不同Waxy蛋白亚基型小麦胚乳淀粉粒度分布特征
2015-03-29刘希伟宋霄君戴忠民蔡瑞国
张 敏,刘希伟,赵 城,宋霄君,戴忠民,蔡瑞国*
(1 河北科技师范学院生命科技学院,河北 秦皇岛,066600;2德州学院生物系)
不同Waxy蛋白亚基型小麦胚乳淀粉粒度分布特征
张 敏1,刘希伟1,赵 城1,宋霄君1,戴忠民2,蔡瑞国1*
(1 河北科技师范学院生命科技学院,河北 秦皇岛,066600;2德州学院生物系)
选用11个不同Wx蛋白亚基型小麦品种,研究小麦淀粉粒数目、体积和表面积分布特征及其与直链淀粉含量之间的关系。结果表明,小麦胚乳淀粉粒的粒径范围为0.4~48 μm,且最大粒径存在品种间差异。小麦淀粉粒数目分布呈单峰曲线变化,峰值粒径小于1 μm;B型淀粉粒(粒径<10 μm)占淀粉粒数量的99%以上,A型淀粉粒(粒径≥10 μm)占淀粉粒数量不足1%。小麦淀粉粒体积分布呈双峰曲线变化,峰值粒径分别位于4.0~5.9 μm和21.7~23.8 μm,峰谷位于10 μm;A型淀粉粒占淀粉体积的40.4%~61.3%,B型淀粉粒占38.7%~59.6%。小麦淀粉粒表面积分布呈双峰或3峰曲线变化,双峰曲线的峰值粒径分别处于2.1~2.8 μm和19.8~21.7 μm,3峰曲线的峰值粒径分别为1.2,4.9,9.8 μm;B型淀粉粒的表面积(占82.0%~90.3%)显著高于A型淀粉粒(占9.7%~18.0%)。3峰曲线中的B型淀粉粒的表面积又可分为B1型(峰值粒径为1.2 μm)占表面积的46.9%~47.8%;B2型(峰值粒径为4.9 μm)占表面积的38.2%~39.5%。Waxy蛋白亚基的完全缺失对小麦淀粉粒的分布特征影响较大。表现为糯小麦的数量峰值粒径变大,1~10 μm淀粉粒数量增加,<1 μm淀粉粒数量减少,B型淀粉粒体积和表面积增加,A型淀粉粒的体积和表面积降低。Waxy蛋白亚基的部分缺失对小麦淀粉粒的分布也存在一定影响。相关性分析表明,直链淀粉含量与B型淀粉粒的体积、表面积百分数和1~10 μm淀粉粒数量百分数呈显著负相关,与A型淀粉粒的体积、表面积百分数和<1 μm淀粉粒数量百分数呈显著正相关。直链淀粉含量与A型淀粉粒的数量百分数相关不显著。
小麦;Wx蛋白亚基;淀粉;粒度分布
淀粉占小麦籽粒干质量的65%~70%[1],以淀粉粒的形式存在于胚乳中。一般认为,成熟小麦胚乳含有2种类型淀粉粒,A型淀粉粒呈圆盘或透镜状,直径为10~35 μm;B型淀粉粒呈球型或多边形,直径为1~10 μm[2]。不同基因型小麦胚乳淀粉的粒度分布存在差异,这种差异对小麦食品及非食品加工有重要影响[2,3]。研究发现,淀粉粒大小分布的改变对面团流变学特性影响较大[4,5]。可见,研究淀粉粒的分布状况将有助于揭示小麦淀粉品质差异的内在原因。
前人研究表明,A型淀粉粒数目占小麦胚乳总淀粉粒数目的3%,而其重量占胚乳淀粉重量的70%;B型淀粉粒数目占胚乳淀粉粒数目的90%以上,但其重量仅占胚乳淀粉总重量的25%~30%[3,6]。通过淀粉重量百分数或根据直径区分的淀粉粒相对数目作图后表明,小麦胚乳淀粉粒大小呈双峰曲线变化,第一个峰是B型淀粉粒,第二个峰是A型淀粉粒[2]。另有研究[7~10]表明,小麦胚乳淀粉粒大小呈三峰曲线变化,除了A型和B型淀粉粒外,还有C型淀粉粒。C型淀粉粒直径小于5 μm,占淀粉粒数量的45.7%,占质量的3.4%。前人关于淀粉粒分布的研究结果存在差异。
淀粉粒主要由直链淀粉和支链淀粉2种淀粉分子组成。在普通小麦中,控制胚乳直链淀粉合成的酶蛋白称为Waxy蛋白,它由Wx-A1,Wx-B1,Wx-D1等3个亚基组成[11],且它们对直链淀粉合成的影响程度不同[12~14]。当3个Waxy蛋白亚基全部缺失时,小麦直链淀粉含量极低,称为糯小麦。前人关于糯小麦及部分糯小麦的直链淀粉含量、膨胀势、粘度特性[12~18]、晶体特性[19]和链长分配[20]进行了研究。但关于Waxy蛋白亚基缺失后小麦淀粉粒度分布变化的研究,尚未见报道。为此,笔者试验选用11个不同Waxy蛋白亚基型小麦品种,研究小麦淀粉粒数目、体积和表面积分布特征及其与直链淀粉含量之间的关系,目的是揭示小麦淀粉粒度分布特征,探讨直链淀粉含量变化对小麦胚乳淀粉粒度分布特征的影响。
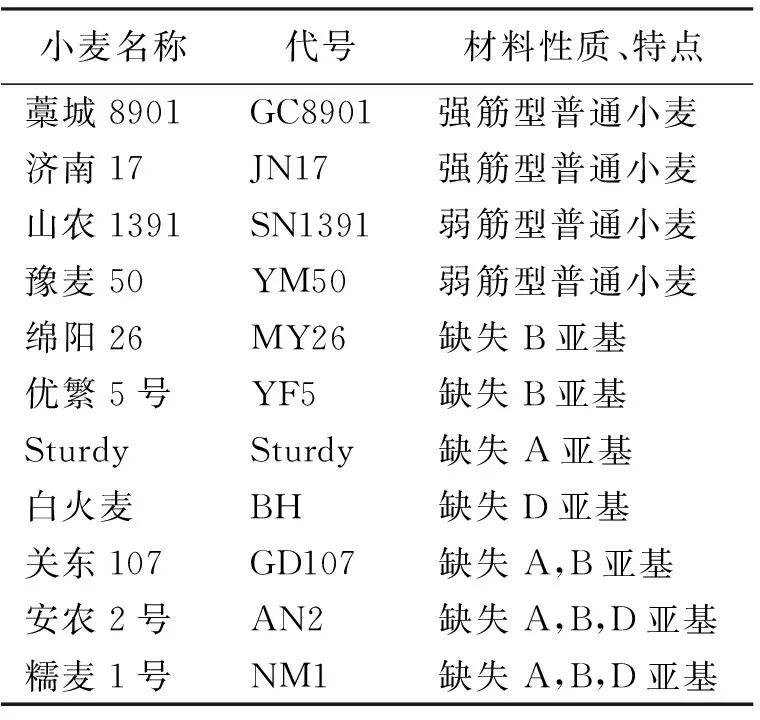
表1 11个不同Waxy蛋白亚基型小麦品种的名称及特性
1 材料与方法
1.1 试验材料
试验选用11个不同Waxy蛋白亚基型小麦品种(表1)。
试验材料分别于2013年的10月1日和2014年的10月1日播种于河北科技师范学院农学实验站。2013年试验为预备试验,主要为扩繁Waxy蛋白亚基缺失型品种。2014年试验为正式试验。小区面积为3 m×3 m=9 m2,3次重复;播种密度180 株·m-2,行距25 cm。试验田肥力均一,植株长势良好。材料分别于2014年6月25日和2015年6月24日收获,籽粒晒干保存。2年小麦籽粒均提取淀粉,测定其粒度分布和直链淀粉含量。2年结果趋势一致,本次研究以2015年收获样品的测定结果为主。
1.2 小麦籽粒淀粉提取
参考Peng[2]、Stoddard[6]和Raeker[9]的方法。具体提取方法如下:取成熟期小麦籽粒5 g,加入0.5 mol·L-1NaCl 25 mL,置于4 ℃冰箱中48 h,去除籽粒的果皮和胚,置于研钵中加入0.5 mol·L-1NaCl溶液研磨,直至有面筋析出,过滤。将未滤过物多次研磨过滤,直至滤出液用碘试剂染色不显蓝色或紫红色为止。合并滤液,3 000 r/min离心10 min,取沉淀。沉淀再分别用2 mol·L-1NaCl,20 g·L-1SDS(十二烷基硫酸钠)和2 g·L-1NaOH溶液顺序振荡冲洗,去除蛋白质,直至沉淀为白色为止。最后沉淀用丙酮振荡冲洗3遍,40 ℃烘干,磨细,贮藏备用。每个样品重复2次。
1.3 淀粉粒度分布的测定
利用LS13 320型激光粒度分析仪(贝克曼库尔特公司生产)测定,采用其通用液体样品池,所用分散液为蒸馏水。具体实验方法如下:称取0.05 g淀粉样品,加入5 mL蒸馏水振荡分散成悬浊液,得待测液。打开仪器,调节泵速为45 r/min,设运行时间为35 s,选Fraunhofertf 780z光学模型,检测仪器背景和光路中心,然后注入约1 mL待测液,待遮蔽率显示OK后开始分析。每个样品重复3~5次。
1.4 直链淀粉含量的测定
参照GB7648-87的方法,略有改动,所用样品为提纯的淀粉。每个样品重复3次。
1.5 统计分析与作图
应用DPS统计软件进行数据统计与分析;利用Excel软件和LS13 320型激光粒度分析仪自带软件进行作图。因本次试验选用品种数目较多,为更直观、清晰的显示试验结果,将所有品种分为5组,且将藁城8901设为对照,以利比较。
2 结果与分析
2.1 小麦胚乳淀粉粒的大小与数量分布
本研究结果表明,小麦胚乳淀粉粒的粒径范围为0.40~47.94 μm,且最大粒径存在品种间差异(表2)。弱筋小麦SN1391的最大粒径为47.94 μm;JN17,BH,NM1的最大粒径较小,为39.78 μm,其它品种的最大粒径接近44 μm。可以看出,Waxy的缺失与否对小麦淀粉粒的粒径分布范围影响不显著。
小麦胚乳淀粉粒的数量分布呈单峰曲线变化(图1 A1,B1,C1;图2 D1,E1),这与前人的研究结果明显不同。所有参试品种的峰值粒径均小于1 μm(表2)。AN2和NM1的峰值粒径为0.91 μm,JN17和BH的为0.83 μm,其它品种的约为0.55 μm。可见,糯小麦的峰值粒径较大。统计分析表明,A型淀粉粒(粒径≥10 μm)的数量仅占总淀粉粒数量的0.08%~0.12%,而B型淀粉粒(粒径<10 μm)的数量占总淀粉粒数量的99%以上(表3),说明小麦胚乳中的淀粉粒主要是B型淀粉粒。
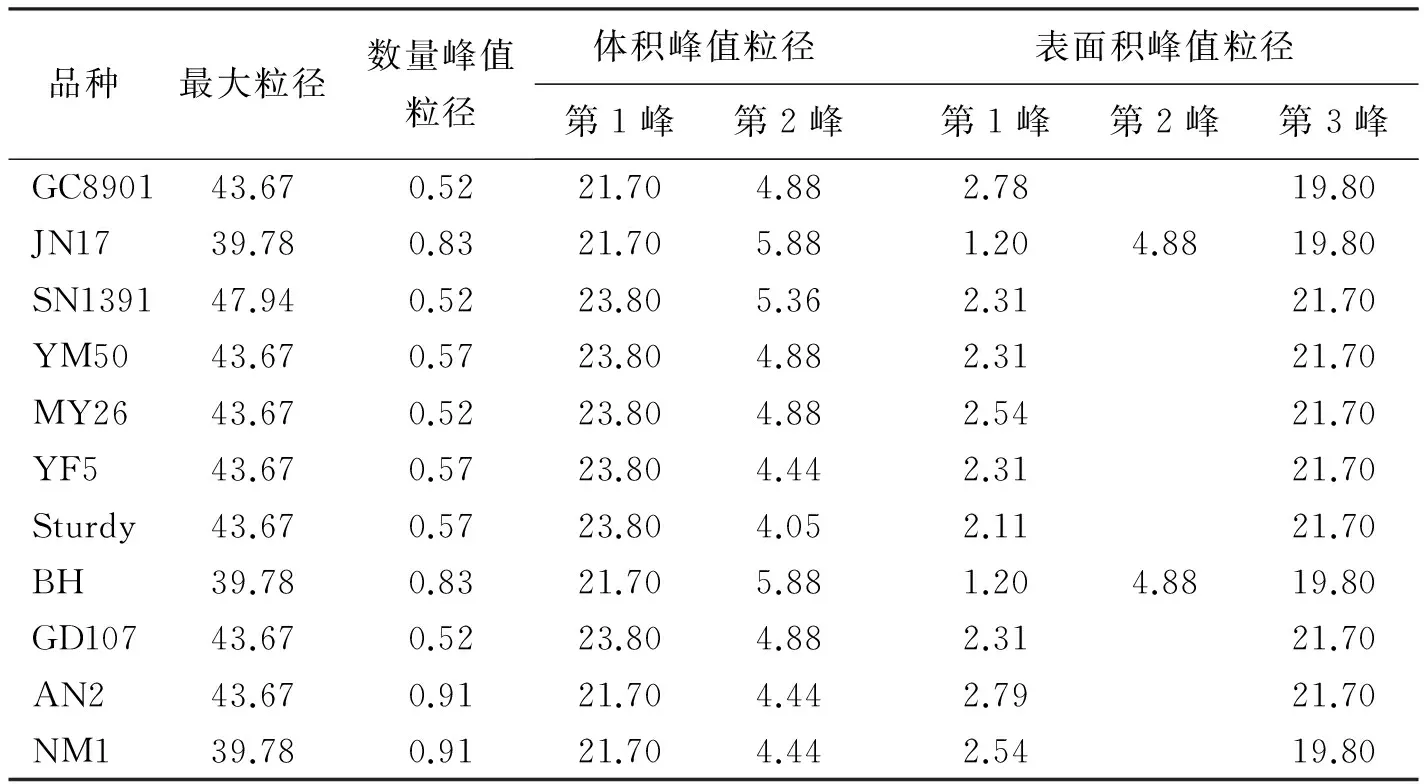
表2 小麦胚乳淀粉粒的最大粒径和其数量、体积与表面积分布的峰值粒径 μm
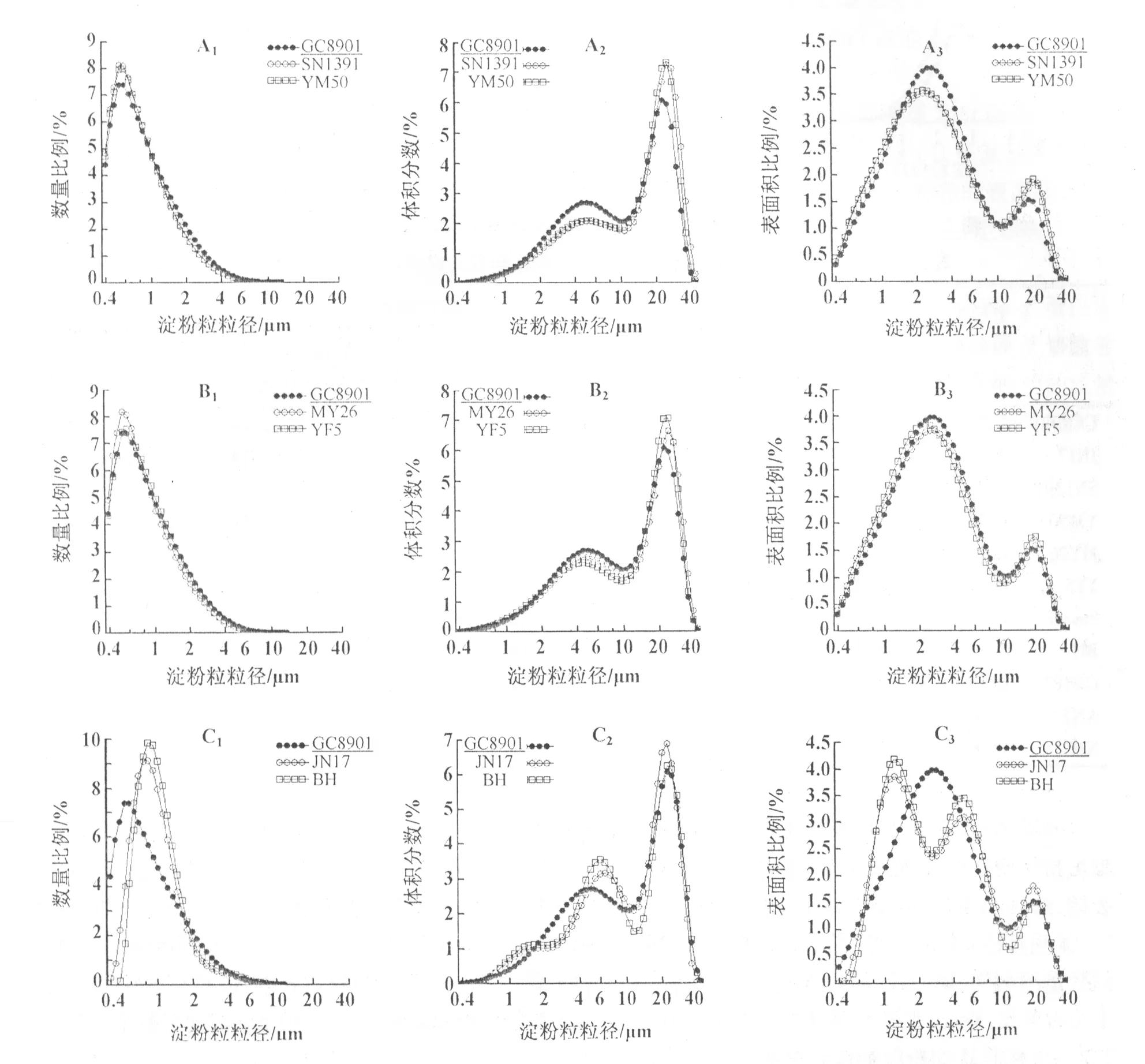
图1 小麦品种GC8901,JN17,SN1391,YM50,MY26,YF5号和BH胚乳淀粉粒的数量、体积与表面积分布
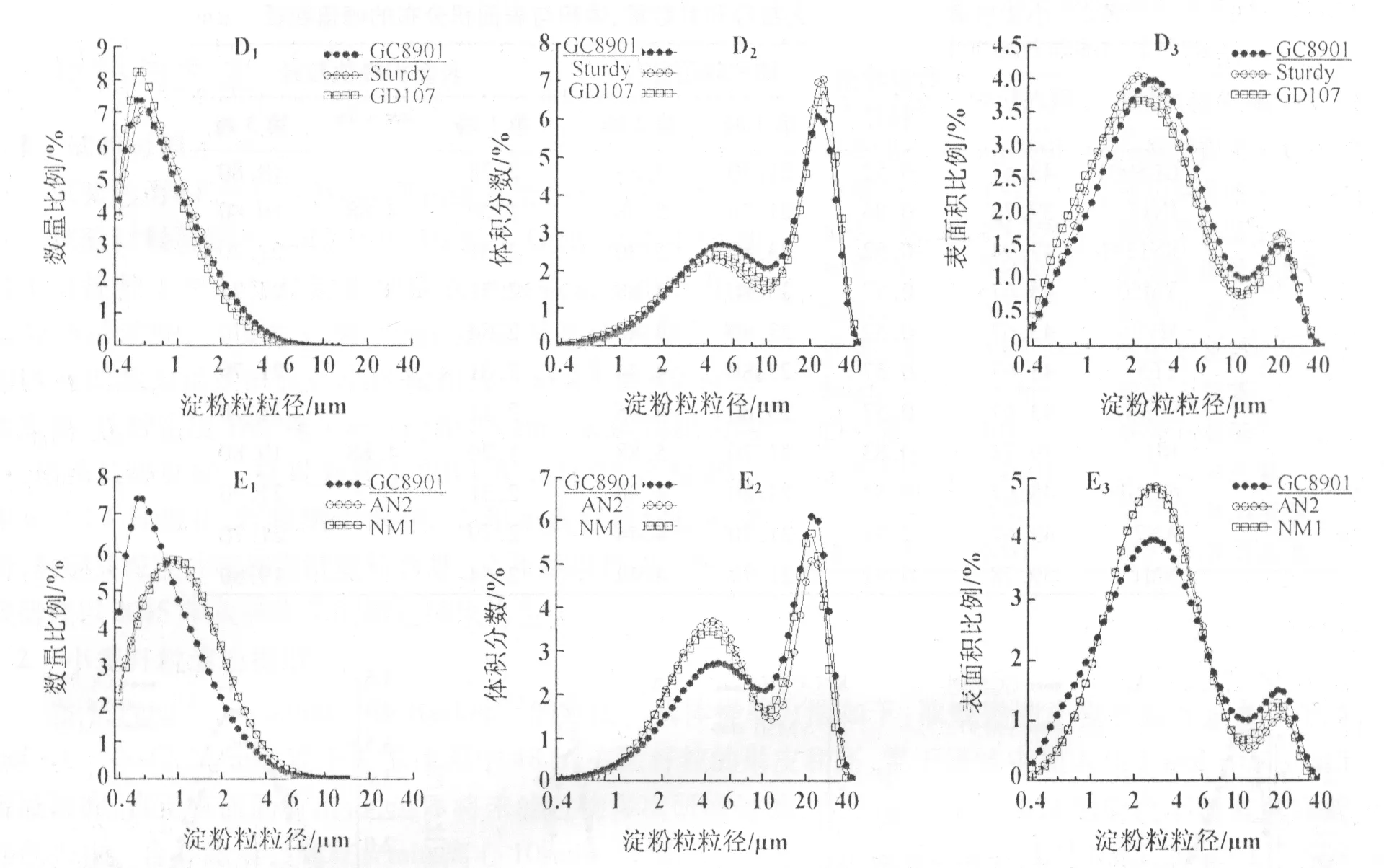
图2 小麦品种GC8901,Sturdy,GD107,AN2和NM1胚乳淀粉粒的数量、体积与表面积分布
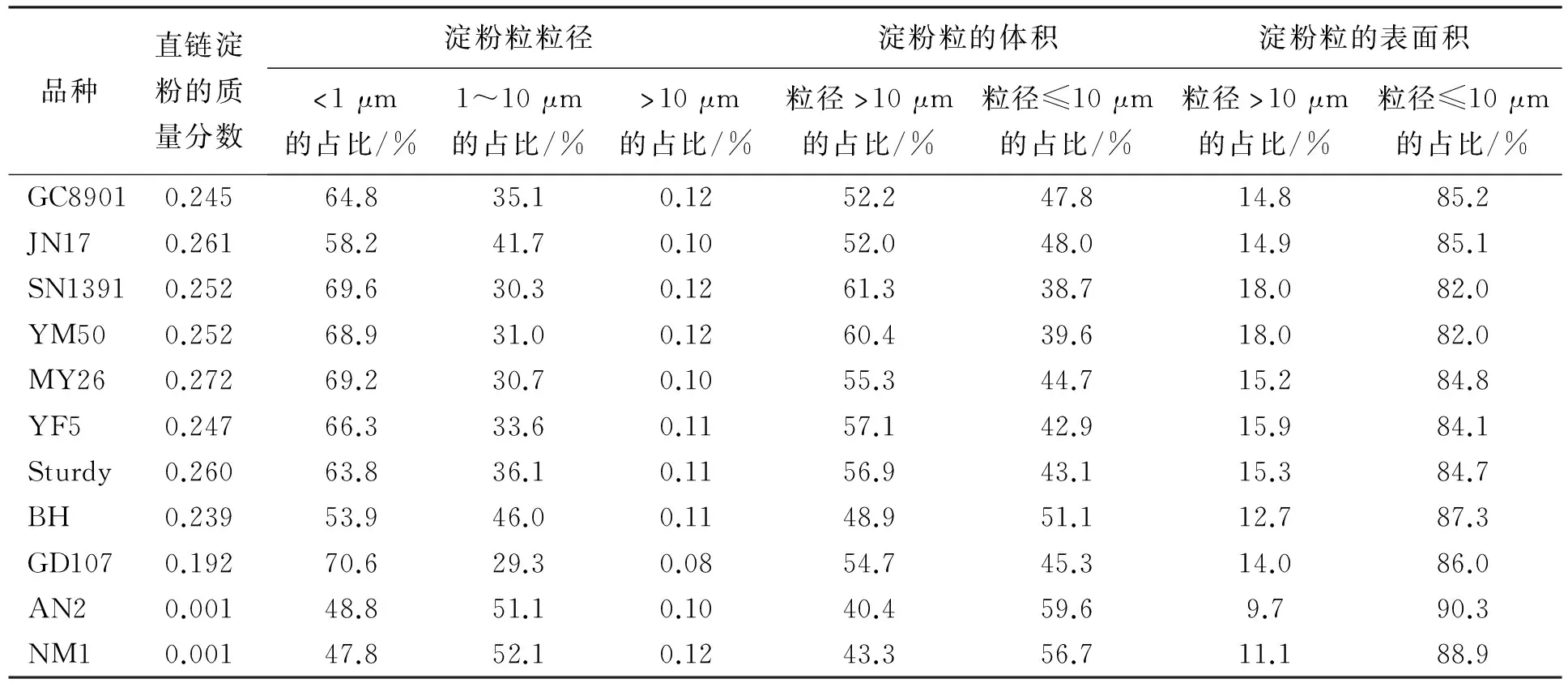
表3 小麦胚乳淀粉中直链淀粉的质量分数和淀粉粒的数量、体积与表面积分布状况
不同Waxy蛋白亚基型小麦品种的B型淀粉粒分布存在差异。为了更清楚的说明这一现象,笔者把B型淀粉粒又分为BS型淀粉粒(直径<1 μm淀粉粒)和BL型淀粉粒(直径1~10 μm淀粉粒)。统计分析表明,非糯小麦的BS型淀粉粒占淀粉粒数量的53.9%~70.6%,BL型淀粉粒占29.3%~46.0%(表3),说明直径<1 μm的淀粉粒是非糯小麦B型淀粉粒的主要组成部分。糯小麦的BS型淀粉粒约占淀粉粒数量的48%,BL型淀粉粒占51%以上(表3)。可见,糯小麦的B型淀粉粒主要以BL型淀粉粒为主。综上可以看出,Waxy亚基全部缺失后小麦胚乳中<1 μm淀粉粒数量减少,而1~10 μm淀粉粒数量增加。
2.2 小麦胚乳淀粉粒的体积分布
小麦胚乳淀粉粒的体积分布呈双峰曲线变化(图1 A2,B2,C2;图2 D2,E2)。峰值粒径分别位于4.0~5.9 μm和21.7~23.8 μm,峰谷位于大约10 μm。可见,粒径较小的峰对应B型淀粉粒,粒径较大的峰对应A型淀粉粒。
统计结果显示,小麦A型淀粉粒体积占胚乳淀粉总体积的40.4%~61.3%,而B型淀粉粒占38.7%~59.6%(表3)。进一步分析发现,不同基因型品种间A型淀粉粒的体积V分布表现为,V(弱筋小麦)>V(部分糯小麦(BH除外))>V(强筋小麦)>50%>V(糯小麦);B型淀粉粒的体积分布呈相反趋势。糯小麦B型淀粉粒的体积占56.7 %~59.6%(表3),显著高于普通小麦和部分糯小麦,说明Waxy蛋白的全部缺失后小麦B型淀粉粒体积增加;相对应而言,糯小麦A型淀粉粒体积降低。普通小麦和部分糯小麦(白火麦除外)胚乳中A型淀粉粒的体积占50%以上,而B型淀粉粒的体积低于50%。
2.3 小麦胚乳淀粉粒的表面积分布
小麦胚乳淀粉粒的表面积分布呈双峰(图1 A3,B3;图2 D3,E3)或3峰(图1 C3)曲线变化。双峰曲线的峰值粒径分别处于2.1~2.8 μm和19.8~21.7 μm;3峰曲线的峰值粒径分别处于1.2,4.9,19.8 μm。双峰曲线的峰谷和3峰曲线的1个峰谷均处于大约10 μm(图1 A3,B3,C3;图2 D3,E3),3峰曲线的另1个峰谷位于大约2.5 μm。可见,10 μm仍然可以作为小麦胚乳淀粉粒表面积分布的分界线。
统计分析表明,小麦胚乳A型淀粉粒的表面积可占总表面积的9.7%~18.0%,B型淀粉粒可占82.0%~90.3%(表3)。可见,小麦胚乳中B型淀粉粒的表面积显著高于A型淀粉粒。济南17和白火麦中的B型淀粉粒的表面积又可分为2部分,B1型(峰值粒径为1.2 μm)占表面积的46.9%~47.8%,B2型(峰值粒径为4.9 μm)占38.2%~39.5%。
不同基因型间小麦A型、B型淀粉粒的表面积S分布存在差异。A型淀粉粒表现为S(弱筋小麦)>S(B或A亚基缺失型小麦)>S(强筋小麦)>S(关东107(AB亚基缺失))>S(白火麦(D亚基缺失))>S(糯小麦);B型淀粉粒呈相反趋势。可见,Waxy蛋白亚基的完全缺失显著降低A型淀粉粒的表面积,提高B型淀粉粒的表面积;Waxy蛋白亚基的部分缺失对A型和B型淀粉粒的表面积分布存在一定影响。
2.4 小麦胚乳淀粉粒分布与其直链淀粉质量分数的相关性分析
前人研究结果表明,Waxy蛋白亚基的缺失降低小麦直链淀粉的质量分数,并且不同 Waxy 蛋白亚基对直链淀粉合成的影响程度不同[12~14]。本次研究结果表明,糯小麦淀粉中直链淀粉的质量分数极低,仅为0.001;Waxy亚基的部分缺失影响小麦直链淀粉的质量分数,且双缺失型小麦直链淀粉的质量分数低于单缺失型的(表3)。可见,不同Waxy蛋白亚基型品种小麦中的直链淀粉的质量分数差异较大。
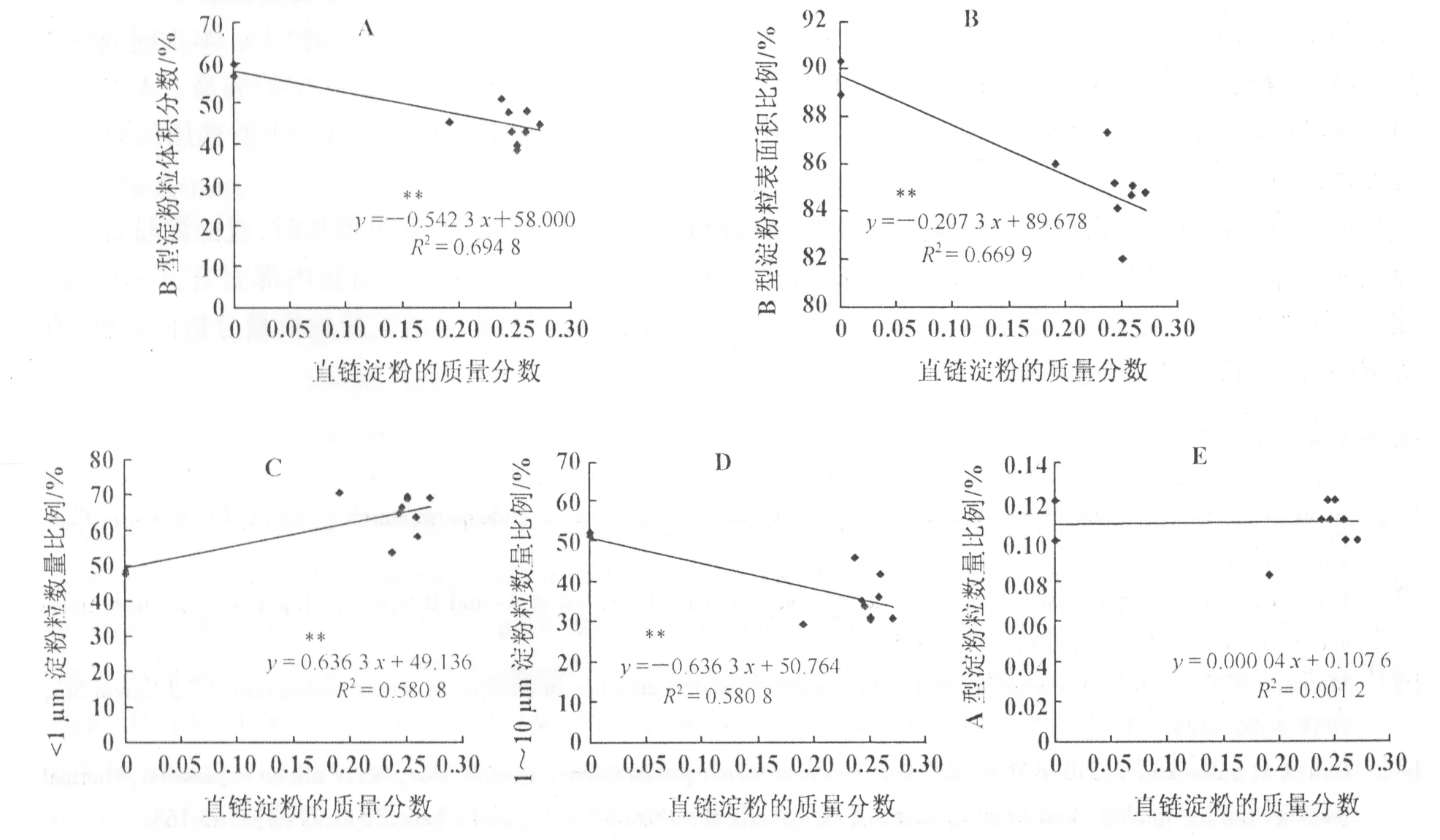
图3 直链淀粉含量和淀粉粒体积、表面积和数量分布的相关性分析(**代表0.01显著水平)
从前面的试验结果可以看出,不同Waxy蛋白亚基型品种的淀粉粒分布存在较大差异。那么,对直链淀粉的质量分数和淀粉粒分布各参数间的关系进行相关性分析,结果表明,直链淀粉的质量分数与B型淀粉粒的体积分数、表面积比例、1~10 μm淀粉粒数量比例之间呈负相关,与A型淀粉粒的体积、表面积比例、<1 μm淀粉粒数量比例之间呈正相关,且相关系数均达极显著水平(图3)。直链淀粉的质量分数与A型淀粉粒的数量比例之间相关不显著(图3 E)。
3 结论与讨论
小麦胚乳淀粉粒的粒径范围为0.4~48 μm,其数量分布呈单峰曲线变化,B型淀粉粒(粒径≤10 μm)占99%以上,A型淀粉粒(粒径>10 μm)不足1%;其体积分布呈双峰曲线变化,A型淀粉粒占40.4%~61.3%,B型淀粉粒占38.7%~59.6%;其表面积分布呈双峰或3峰曲线变化,B型淀粉粒的表面积(占82.0%~90.3%)显著高于A型淀粉粒(占9.7%~18.0%),3峰曲线中的B型淀粉粒的表面积又可分为B1型(峰值粒径为1.2 μm)占表面积的46.9%~47.8%,B2型(峰值粒径为4.9 μm)占38.2%~39.5%。
Waxy蛋白亚基的完全缺失对小麦淀粉粒的分布特征影响较大。表现为峰值粒径变大,1~10 μm淀粉粒数量增加,<1 μm淀粉粒数量减少;B型淀粉粒体积和表面积增加,A型淀粉粒的体积和表面积降低。Waxy蛋白亚基的部分缺失对小麦淀粉粒的分布也存在一定影响。相关性分析表明,直链淀粉的质量分数与B型淀粉粒的体积、表面积百分数、1~10 μm淀粉粒数量百分数之间呈显著负相关,与A型淀粉粒的体积、表面积百分数、<1 μm淀粉粒数量百分数之间呈显著正相关。直链淀粉的质量分数与A型淀粉粒的数量百分数之间相关不显著。
前人研究结果表明,淀粉粒组成对食品及非食品加工业有重要影响[2,3],由此促使人们开展有关淀粉粒分布状况的研究。但是关于淀粉粒分布的研究结果存在差异,指出小麦胚乳淀粉粒大小呈双峰[2]或3峰[7~10]曲线变化。
本研究中所选材料的淀粉粒最大粒径高于前人的研究结果(最大35 μm)。这可能与试验所选材料的品种特性有关。从最大粒径的分布范围可以看出,Waxy蛋白亚基的缺失对其影响不显著。由此可以推测,直链淀粉不是形成淀粉粒所必需的。
本次研究中所用仪器的检测下限为0.4 μm,而粒径为0.4~1.0 μm的淀粉粒占淀粉粒数量的47.8%~70.6%。正是因为<1 μm淀粉粒的检出,使得本研究中A型淀粉粒的数量降幅较大。本次研究中A型或B型淀粉粒的质量百分数与前人的研究结果不同,A型的质量百分比偏低,B型的偏高。A型和B型淀粉粒体积(或质量)的变化主要是由于淀粉粒数目的变化而引起的,与测定过程中所选用的光学模型和计算方法也有关。
直链淀粉含量和淀粉粒度分布各参数的相关性分析表明B型淀粉粒的数量增加时,直链淀粉含量有降低的趋势。可推测B型淀粉粒的直链淀粉含量较A型淀粉粒低;就B型淀粉粒内部而言,1~10 μm淀粉粒的直链淀粉的质量分数较<1 μm淀粉粒的低。不同粒级淀粉中直链淀粉的质量分数的高低,有待于将其分离提纯后测定证实。
[1] Rahman S,Kosar-Hashemi B,Samuel M S,et al.The major proteins of wheat endosperm starch granules[J].Aust J of Plant Physiol,1995,22:793-803.
[2] Peng M S,Gao M,Abdel-Aal E S M,et al.Separation and characterization of A- and B-type starch granules in wheat endosperm[J].Cereal Chem,1999,76(3):375-379.
[3] Morrison W RGadan H.The amylose and lipid contents of starch granules in developing wheat endosperm[J].J Cereal Sci,1987,5:263-275.
[4] Gaines C S,Raeker M,Tilley M,et al.Associations of starch gel hardness, granule size, waxy allelic expression, thermal pasting, milling quality, and kernel texture of 12 soft wheat cultivars [J].Cereal Chem,2000,77(2):163-168.
[5] Sahlstr m S,Brathen E,Lea P,et al.Influence of starch granule size distribution on bread characteristics[J].J of Cereal Sci,1998,28(2):157-164.
[6] Stoddard F L.Survey of starch particle-size distribution in wheat and related species[J].Cereal Chem,1999,76(1):145-149.
[7] Bechteal D B,Zayas I,Kaleikau L,et al.Size-distribution of wheat starch granules during endosperm development[J].Cereal Chem,1990,67(1):59-63.
[8] Bechtel D,Zayas I,Dempster R,et al.Size-distribution of starch granules isolated from hard red winter and soft red winter wheat[J].Cereal Chem,1993,70(4):238-240.
[9] Raeker M,Gaines C S,Finney P L,et al.Granule size distribution and chemical composition of starches from 12 soft wheat cultivars[J].Cereal Chem,1998,75(5):721-728.
[10] Takada Y,Takeda C,Mizukami H,et al.Structures of large, medium and small starch granules of barley grain[J].Carbohydratte polymers,1999,38:109-114.
[11] Nakamura T,Yamamori M,Hirano H,et al.Production of waxy (amylose free) wheat[J].Mol Gen Genet,1995,248(3):253-259.
[12] 姚金保,R Sharma,C F Jenner,等.缺失不同Wx蛋白对普通小麦直链淀粉含量及淀粉特性的影响[J].麦类作物学报,2005,25(6):29-33.
[13] 陈东升,C Kiribuchi-Otobe,徐兆华,等.Waxy蛋白缺失对小麦淀粉特性和中国鲜面条品质的影响[J].中国农业科学,2005,38(5):865-873.
[14] 姚大年,孙辉,李保云,等.小麦品种Waxy蛋白亚基缺失类型若干淀粉性状研究[J].中国粮油学报,1999,14(1):6-9.
[15] 李继刚,梁荣奇,张义荣,等.糯性普通小麦的产生及其淀粉特性研究[J].麦类作物学报,2001,21(2):10-13.
[16] 梁荣奇,张义荣,唐朝晖,等.糯性普通小麦的籽粒成分和淀粉品质研究[J].中国粮油学报,2002,17(4):12-16.
[17] 穆培源,何中虎,徐兆华,等.CIMMYT普通小麦品系Waxy蛋白类型及淀粉糊化特性研究[J].作物学报, 2006,32(7):1 071-1 075.
[18] 翟红梅,田纪春.小麦Wx基因突变体的建立及其淀粉特性的研究[J].作物学报,2007,33(7):1 059-1 066.
[19] 李春燕,封超年,王亚雷,等.不同小麦品种籽粒淀粉理化特性的差异及其相关[J].作物学报,2007a,33(7):1 129-1 134.
[20] 李春燕,封超年,王亚雷,等.不同小麦品种支链淀粉链长分配及其与淀粉理化特性的关系[J].作物学报,2007b,33(8):1 240-1 245.
(责任编辑:朱宝昌)
Size Distribution of Starch Granules in the Endosperm of Wheat with Different Waxy Protein Subunits
ZHANG Min1, LIU Xi-wei1, ZHAO Cheng1, SONG Xiao-jun1,DAI Zhong-min2, CAI Rui-Guo1
(1 College of Life Science and Technology, Hebei Normal University of Science & Technology, Qinhuangdao,Hebei, 066600;2 Department of Biology, Dezhou University;China)
In this research work, we used eleven wheat cultivars, which carried different Waxy protein subunits, to sdudy the the size-distribution characteristics of starch granules and their relationship with amylose content. The results showed that diameters of starch granules in wheat endosperm ranged from 0.4 μm to 48 μm and the maximum diameter varied with wheat cultivars. Wheat endosperm had a unimodal curve in starch granule number distribution and its peak diameter was below 1μm. B-type granules (diameter<10 μm) made up to more than 99% of the totle number of starch granules in wheat endosperm, while A-type granules (diameter≥10 μm) only accounted for less than 1%. There was a bimodal curve in starch guanule volume distribution in wheat endosperm and the two peak diameters locateded in 4.0-5.9 μm and 21.7-23.8 μm, respectively. B-type granules contributed to 38.7%-59.6% of the totle volume of starch granules in wheat endosperm and A-type granules accounted for 40.4%-61.3%. Wheat endosperm had a bimodal or trimodal curve in starch guanule surface area distribution. In the bimodal surface-area curve, peak diameters locateded in 2.1-2.8 μm and 19.8-21.7 μm, respectively. In the trimodal surface-area curve, peak diameters were 1.2 μm, 4.9 μm and 19.8 μm. The surface-area percent of B-type granules (account for 82.0%-90.3%) was more larger than A-type granules,( account for 9.7%-18.0%) and it could be diveded into two parts in trimodal curve: B1-type granules(peak diameter was 1.2 μm ) contributed to 46.9%-47.8% of the totle surface area of starch granules in wheat endosperm and B2-type granules(peak diameter was 4.9 μm ) accounted for 38.2%-39.5%. The size distribution of starch granules in wheat endosperm was influenced significantly by deficiency of all the three waxy protein subunits. Compared with non-waxy wheat, waxy wheat had a much larger peak diameter in the number distribution curve of starch granules. The percent of starch granules (daimeter1-10 μm) of waxy wheat increased, but the percent of starch granules (diameter<1 μm) decreased. The volume and surface area percent of B-type granules of waxy wheat increased, on the contrary, they decreased in A-type granules. The partial deficiency of waxy protein also had some effect on the size distribution of wheat starch granules. The results of correlation anslysis indicated that amylose content of wheat starch was significantly and negatively correlated with the volume and surface area percent of B-type granules and the number percent of starch granules (daimeter from 1 to 10 μm). On the contrary, it was significantly and positively correlated with the percent of volume and surface area and of A-type granules and the number percent of granules (daimeter<1 μm). There was no significent correlation between amylose content and the number percent of A-type starch granules in wheat.
wheat (TriticumaestivumL.); waxy protein subunit; starch; size distribution
国家自然科学基金项目(项目编号:31201157);河北省自然科学基金项目(项目编号:C2015407061);河北省教育厅优秀青年基金项目(项目编号:Y2012032)。
2015-07-27; 修改稿收到日期: 2015-10-14
10. 3969 /J. ISSN. 1672-7983. 2015. 04. 010
S512.1;S311
A
1672-7983(2015)04-0047-08
张敏(1980-),女,实验师,硕士。主要研究方向:种子质量检验与作物逆境生理。
*通讯作者,男,博士,副教授。主要研究方向:作物高产优质栽培。E-mail: cairuiguo@126.com。
