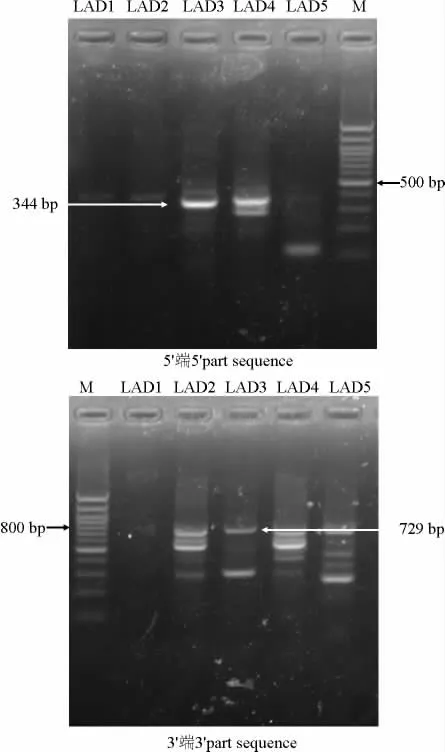
图2 Hi-TAIL技术克隆沙打旺黄矮根腐病菌钙调素基因5′和3′端部分片段电泳图Fig.2 Result of agarose gel-ectrophoresis of cal modulin 5′and 3′part sequence of E.astr agali using Hi-TAIL PCR
2.3 全长基因克隆
将保守片段、5′端片段和3′端片段电子拼接,获得1 290 bp长度片段,其中编码区840 bp。设计全长引物,以基因组DNA 和c DNA 为模板,扩增获得沙打旺黄矮根腐病菌DNA 和c DNA 编码区全长,测序,获得840 bp的DNA 和450 bp的c DNA 编码区全长序列(图3),与电子拼接结果一致,BLAST比对后与Gen Bank中其他物种钙调素基因高度一致。DNA 全长序列中包括4个内含子,均符合GTAG 规则(图4)。c DNA 编码序列翻译149个氨基酸的多肽,BLAST 比对也与SWISS-PROT 中其他物种钙调素蛋白序列高度一致。

图3 黄矮根腐病菌钙调素基因DNA和c DNA全长序列克隆电泳图Fig.3 Result of cal modulin DNA complete coding sequence of E.astr agali

图4 沙打旺黄矮根腐病菌钙调素DNA和c DNA编码基因比对图Fig.4 Align ment of cal modulin DNA and c DNA sequence of E.astr agali
3 讨论
常规克隆基因的方法是从c DNA 到DNA,即先以mRNA 反转录的c DNA 为模板,利用简并引物扩增获得目的基因部分区段,然后利用RACE 技术获得5′端和3′端序列,最后拼接全长,再设计全长引物分别以c DNA 和DNA 为模板扩增获得全长c DNA 序列和DNA 序列。该方法的关键环节在于c DNA 末端快速扩增(Rapid Amplification of c DNA Ends,RACE)技术,试剂盒费用高,对mRNA质量要求也较高,费时,技术难度也大。Hi-TAIL PCR 技术是一种根据已知序列基因获得其侧翼序列的基因组步移新技术,以DNA 为模板,需4~5条通用引物和2~3条特异引物,3 轮PCR 即可获得结果,1 d可获得试验结果,已被广泛用于克隆启动子或探知T-DNA 突变体插入位点。既然Hi-TAIL PCR 是一种从已知序列获得近旁未知序列的方法,从理论上分析,Hi-TAIL PCR 技术可以用于植物病原真菌基因的克隆[20-21]。本研究在得到同源区段的基础上,成功利用Hi-TAIL PCR 技术克隆到沙打旺黄矮根腐病钙调素编码基因全长,证明Hi-TAIL PCR 技术可以用于植物病原真菌基因克隆。相比较RACE技术,其以DNA 为模板,最大的优点是操作简单、成本低,在已知片段GC 含量较高的时候,成功机率尤为高。
对钙调素参与真菌致病性调控研究已进行了多年。最早研究者用钙调素特异性抑制剂阻断信号通路试验推测钙调素参与真菌致病性调控,例如,钙调素参与玉米小斑菌(Cochliobol us heterostrophus)[22]、稻梨孢(Magnaporthe grisea)[23]、盘长孢状刺盘孢菌[24]、三叶草刺盘孢菌[15]、桃褐腐病菌(Monilinia f r ucticol a)[25]、水稻胡麻斑菌(Cochliobol us miyabeanus)[26]、胶孢炭疽菌(Colletotrichu m gloeosporioides)[27]附着胞的形成;钙调素抑制白假丝酵母由Y 型(yeast)向M 型(myceliu m)的形态转化过程[28];钙调素参与尾孢菌(Cercospor a nicotianae)[29]毒素的合成。近年来,采用反向遗传学和正向遗传学手段对钙调素参与调控的研究也陆续展开。2000年,War war等[30]通过反义表达技术(Antisense Expression)也证实,钙调素基因参与稻梨孢和盘长孢状刺盘孢附着胞的形成。2005 年,Kraus等[17]利用一个 新型隐球菌(Cr y ptococcus neof or mans)钙调素插入突变体,发现钙调素调控新型隐球菌形态构建和温度适应性及致病性。Liu和Kolattukudy[31]、St Leger等[4]把钙调素基因启动子与GFP构建载体,转到其稻梨孢基因组中,发现随附着胞的不断形成,钙调素基因的表达量迅速增加。Dick man等[5]利用钙调素特异性探针或引物,发现抑制钙调素基因的表达,苜蓿炭疽仍能形成附着胞,但会使形成率降低。这些研究结果都表明,钙调素参与了病原真菌菌丝生长、孢子萌发、附着胞形成或致病性的调控等。下一步将采用基因敲除或RNAi技术研究钙调素对沙打旺黄矮根腐病菌生长、发育、代谢及致病性的调控。
致谢:该论文是第二届全国草业生物技术大会评选出的优秀论文,并得到中国草业生物技术专业委员会提供的版面费支持。
[1] Li Y Z,Nan Z B.A new species,Embellisia astr agali sp.nov.,causing standing milk-vetch disease in China[J].Mycologia,2007,99(3):406-411.
[2] Li Y Z,Nan Z B,Hou F J.The roles of an Embellisia sp.causing yellow stunt and root rot of Astragal us adsur gens and other f ungi in the decline of legu me pastures in norther n China[J].Australasian Plant Pathology,2007,36(4):397-402.
[3] Lengeler K B,Davidson R C,D’souza C,Harashi ma T,Shen W C,Wang P,Pan X,Waugh M,Heit man J.Signal transduction cascades regulating f ungal develop ment and vir ulence[J].Microbiology and Molecular Biology Reviews,2000,64(4):746-785.
[4] St Leger R J,Roberts D W,Staples R C.Calcium and cal modulin-mediated protein synthesis and protein phosphorylation during ger mination,growth and protease production by Metar hizium anisopliae[J].Journal of General Microbiology,1989,135(8):2141-2154.
[5] Dick man M B,Buhr T,War war V,Tr uesdell G,Huang C.Molecular signals during the early stages of alfalfa ant hracnose[J].Canadian Jour nal of Botany,1995,73(S1):1169-1177.
[6] de Car valho M J A,Amori m Jesuino R S,Daher B S,Silva-Pereira I,de Freitas S M,Soares C,Felipe M S S.Functional and genetic characterization of cal modulin fro m t he di mor phic and pathogenic f ungus Par acoccidioides br asiliensis[J].Fungal Genetics and Biology,2003,39(3):204-210.
[7] Si mño R d C G,Go mes S L.Str ucture,expression,and f unctional analysis of the gene coding f or cal modulin in the Chytridiomycete Blastocladiell a emersonii[J].Jour nal of Bacteriology,2001,183(7):2280-2288.
[8] Ras mussen C D,Means R L,Lu K P,May G S,Means A R.Characterization and expression of the unique cal modulin gene of Asper gill us nidul ans[J].Journal of Biological Chemistry,1990,265(23):13767-13775.
[9] Saporito S M,Sypherd P S.The isolation and characterization of a cal modulin-encoding gene(CMD)from the dimorphic f ungus Candida albicans[J].Gene,1991,106(1):43-49.
[10] Liu T,Williams J,Clar ke M.Inducible expression of cal modulin antisense RNA in Dictyosteliu m cells inhibits the co mpletion of cytokinesis[J].Molecular Biology of t he Cell,1992,3(12):1403-1413.
[11] Pieterse C M,Verbakel H M,Spaans J H,Davidse L,Govers F.Increased expression of the cal modulin gene of the late blight f ungus Phytophthor a inf estans during pathogenesis on potato[J].Molecular Plant-Microbe Interactions,1993,6(2):164-172.
[12] Melnick M B,Melnick C,Lee M,Wood ward D O.Str ucture and sequence of the cal modulin gene fro m Neurospor a cr assa[J].Biochi mica et Biophysica Acta(BBA)-Gene Structure and Expression,1993,1171(3):334-336.
[13] Yasui K,Kitamoto K,Go mi K,Ku magai C,Ohya Y,Tamura G.Cloning and nucleotide sequence of t he cal modulin-encoding gene(c md A)fro m Asper gill us or yzae[J].Bioscience,Biotechnology,and Biochemistry,1995,59(8):1444-1449.
[14] El-Rady J,Shearer G.Isolation and characterization of a cal modulin-encoding c DNA from the pathogenic f ungus Histopl asma capsul atum[J].Medical Mycology,1996,34(3):163-169.
[15] War war V,Dick man M B.Effects of calcium and cal modulin on spore ger mination and appressorium develop ment in Colletotrichu m trif olii[J].Applied and Environ mental Microbiology,1996,62(1):74-79.
[16] Ki m Y K,Li D,Kolattukudy P E.Induction of Ca2+-cal modulin signaling by hard-surface contact primes Colletotrichum gloeosporioides conidia to ger minate and f or m appressoria[J].Jour nal of Bacteriology,1998,180(19):5144-5150.
[17] Kraus P R,Nichols C B,Heit man J.Calciu m-and calcineurin-independent roles for cal modulin in Cr yptococcus neof ormans morphogenesis and high-temperature growth[J].Eukaryotic Cell,2005,4(6):1079-1087.
[18] Pryor B M,Bigelo w D M.Molecular characterization of Embellisia and Ni mbya species and their relationship to Alter naria,Ulocl adium and Stemphylium[J].Mycologia,2003,95(6):1141-1154.
[19] Liu Y G,Chen Y L.High-efficiency ther mal asy mmetric interlaced PCR for amplification of unknown flanking sequences[J].Biotechnology Techniques,2007,43:649-656.
[20] 刘自勇,孙宪昀,曲音波.斜卧青霉114-2 cbh1基因的TAIL-PCR克隆及与抗阻遏突变株JU-A10的比较[J].微生物学报,2008,48(5):667-671.
[21] 杨帆,林俊芳,苗灵凤,郭丽琼.利用TAIL-PCR 和RT-PCR 技术克隆云芝植酸酶基因[J].应用与环境微生物学报,2006,12(5):618-622
[22] 赵俊霞,陈颖,王立安.钙信号途径参与小斑病菌致病过程的调控[J].微生物学通报,2005,32(4):1-4.
[23] 王立安,王源超,李昌文,郑小波.Ca2+信号途径参与稻瘟病菌分生孢子萌发及附着胞形成的调控[J].菌物系统,2003,22(3):457-465.
[24] Uh m K H,Ahn I P,Ki m S,Lee Y H.Calciu m/cal modulin-dependent signaling for prepenetration develop ment in Colletotrichu m gloeosporioides[J].Phytopathology,2003,93(1):82-87.
[25] Lee M H,Bostock R M.Induction,regulation,and role in pathogenesis of appressoria in Monilinia f r ucticola[J].Phytopat hology,2006,96(10):1072-1080.
[26] Ahn I P,Suh S C.Calcium restores prepenetration morphogenesis abolished by methylglyoxal-bis-guanyl hydrazone in Cochliobol us miyabeanus infecting rice[J].Phytopathology,2007,97(3):331-337.
[27] Ahn I P,Ki m S,Choi W B,Lee Y H.Calciu m restores prepenetration morphogenesis abolished by polyamines in Colletotrichu m gloeosporioides infecting red pepper[J].FEMS Microbiology Letters,2003,227(2):237-241.
[28] Sabie F,Gadd G.Involvement of a Ca2+-cal modulin interaction in the yeast-mycelial(YM)transition of Candida albicans[J].Mycopathologia,1989,108(1):47-54.
[29] Chung K R.Involvement of calciu m/cal modulin signaling in cercosporin toxin biosynt hesis by Cercospor a nicotianae[J].Applied and Environmental Microbiology,2003,69(2):1187-1196.
[30] War war V,Oved S,Dick man M B.Antisense expression of t he cal modulin gene fr o m Colletotrichu m trif olii i mpairs prepenetration develop ment1[J].FEMS Microbiology Letters,2000,191(2):213-219.
[31] Liu Z M,Kolattukudy P E.Early expression of the cal modulin gene,which precedes appressoriu m for mation in Magnaporthe grisea,is inhibited by self-inhibitors and requires surface attach ment[J].Journal of Bacteriology,1999,181(11):3571-3577.
