不同因子驱动下通过不同途径发生的红树林斑块数量和面积变化量的计量方法
2015-03-11李春干代华兵
李春干, 代华兵
广西林业勘测设计院, 南宁 530011
不同因子驱动下通过不同途径发生的红树林斑块数量和面积变化量的计量方法
李春干*, 代华兵
广西林业勘测设计院, 南宁 530011
为深入阐明区域红树林空间演变机理,需对红树林斑块数量和面积在不同因子驱动下通过不同途径发生的变化量进行准确计量。提出了在GIS平台支持下的通过空间叠置分析方法进行斑块数量和面积变化量计量的两种方法——精确计量法和整体计量法。首先将前后两期通过遥感图像提取的红树林斑块分布图、遥感图像进行叠合,采用视觉信息叠合方法,将全部斑块划分为众多具有相同主要驱动因子和变化途径的分析单元;若前后期遥感图像能够精确配准,采用精确计量法计量:通过线与多边形叠置方法,用前期斑块的线状图对后斑块的面状图、后期斑块的线状图对前斑块的面状图分别进行切割,每个分析单元得到多个亚斑块,逐一确定每个亚斑块的驱动因子、变化途径,据此统计每个分析单元中斑块数量和面积在不同因子驱动下通过不同途径发生的变化量;若前后期遥感图像难以精确配准,采用整体计量法计量:对于每个分析单元,根据斑块恢复的难易程度、面积和斑块数量变化量的大小,确定其主要驱动因子和主要变化途径,该分析单元前、后期斑块数量和面积之差即为其在监测期间由该因子驱动通过该途径发生的斑块数量和面积变化量。尽管整体计量法对红树林空间演变机制分析的结果与精确计量法存在一定差异,但也属于定量分析范畴,都能深刻阐明红树林空间演变机制,能够全面、准确地反映了区域红树林斑块数量和面积在监测期内增加、减少的动态过程。
红树林; 空间分布; 斑块数量; 面积; 斑块动态; 计量
红树林是重要的海洋生态系统,具有重要的社会经济和环境生态功能。1980—2005年,全球红树林减少了19%[1]。很多学者在红树林动态监测、红树林减少原因等方面做了大量的研究,基本上阐明了红树林减少的主要原因——人口增长压力造成的传统利用者过度开发、木材采伐等商业利用、滩涂用途转换(盐田、水产、农业、旅游、港口、码头等)、污染、自然灾害和管理失败等[1-24]。但是,现有分析方法大多是从整体的角度研究各种因素(驱动因子)对其面积变化(通常为减少)的影响和作用,所得结论通常是定性的,如“导致红树林面积减少的原因是农业、水产、城市扩展…”,很少得到定量的结论[1],即使进行定量分析,也只考虑人为因素影响造成的红树林面积减少量[3],并未全面阐明红树林在长期演替过程中,面积增加和减少的动态过程。已有研究提出了通过两期遥感图像提取红树林空间分布信息,在GIS支持下采用叠置分析方法,根据前、后两期各个斑块的空间位置、形状和面积变化情况以及图像表征,逐一分析确定每个斑块变化的主要驱动因子和变化途径,在此基础上构建斑块数量和面积变化的驱动因子-变化途径状态矩阵,定义了总驱动量、总驱动率、净驱动量、净驱动率、趋势驱动率、总流量、总流率、净流量、净流率、趋势净流率和作用力等系列指标概念及其计算方法,建立了红树林斑块空间演变机理分析体系,初步实现了红树林空间动态变化分析的定位化、定量化和精确化[25]。但关于红树林斑块数量和面积变化量计量方法的阐述过于简单,且存在着增加量与减少量相互抵消,导致结果不准确的缺陷,现将修正完善方法阐述如下。
1 红树林斑块变化情况分析及变化途径与驱动因子的分析确定
1.1 斑块的整体变化与局部变化
在一个不太短的时段内,受自然过程影响和各种人为活动干扰,红树林斑块的边界或多或少都会发生变化,不可能保持绝对稳定,可以说,红树林斑块边界的变化是绝对的,不变是相对的。如图1所示斑块,2001年和2007年的面积分别为93.4 hm2和86.4 hm2,边界长度分别为5472 m和6603 m,6年间,该斑块面积减少了7.5%,边界长度增加了20.7%。将两个年度的斑块进行叠合,可以发现除局部地段的边界没有发生变化(或变化量很小)外,大部分地段的边界都发生了变化(图1)。
由图1可以看出,与2001年相比,2007年该斑块大部分地段的边界在空间上呈收缩状态,致使该斑块边界在整体上呈萎缩状态。进一步分析还发现:一些地段边界呈收缩状态,另一些地段边界呈扩展(范围扩大)状态,并有一些地段边界保持不变,表明该斑块各地段的边界变化与整体变化不尽一致。作空间叠置分析后,两个年度的边界各截断为8段(图1),各线段长度的变化、空间位置变化情况及其引起斑块面积变化的情况详见表1。
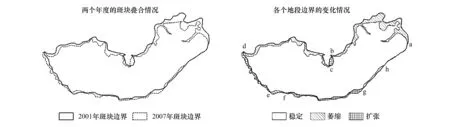
图1 通过叠置分析一个斑块的空间变化情况Fig.1 Overlay analyses on changes in the extent of a mangrove patch
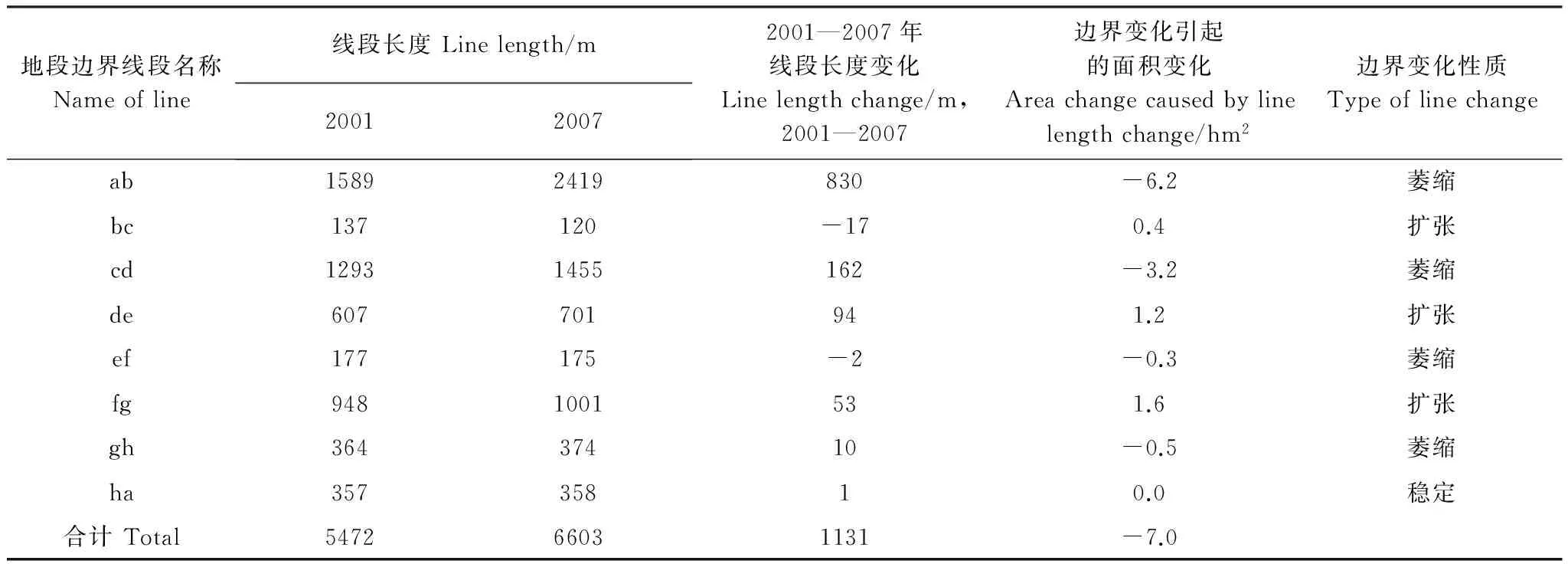
表1 斑块边界变化及其导致的面积变化情况Table 1 Changes in patch area caused by borderline changes
由表1可以看出,在8段边界中,只有一段边界(ha)保持稳定,长度为357 m,占总长度的6.5%,其余93.5%的边界都发生了变化,其中:有4段边界萎缩,长度为3423 m,占总长度的62.6%,由此减少的面积为10.2 hm2;有3段边界扩张,长度为1692 m,占30.9%,增加的面积为3.2 hm2。
对更多斑块的变化情况进行深入分析,结果表明:斑块整体变化情况与局部地段变化情况不完全一致的现象普遍存在,尤其是面积较大、边界较长的斑块,情况更是如此。其原因是斑块的各个地段遭受外界作用力的种类、影响程度不尽相同。如某些地段受养殖塘建设的破坏而萎缩,另一些地段因繁殖体扩展、生长成林而扩张。
整体由众多局部组成,局部地段变化之和构成了斑块整体的变化。由于局部变化不尽一致并且存在收缩与扩展等互补情况,因此,要全面、准确地阐明斑块的空间变化,不但需要分析斑块整体空间位置、形状和面积的变化情况,而且需要详细分析各个地段边界的变化情况及其导致的面积变化情况,在此基础上,深入分析斑块整体变化、局部变化发生的途径及其产生的原因。
1.2 斑块变化类型和变化途径
1.2.1 斑块整体变化类型与变化途径
斑块的整体变化是指在监测期内,斑块整体由前期的状态(空间范围、形状、面积大小)变成后期的状态的类型。根据监测期间斑块整体形状、边界空间位置和面积的变化情况,斑块整体空间分布的变化类型分为稳定斑块、扩张斑块、萎缩斑块、碎化斑块、消失斑块和新增斑块6种[25],不同变化类型的斑块状态变化特征见表2。

表2 不同变化类型的斑块空间整体变化特征Table 2 Overall spatial-temporal characteristics of patches with various dynamic procedures
斑块整体变化类型可在GIS平台支持下将两期红树林空间分布图叠合后,根据前、后期斑块的状态特征,按照上述定义通过目视解译方法确定。
以上斑块整体变化类型划分只是一般的、或某种程度上是典型的情况。在监测期内,尤其是当监测期较长时,受自然和人为因素影响,一个斑块可能出现不同类型的变化,即存在两个或多个变化类型的现象。如一些斑块中可能出现某些地段边界扩张,一些地段边界萎缩,还有一些地段边界碎化的情况。为便于统计分析,将斑块在监测期内变化的主要类型确定该斑块的整体变化类型,此时,一个斑块在监测期内只属于一个整体变化类型。主要变化类型根据如下条件确定:(1)斑块数量变化量的大小;(2)斑块恢复的难易程度;(3)面积变化量的大小。
如上所述,各个变化类型斑块都可能存在着面积变化的情况,即使斑块类型为稳定、扩张和萎缩时,斑块数量也可能存在少量变化,因此,可将斑块整体变化类型视为斑块整体变化的途径,即斑块整体变化途径等同于斑块整体变化类型。
1.2.2 斑块局部变化类型与变化途径
斑块局部地段变化是指斑块局部地段边界空间位置的变化,其将导致斑块数量和面积发生变化。
局部地段边界的变化由局部地段红树林群落个体增加或减少造成。一般情况下,一个地段红树林个体增加,必然导致该地段边界扩展;反之,必定出现该地段边界萎缩或碎化。从这个角度来看,对于局部地段而言,边界扩张与新增同义,边界萎缩、碎化与消失同义。然而,由于局部地段变化与斑块原有空间位置密切相关,扩展地段新增的红树林个体大多由原有红树林的繁殖体发展而来,并且与原有红树林群体紧密相连,萎缩或碎化地段的红树林个体虽然消失,但其紧邻区域尚有红树林个体存在。鉴于此,在文中,将新增的定义限于整体新增,新增的斑块与原有斑块在空间上不相连,消失限于整体消失。此时,斑块局部地段变化途径包括也稳定、扩张、萎缩、碎化、新增和消失6种[25]。
对两期斑块空间分布图作叠置分析后,只要斑块边界不完全重叠,总会得到很多小的亚斑块,为减少分析的工作量,规定只有某一亚斑块大于一定面积时(如0.1hm2),才认为其相应地段的边界发生了变化,否则,认为该地段的边界保持稳定。
1.3 斑块变化驱动因子的图像表征
斑块变化的驱动因子包括自然过程、养殖塘和盐田建设、围垦、工程建设、人工造林[25],各因子在遥感图像上具有不同的表征。
(1)自然过程
由于研究区内未出现过不以滩涂利用为目的的红树林连片砍伐活动,因此,若在后期的遥感图像上无人工设施(如海堤、港口码头、城市、道路、工业园区、养殖塘和盐田等)直接覆盖在前期红树林斑块上,或与斑块边界十分接近,则认为此斑块在监测期内的变化属自然过程引起的变化,或更准确地说是在无明显破坏性人为活动干扰下发生的变化。
(2)养殖塘和盐田建设
由于潮上带沿岸建造的高位池一般对红树林不构成直接影响,因此,养殖塘主要指低位池。低位池和盐田均建于潮间带的滩涂上。在开阔滩涂上,低位养殖塘的形状大多为矩形、近矩形或近梯形,形状规则,呈连片分布;在丘陵区海汊上,养殖塘多呈椭圆形或半圆形。每个养殖塘面积一般为0.5—5 hm2,池水深度为1.5—1.8 m,养殖塘之间或其边缘有完善的进、排水系统。由于蓄水,养殖塘在图像的灰度值接近滩涂外侧海水,与其直接相邻的滩涂存在较为明显的区别。盐田通常建在平坦的滩涂上,由多个蒸发池和结晶池组成,多为矩形、近矩形,形状十分规整且面积相等或接近,建有完善的进、排水系统。在遥感图像上,蒸发池的光谱特征因水位较深而与海水较为接近,灰度值较小;结晶池因水位较浅,灰度值较大,在图像上呈浅色;养殖塘与盐田主要区别为盐田内分布数量较多、面积小、形状十分规整的结晶池。
(3)围垦
围垦多在浅滩上进行,面积较大,向海一侧有较宽的海堤,其内分隔成形状较为规整、大小接近的水田或旱地。由于围垦后土地无水体覆盖,因此与周围的海面相比,图像的灰度值较大,表现为颜色较浅而与深色的海面形成较大的反差。
(4)工程建设
公路和铁路为线状地物,且路面反射率极高,在图像上呈浅白色,极易辨识。港口、码头因有高反射率硬化地面(货场)存在,并且图形较为规整,在图像上呈较大面积均质的浅色区域,与深色的海面具有明显的区别,在SPOT5、ALOS图像上甚至可见货物(如集装箱)堆积和船舶,工业园区的厂房也清晰可见,也见城市和工业厂区中规则分布的街道或道路。
(5)人工造林
人工造林多见于河口、内湾平缓的泥质滩涂。在空间分辨率极高的低空遥控无人机航空图像上,很容易将规则成行分布的红树林人工林与随机分布的天然群落分辨出来。在空间分辨率为2.5 m的ALOS、SPOT5图像上,难以通过图像分析区分人工林和天然林,因此,需对研究区进行全面的考察,深入访问当地红树林主管部门和沿海居民,详细了解造林的地点、范围、面积和造林年度,并尽可能取得造林设计或验收图,然后在此基础上,结合图像分析将人工林分布范围准确地标绘在红树林空间分布图上。
与斑块变化类型相似,斑块变化的驱动因子也分整体驱动因子和局部驱动因子。一个斑块在长期演变过程中,通常先后或同时受到多个因素影响,产生多种类型的变化。与斑块变化类型相似,将影响程度最大的因素,造成斑块整体变化的因素称为斑块变化的整体驱动因子,而其他一些因素,只在局部地段对斑块变化产生影响,造成局部地段边界发生变化,这些因素称为局部驱动因子。
2 分析单元划分及斑块数量和面积变化量的计量
2.1 斑块分析单元的划分
由于区域红树林斑块数量和面积存在着一些地段增加,另一些地段减少,从而相互抵消的现象,因此,为准确计量和分析监测期内红树林斑块数量和面积在不同因素驱动下通过各个变化途径发生的变化量,需将前、后期红树林空间分布图作叠置分析,逐一计算斑块数量和面积的变化量,分析确定其驱动因子和变化途径的种类。由于监测期内斑块存在各种类型的空间变化,期初斑块的空间范围与期末的空间范围不完全重叠。在作监测期内斑块数量和面积变化量计量、分析确定其驱动因子和变化途径时,将前、后两期红树林空间分布图叠合后,以期初斑块为基础,将研究区划分为众多包含1个或多个受相同因子驱动、整体变化类型相同的相邻斑块组成小区域。这些小区域称为斑块分析单元,是计算斑块数量和面积变化量、进行驱动因子和变化途径定量分析的基本单元,其划分方法如下:
(1)对于整体稳定斑块,期初斑块和期末斑块叠合后所占的空间范围为一个分析单元(图2)。若相邻区域几个斑块均为稳定斑块,则这几个斑块所占的空间范围可划为一个分析单元。

图2 通过叠合划分分析单元Fig.2 Demarcation of analysis unit by overlay
(2)对于斑块数量不发生变化的整体扩张斑块,期初斑块和期末斑块叠合后所占的空间范围为一个分析单元,若期初几个斑块扩张后至期末变成了1个斑块,则前、后期几个斑块叠合后所占的空间范围为一个分析单元(图2)。若相邻区域几个斑块均为扩张斑块,且其驱动因子相同,则这几个斑块所占的空间范围可划为一个分析单元。
(3)对于整体萎缩斑块,期初斑块和期末斑块叠合后所占的空间范围为一个分析单元(图2),若相邻区域几个斑块均为萎缩斑块,且其驱动因子相同,则这几个斑块所占的空间范围可划为一个分析单元。
(4)对于整体碎化斑块,期初斑块及由其碎化而得到的期末斑块叠合后所占的空间范围为一个分析单元(图2)。若相邻区域几个斑块均为碎化斑块,且其驱动因子相同,则这几个斑块所占的空间范围可划为一个分析单元。
(5)对于消失斑块,以由相同驱动因素影响而导致消失的前期的1个或多个邻近斑块的空间范围为一个分析单元(图2)。
(6)对于新增斑块,以由相同驱动因素(人工造林或自然过程)作用而得到的后期1个或多个斑块的空间范围为一个分析单元(图2)。
分析单元划分在GIS环境中进行,首先是建立一个面图层——分析单元图层,其次是叠合前、后两的斑块分布图层、遥感图像,根据图像、图形表征分析确定各个斑块变化的驱动因子和变化类型,然后通过屏幕矢量化方法绘画出互不重叠的多边形区域,即分析单元,并对其统一编号(图3)。

图3 分析单元划分图Fig.3 Distribution of analysis unites in a bay
一个分析单元内所有斑块的整体的变化类型(途径)和驱动因子必须相同,并且相邻分布。由于不同监测期的斑块空间分布不同,斑块变化的驱动因子和整体变化类型也不同,因此,需要逐一监测期划分分析单元,如2001—2007年的分析单元范围与2007—2010年的分析单元范围一般不完全重叠。
2.2 斑块数量和面积的变化量的计量
由于斑块的变化存在整体变化和局部变化两种情况,因此,要准确地掌握斑块的变化情况,需要深入细致地分析斑块各个局部地段的变化情况,对每个斑块各个地段的变化量作精确的计量,在此基础上综合得到区域红树林空间分布变化的准确结果,这种变化量的计量方法称为精确计量法。后续讨论中将会看到,这种方法需要满足极为严格的空间配准条件,在空间配准不准确情况下,采用这种方法将会得到错误的结论,此时,可以从整体变化途径和整体驱动因子的角度,采用另一种量化方法对斑块数量和面积的变化量进行计量,这种方法称为整体计量法。
2.2.1 精确计量法
(1)将前、后两期斑块空间分布图叠合,将研究区划分为众多相互独立的分析单元。
(2)采用面转换线的方法,将监测期间前、后两期的斑块空间分布面图层分别转换为线图层。
(3)以后期的线图层对前期的面图层进行切割(叠置),若后期斑块的空间范围位于前期斑块的空间范围以内,则将前期斑块切割为若干个小斑块(亚斑块),由此可计算前期斑块萎缩的面积,或碎化后斑块的个数和各个亚斑块的面积。
(4)以前期的线图层与后期的面图层进行切割(叠置),若后期斑块的空间范围超出前期斑块的空间范围,可将后期斑块切割为若干个亚斑块,并可计算得到后期斑块扩张的范围和面积。
(5)若前期斑块存在而后期不存在,则斑块消失,面积变化(消失)量等于期初斑块面积,斑块个数变化量等于该分析单元期初的斑块数量;若某一局部区域前期无红树林斑块而后期出现斑块,则这个(些)斑块为新增斑块,面积新增量等于期末斑块面积之和,斑块数量新增量等于该区域后期斑块的数量。
(6)若分割后得到的某个亚斑块面积小于0.1 hm2,则将其与相邻且驱动因子、变化途径相同的亚斑块合并。
(7)对全部亚斑块重新计算面积,根据图像表征逐一分析确定其驱动因子、变化途径,记录在面图层的属性表中,并记录其所处的分析单元编号。
如图4所示,2001年1个面积为24.4 hm2的斑块,至2007年时变成了5个斑块,总面积为18.1 hm2。作空间叠置分析,辅以人工分割(因考虑变化途径和驱动因子的不同,亚斑块13、15、17、18由原同一亚斑块经人工分割而得)并对面积小于0.1 hm2的小斑块删除后,该分析单元分割为18个亚斑块。对斑块数量和面积通过各个变化途径发生的变化量进行分析,结果表明:海堤建设导致2001年的一个斑块碎化为3个部分,由此减少的面积为5.6 hm2,其中,1号亚斑块和2号亚斑块各构成一个部分,3号、4号、5号和6号亚斑块构成另一个部分,故认为海堤建设引起的斑块数量变化量为2个;3号、4号、5号和6号4个亚斑块所在的区域,从图像上未见工程或养殖塘建设等直接的人为活动影响,故认为该区域的斑块碎化为自然状态下发生的,即自然过程导致斑块的变化量为2个。4个地段的边界扩张;7个地段的边界出现萎缩;2个地段的边界碎化。将叠置结果与后期遥感图像叠合,通过目视解译方法,逐一判断每个亚斑块变化产生的原因,即得到该分析单元由各个驱动因素引起的斑块数量和面积变化量(图4)。对该分析单元中18个亚斑块的变化原因和变化途径进行逐一分析,得到表3的结果。
对表3进一步统计,得到该分析单元红树林斑块个数和面积在各个因素驱动下通过各个途径发生的变化量,见表4。
表4也说明了红树林斑块变化的复杂性:一是在同一因素驱动下,红树林斑块面积的变化可通过多个途径发生;二是在监测期内,斑块面积一方面通过扩张而增加,另一方面通过萎缩、碎化而减少,呈增加-减少的动态过程,如本例的斑块面积由2001年的26.0 hm2减少至2007年的17.0 hm2,净减少了9.0 hm2,但期间通过斑块扩张而使面积毛增加了0.4 hm2。
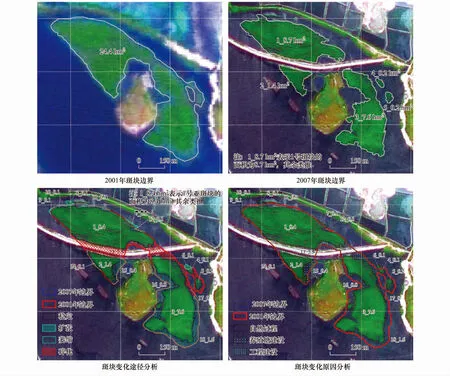
图4 精确计量法用于斑块数量和面积变化量计量Fig.4 Computing changes in ptatch count and area with the accurate measurement method
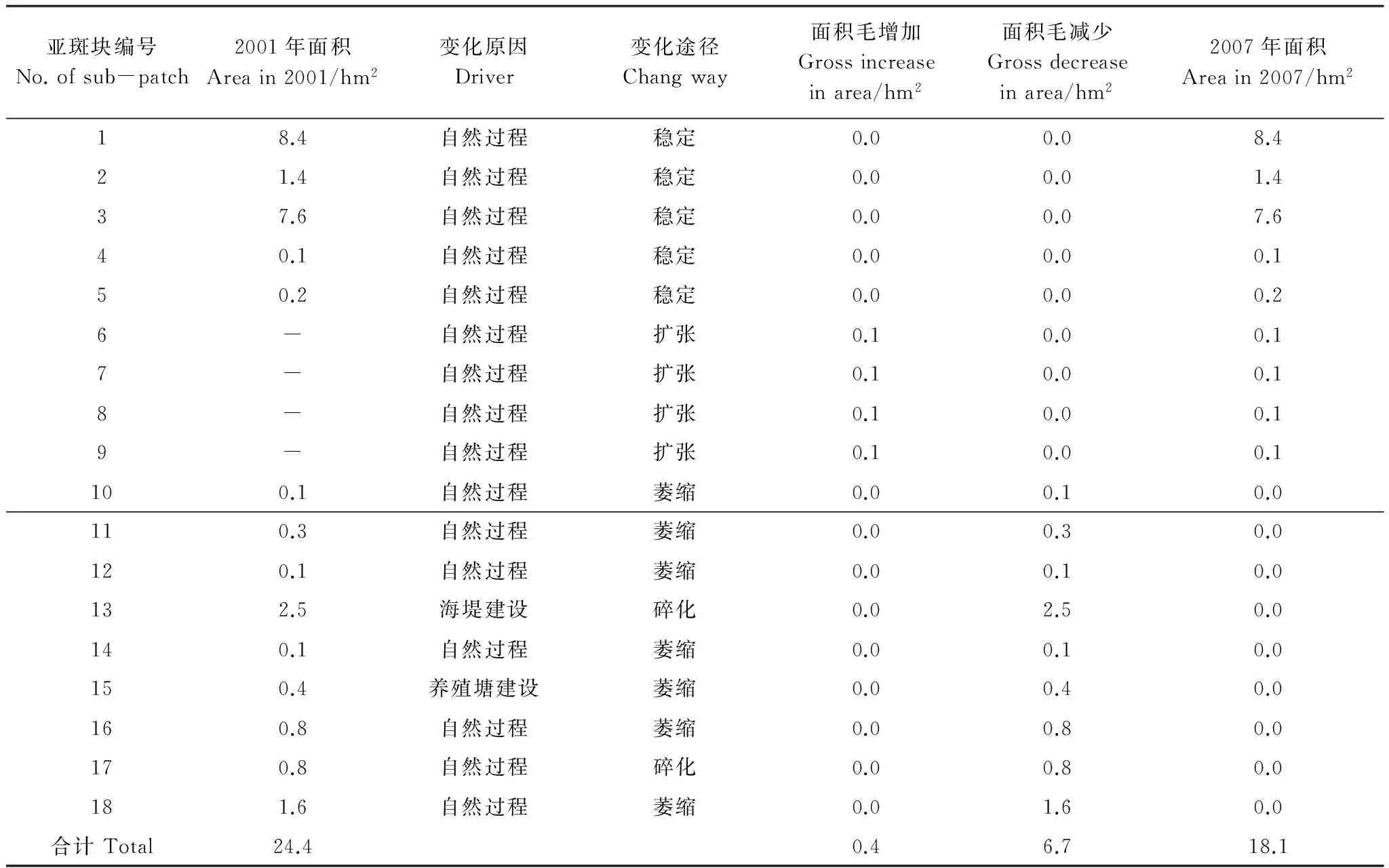
表3 各个亚斑块面积变化量详细表Table 3 Dynamic procedures and drivers of sub-patches
表4 用精确计量法计量的各个因素驱动下通过各个途径发生的斑块个数和面积变化量
Table 4 Changesin patch count and area through various dynamic procedures caused by various driversdetermined with the accurate measurement method

项目Item驱动因素Driver稳定Stable扩张Expand萎缩Shrink碎化Fragmentation消失Disappear新增New毛增加量Grossincrease毛减少量Grossdecrease总变化量Totalchange净变化量Netchange斑块数量/个自然过程0002002022Patchcount围垦0000000000养殖塘和盐田0000000000工程建设0002002022人工造林0000.0000000合计0004004044面积自然过程0.00.4-3.0-0.80.00.00.43.84.2-3.4Area/hm2围垦0.00.00.00.00.00.00.00.00.00.0养殖塘和盐田0.00.0-0.40.00.00.00.00.40.4-0.4工程建设0.00.00.0-2.50.00.00.02.52.5-2.5人工造林0.00.00.00.00.00.00.00.00.00.0合计0.00.4-3.4-3.30.00.00.46.77.1-6.3
2.2.2 整体计量法
精确计量法适用于前、后期遥感数据能够精确配准的研究案例。在通常的研究中,特别是时间跨度较大的研究,由于遥感数据种类多样、传感器多样,空间分辨率不同甚至相差很大,因此,难以保证每个监测期的基础数据都能精确配准。在这种情况下,若对面积变化量采用上述方法进行精确计量,将会得到错误的结果。如一个本来边界无任何变化,面积变化量为0.0 hm2的绝对稳定斑块,由于图像配准误差而出现了中心位置平移偏差了20.6 m。作叠置分析后,得到几个途径的变化量为:稳定10.9 hm2、萎缩1.0 hm2、扩张1.0 hm2。显然,这个结果是错误的。
中、高空间分辨率图像之间,虽然几何精校正都能够将误差控制在一个像元之内,但由于中分辨率图像提取的斑块边界精度低于高空间分辨率图像,因此,当采用精确计量法分析其斑块变化时,也将会得到错误的结果。对于历史的纸质红树林分布图,由于调查时都采用1∶10000—1∶50000地形图目测勾绘方法确定斑块边界,准确性较差,另一方面,由于制图技术和手段限制,图面几何精度不高,加以年代久远,纸张变形,要做到与遥感图像精确配准,十分困难。因此,在时间跨度大的多期红树林空间分布动态监测中,对于其中一些基础数据不能精确配准(如扫描航空图像),或前后期图像空间分辨率相差较大的监测期,即使采用精确计量法对斑块数量和面积变化进行计量,也难以取得正确的分析结果。
为解决上述问题并做到红树林空间分布动态分析的定量化,采用下述整体计量法对斑块面积和斑块数量变化量进行计量。
整体计量法的基本思想:对于每个分析单元,无论其在监测期内受到多少个驱动因素影响,通过多少个途径发生变化,一律只考虑一个影响程度最大的主要驱动因子和一个变化量最大的主要变化途径,监测期间该分析单元发生的斑块数量和面积变化量都记为该因子驱动、通过该途径发生的变化量。当一个斑块同时受多个因子驱动、存在多种变化途径时,从因果关系的角度出发,首先分析确定其主要驱动因子,然后在该主要驱动因子造成的多个变化途径中确定主要变化途径。主要驱动因子和主要变化途径的确定,综合考虑3个因素:①斑块恢复的难易程度;②面积变化量的大小;③斑块数量变化量的大小。
如图4,虽然工程(海堤)建设、养殖塘建设和自然过程都是导致斑块变化的原因,但由于海堤是永久性构筑物,一旦建成,不会轻易毁坏,因此,海堤建设是导致斑块发生变化的主要原因。在海堤建设影响下,斑块通过碎化、萎缩两个途径发生变化,但碎化造成的面积变化量最大,因此,该斑块主要变化途径为碎化。也就是说,在海堤建设的影响下,通过碎化途径,斑块数量增加了4个,面积减少了6.3 hm2。据此,得到斑块数量和面积变化量的计量结果如表5。
比较表4和表5,除每个分析单元只考虑1个驱动因子、1个变化途径外,整体计量法只考虑、计算斑块数量和面积的净变化量,而在精确计量法中能够同时考虑多个驱动因子、多个变化途径,并且能够计算监测期间斑块数量和面积的增加量和减少量。故整体计量法不能真正全面、准确地反映监测期内斑块数量和面积增加-减少的动态过程。尽管如此,该方法也逐一分析单元分析了斑块数量和面积变化的主要驱动因子和变化发生的主要途径,并计算了斑块数量和面积变化量,在一定程度上能够较为准确地反映了斑块数量和面积增加-减少的动态过程,与传统只考虑区域红树林面积净变化量的分析方法相比,其结果的科学性更强,也更具实际应用价值。
表5 用整体计量方法计量的各个因素驱动下通过各个途径发生的斑块个数和面积变化量
Table 5 Changesin patch count and area through various dynamic procedures caused by various drivers determined with the holistic measurement method
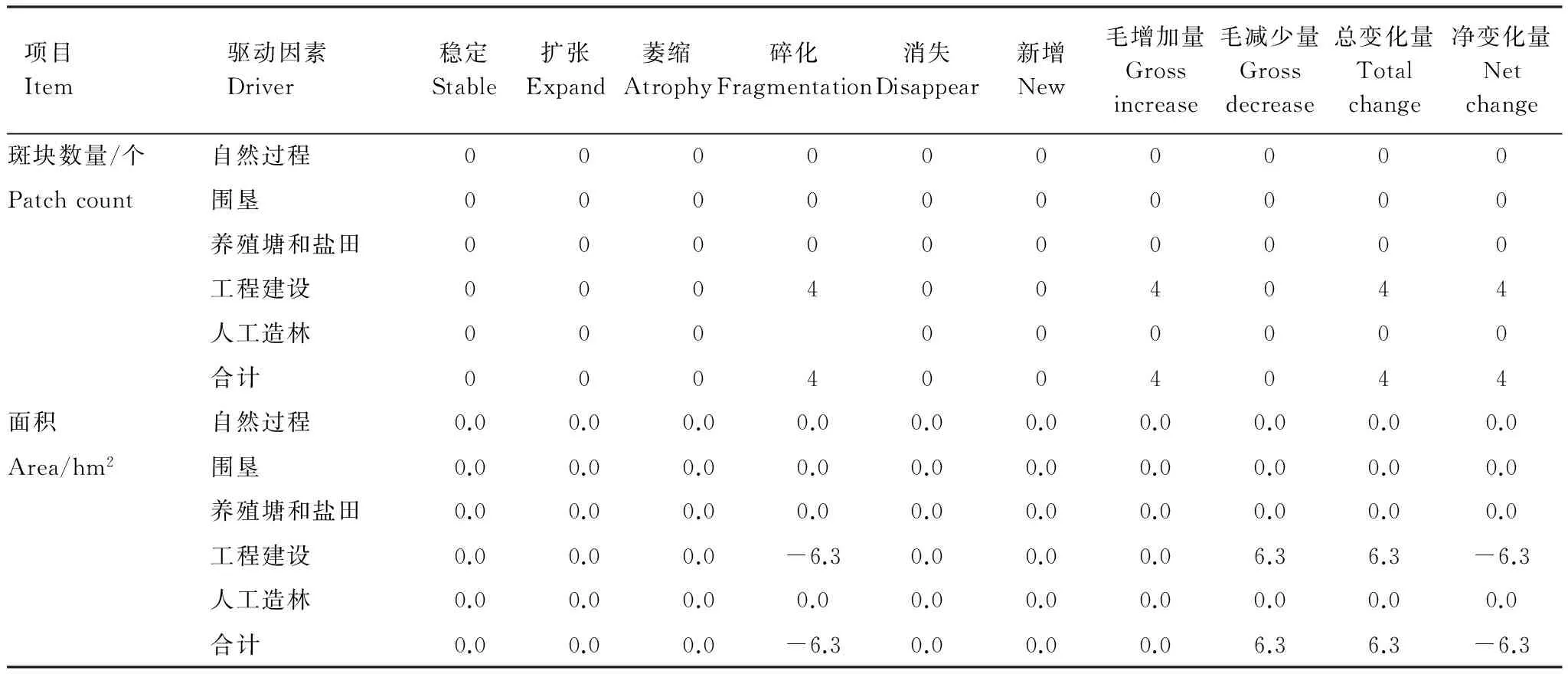
项目Item驱动因素Driver稳定Stable扩张Expand萎缩Atrophy碎化Fragmentation消失Disappear新增New毛增加量Grossincrease毛减少量Grossdecrease总变化量Totalchange净变化量Netchange斑块数量/个自然过程0000000000Patchcount围垦0000000000养殖塘和盐田0000000000工程建设0004004044人工造林000000000合计0004004044面积自然过程0.00.00.00.00.00.00.00.00.00.0Area/hm2围垦0.00.00.00.00.00.00.00.00.00.0养殖塘和盐田0.00.00.00.00.00.00.00.00.00.0工程建设0.00.00.0-6.30.00.00.06.36.3-6.3人工造林0.00.00.00.00.00.00.00.00.00.0合计0.00.00.0-6.30.00.00.06.36.3-6.3
3 两种计量方法对红树林空间演变机理分析结果的比较分析
为比较分析上述两种计量方法对区域红树林空间演变机理分析结果的差异,以防城港湾(包含东湾和西湾)为试验区采用文献[25]中提出的方法进行红树林空间演变机理分析。
3.1 研究区概况和数据源
防城港湾位于广西北部湾的西部,包含沙田墩以西、江山半岛白龙以东的海域,防城港城市和港口将该海湾分为东、西两个部分。2010年有红树林面积672.3 hm2,占广西红树林面积的9.7%,其中有我国面积最大的城市红树林(防城港渔洲坪一带)。
卫星遥感数据为2007年10月28日成像的SPOT5 HRG和2010年9月19日成像的ALOS PRISM/AVNIR-2图像,空间分辨率均为:全色波段2.5 m,多光谱波段10 m。用于几何精校正的参考数据为2004年真彩色正射航空图像(1∶10 000)。
3.2 信息提取与比较方法
3.2.1 图像预处理及红树林空间分布信息提取
为确保前、后期遥感图像精确配准,卫星图像几何精校正的总体均方根误差(RMS)都控制在1个像元以内。红树林斑块信息提取采用多方法联合应用的方法进行:①将研究区裁剪为众多仅含红树林和少量滩涂的小区域,个别情况下,一个小区域仅包含一个红树林斑块;②联合应用波谱运算、监督分类、无监督分类和面向对象分类以及图像分析方法提取斑块空间分布信息;③在GIS平台上对分类结果图采用屏幕矢量化方法提取红树林斑块边界,并确保前后期斑块边界变化符合空间逻辑。
3.2.2 斑块数量和面积变化量计量
研究区内2007年和2010年红树林斑块数量分别为154个和157个。根据斑块变化的驱动因素和变化途径,将研究区划分为157个分析单元。斑块数量和面积变化量的计量分别采用精确计量法和整体计量法两种方法进行。
3.3 结果分析
2007年研究区红树林斑块数量和面积分别为154个、705.6 hm2,至2010年斑块数量增加至157个,面积减少为672.3 hm2,3年间面积减少了33.3 hm2,年均减少11.1 hm2,年均变化率为1.6%。
3.3.1 变化途径比较分析
对研究区2007—2010年斑块数量变化情况作流量分析,结果表明:两种方法的得到结果相同(表6)。

表6 斑块数量和面积通过各个变化途径发生的流量(率)比较Table 6 Comparison of the amount (rate) of changes in patch count and area through various dynamic procedures
ST:稳定 Stable;EX:扩张Expand;AT:萎缩Atrophy;FR:碎化Fragmentation;DI:消失Disappear;NE:新增New;TC:总变化量Total change
由于监测期较短,斑块数量都只通过一个途径发生变化,故两种计量方法对斑块数量变化途径分析的结果相同。然而,对于面积变化,两种方法得到的结果相差较大:①面积变化的总流量相差较大,精确计量法的总流量几乎大于整体计量法总流量的一倍;②面积变化在各个途径上的流量(率)也存在较大差别,在精确计量法中,通过斑块萎缩和扩张途径发生的面积变化总流率分别为61.2%和30.8%,而在整体计量法中,两者的总流率分别为54.7%和11.6%,在斑块稳定的情况下,整体计量法的面积变化总流率达到了17.5%,而在精确计量法中,总流率为0;③在各个途径上发生的面积净变化流量(率)也不相同,在精确计量法中,斑块萎缩是面积减少的最主要途径(趋势流率为-90.0%),其余是消失(-5.7%)和碎化(-4.3%),而在整体计量法中,虽然斑块萎缩(趋势流率为-70.9%)也是面积变化的最主要途径,但其净流率明显减小,稳定变成了面积减少的一个重要途径(-12.8%)。
3.3.2 驱动力比较分析
与变化途径分析结果一样,对于斑块数量变化的驱动量(率),两种方法的结果相同(表7)。但对于面积变化,两种方法得到的结果不甚相同,各个驱动因子的驱动量(率)也存在较大差别。在精确计量法中,在自然过程驱动下面积的总变化量最大(总驱动率为53.8%),其次是养殖塘和盐田建设(36.0%),而在整体计量法中,自然过程的总驱动率只有38.5%,养殖塘和盐田建设的总驱动率达到了50.9%,几乎是两个驱动因子相互调换了次序。
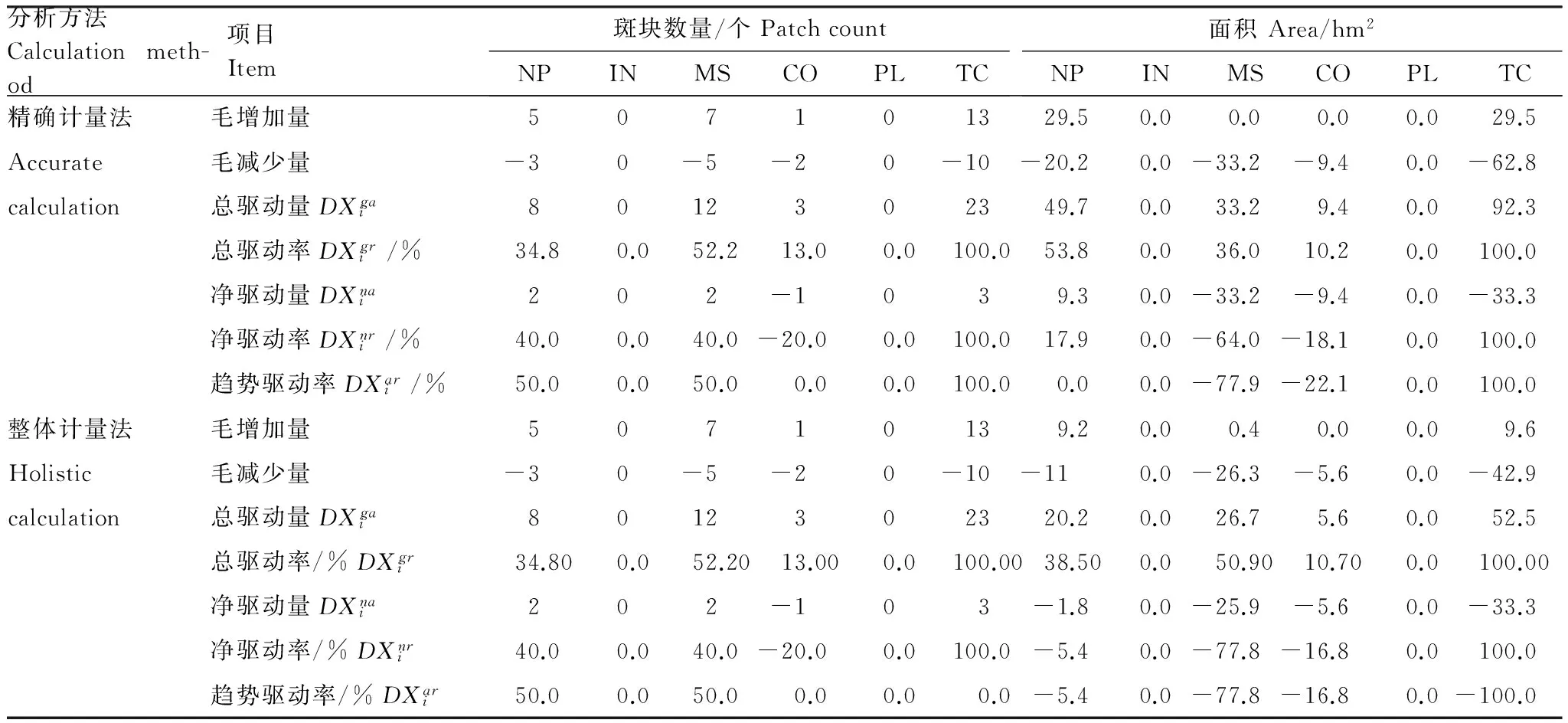
表7 斑块数量和面积变化的驱动力(率)比较Table 7 Comparison of the amount (rate) of changes in patch count and area caused by various drivers
NP:自然过程Natural process;IN:围垦Inning;MS:养殖塘建设Marineculture and saltern;CO:工程建设Construction;PL:人工造林Plantation
3.3.3 作用力比较分析
对于斑块数量的变化,两种方法得到的净作用力分析结果完全一致,但面积变化的净作用力分析结果差异较大(表8)。在对面积变化作精确计量时,斑块面积的净变化主要由养殖塘和盐田建设影响下通过斑块萎缩、自然过程影响下通过斑块扩张和萎缩产生,工程建设影响下通过斑块萎缩也在一定程度上造成面积变化。在采用整体计量法分析时,养殖塘和盐田影响下的斑块萎缩仍然是面积变化的最重要的驱动因子和变化途径组合,并且其净作用力更大,自然过程下的斑块萎缩对面积净变化影响也较大,自然过程影响下的斑块稳定、自然过程影响下的斑块扩张对面积净变化的影响程度也有所减弱。
3.4 两种方法分析结果存在差异的原因分析
以上分析结果可知,两种变化量计量方法对面积变化的分析结果存在较大差异,其根本原因是两种方法对面积变化量的计量方法存在根本的差别。在采用整体计量法对监测期内的面积变化量进行计量时,一个分析单元只考虑一个驱动因子、一个变化途径,并且只考虑一个净变化量(后期面积与前期面积之差),该变化量为在某个因子驱动下通过某个途径发生的面积变化量。而在采用精确计量法时,同时考虑多个变化量、多个驱动因子和多个变化途径。因此,即使研究区只有一个分析单元,其结果是存在较大的差别。

表8 净作用力比较Table 8 Comparison of acting forces with regard to changes in patch count and area
4 讨论和结论
(1)以往研究[25],考虑监测期内每个斑块都受到某个因子驱动、通过某种途径发生了变化,并由此分别得到了全部斑块个数和面积在期初、期末的驱动因子-变化途径状态矩阵,将期末状态矩阵减期初状态矩阵得到了监测期间斑块数量和面积的变化矩阵,并由该矩阵计算总驱动量、总驱动率、净驱动量、净驱动率、趋势驱动率、总流量、总流率、净流量、净流率、趋势净流率和作用力系列指标,用以阐明区域红树林空间分布的演变机制。该文在斑块数量和面积变化量计量中存在着如下缺陷:只从区域的角度计算斑块数量和面积在不同因子驱动下通过每个途径发生的净变化量,由于斑块数量和面积在某一因子驱动下通过某个途径存在着增加和减少两种变化情况,必须同时考虑增加量、减少量、总变化量和净变化量,因此,仅通过净变化量不能全面、准确反映在该因子驱动下通过该途径发生的斑块数量和面积的变化情况。本文通过将研究区划分为众多具有相同驱动因子、相同变化途径的分析单元,逐一分析单元计算斑块数量和面积的毛增加量、毛减少量和总变化量、净变化量,避免了上述问题,能更全面、准确地反映了区域红树林斑块数量和面积在监测期内增加、减少的动态过程。
(2)精确计量法准确地计量了不同驱动因子和变化途径组合下斑块数量和面积的变化量,结果最为可靠。虽然整体计量法对每一个分析单元只考虑一个主要驱动因子、一个主要变化途径,并且只计量斑块数量和面积的净变化量,但是,由于区域由众多分析单元构成,不同分析单元的斑块数量和面积净变化量有增有减,在作区域统计汇总后,可得到某一驱动因子某个变化途径组合的斑块数量和面积的毛增加量、毛减少量、总变化量、净变化量,在一定程度上也能较为准确地反映了区域斑块数量和面积增加-减少的动态过程,以往研究只考虑某个变化途径组合的斑块数量和面积的净变化量[25]相比,其结果的科学性更强,也更具实际应用价值。
(3)在区域红树林空间动态演变分析中,采用文中提出的方法对斑块数量和面积在不同因子驱动下通过不同途径发生的变化量进行计量,通过红树林空间动态定量分析体系[25],可以厘清如下问题:①监测期内斑块数量和面积的变化量(毛增加量、毛减少量、总变化量、净变化量)是多少?②何种因素的影响导致红树林斑块数量和面积发生变化?在任一既定因素驱动下,斑块数量和面积的变化量是多少?何种因素是斑块数量和面积增加(或减少)的主要驱动因子?③斑块数量和面积是通过何种途径发生变化的?通过每个途径发生的变化量是多少?何种途径是斑块数量和面积增加(或减少)的主要途径?④在任一既定的因素驱动下,斑块数量和面积通过任一既定变化途径发生的变化量是多少?何种驱动因素-变化途径组合对斑块数量和面积增加(或减少)的影响程度最大?
[1] FAO (Food and Agriculture Organization of the United Nations). The world′s mangroves 1980—2005//FAO Forestry Paper 153. Rome: FAO, 2007.
[2] Macintosh D J, Ashton EC. A review of mangrove biodiversity conservation and management. 2002. [2010-10-20]. http://mit.biology.au.dk/cenTER/MCB_Files/2002_Review_WB_MCB_Final.pdf
[3] Giri C, Ochieng E, Tieszen L L, Zhu Z, Singh A, Loveland T, Masek J, Duke N. Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecology and Biogeography,2011, 20(1):154-159.
[4] Groombridge B. Global biodiversity: status of the earth′s living resources//UNEPWCMC/The National History Museum/IUCN/Worldwide Fund for Nature/World Resources Institute. London: Chapman and Hall, 1992.
[5] Spalding M D, Blasco F, Field C D. World mangrove atlas.Okinawa: The International Society for Mangrove Ecosystems, 1997.
[6] Aizpuru M, Achard F, Blasco F. Global assessment of cover change of the mangrove forests using satellite imagery at medium to high resolution. EEC Research Project No. 15017-1999-05 FIED ISP FR. Ispra: Joint Research Centre, 2000.
[7] Giri C, Zhu Z, Tieszen L L, Singh A, Gillette S, Kelmelis J A. Mangrove forest distributions and dynamics (1975-2005) of the tsunami-affected region of Asia. Journal of Biogeography, 2008, 35(3): 519-528
[8] Ramsey E W Ⅲ, Jansen J R. Remote sensing of mangrove wetlands: relating canopy spectra to site-specific data. Photogrammetric Engineering and Remote Sensing, 1996, 62(8): 939-948.
[9] Selvam V, Ravichandran K K, Gnanappazham L, Navamuniyammal M. Assessment of community-based restoration of Pichavaram mangrove wetland using remote sensing data. Current Science, 2003, 85(6):794-798.
[10] Ramasubramanian R, Gnanappazham L, Ravishankar T, Navamuniyammal M. Mangroves of Godavari-analysis through remote sensing approach. Wetlands Ecology and Management, 2006, 14(1): 29-37.
[11] Vadlapudi S. Identification and quantification of changes in mangrove forest using remote sensing-a case study near Kakinada Bay, Andhra Pradesh, Noida, India.GIS Development Pvt. Ltd. 2003. [2011-5-20]. http://www.GISdevelopment.net/application/environment/wetland/mi03075.htm.
[12] Krause G, Bock M, Weiers S, Braun G. Mapping land-cover and mangrove structures with remote sensing techniques: A contribution to a synoptic GIS in support of coastal management in North Brazil. Environmental Management, 2004, 34(3): 429-440.
[13] Aksornkoae S. Ecology and Management of Mangroves. Switzerland: IUCN, Wetlands and Water Resources Programme, 1993.
[14] Giri C, Delsol J P. Mangrove forest cover mapping using remote sensing data in conjunction with GIS. Asian and Pacific Remote Sensing Journal, 1995, 8: 13-26.
[15] Blasco F, Aizpuru M, Gers C. Depletion of the mangroves of Continental Asia. Wetlands Ecology and Management, 2001, 9(3): 255-266.
[16] Verheyden A, Dahdouh-Guebas F, Thomaes K, de Genst W, Hettiarachchi S, Koedam N. High-resolution vegetation data for mangrove research as obtained from aerial photography. Environment, Development and Sustainability, 2002, 4(2): 113-133.
[17] Fromard F, Vega C, Proisy C. Half a century of dynamic coastal change affecting mangrove shorelines of French Guiana. A case study based on remote sensing data analyses and field surveys. Marine Geology, 2004, 208(2/4): 265-280.
[18] 范航清, 黎广钊. 海堤对广西沿海红树林的数量、群落特征和恢复的影响. 应用生态学报, 1997, 8(3): 240-244.
[19] 陈树培, 梁志贤, 邓义. 中国南海海岸的红树林. 广西植物, 1988, 8(3): 215-224.
[20] 陈远生, 甘先华, 吴中享, 钟德群. 广东省沿海红树林现状和发展. 广东林业科技, 2001, 17(1): 20-26.
[21] 张乔民, 隋淑珍. 中国红树林湿地资源及其保护. 自然资源学报, 2001, 16(1): 28-36.
[22] 黎夏, 刘凯, 王树功. 珠江口红树林湿地演变的遥感分析. 地理学报, 2006, 61(1): 26-34.
[23] 曹林, 韩维栋, 李凤凤, 张裕娇. 雷州湾红树湿地景观格局演变及驱动力分析. 林业科技开发, 2010, 24(4): 18-23.
[24] 王胤, 左平, 黄仲琪, 邹欣庆. 海南东寨港红树林湿地面积变化及其驱动力分析. 四川环境, 2006, 25(3): 44-49.
[25] 李春干, 刘素青, 范航清, 代华兵. 基于斑块的红树林空间演变机理分析方法. 生态学报, 2012, 32(14): 4329-4342.
Two methods for measuring the changes in patch-number and area of mangrove driven by various factors through different ways
LI Chungan*, DAI Huabing
GuangxiForestInventoryandPlanningInstitute,Nanning530011,China
In order to investigate the mechanisms of mangrove′s spatial dynamics, changes in landscape patch count and area caused by various drivers through different dynamic procedures need to be accurately quantified. Based on previous work, two calculation methods, namely accurate measurement method and holistic measurement method, are introduced in this paper. Firstly, two time-period distribution maps of mangrove and remote sensing images were overlaid with geographic information systems (GIS) and all mangrove patches were divided into analysis units, in which each patch was caused by the same driver and changed through the same procedure. Secondly, if the time-1 image could be registered accurately with time-2 image, we used the accurate measurement method. With the overlay methods of lines and polygons, the time-1 polygon layer of mangrove patches was subset by the time-2 line layer of mangrove patches, and the time-2 polygons were subset by the time-1 lines, resulting in several sub-patches in an analysis unit. Each sub-patch was visually interpreted to identify the driver and the dynamic procedure. Subsequently, the amount of changes with regard to the patch count and area of each analysis unit caused by various driving factors through different change ways were calculated. Thirdly, if the earlier image cannot be registered accurately with later image, we used the holistic measurement method. Each analysis unit was visually interpreted to identify the key driver and the major dynamic procedure according to both the level of difficulty in patch recovery and the amount of changes in patch count and area. Accordingly, the differences in patch count and area between the two periods in the analysis unit were regarded as the amount of changes in patch count and area caused by this driver and through this procedure. Both methods could be used to conduct quantitative analysis for the changes in patch count and area. The accurate measurement method is more reliable because it computes changes in patch count and area under various combinations of different drivers and dynamic procedures. Although the holistic measurement method only considers the major driver and dynamic procedure within an analysis unit, its statistics include the gross gain, gross loss, the total changes and net changes in patch count and area within an analysis unit, which also have scientific values and practical significance.
mangrove; distribution; patch count; area; patch dynamics; measurement
国家自然科学基金(41166001); 中央财政林业科技推广示范资金(GXTG200904); UNEP-GEF-SCS-防城港红树林国际示范区资助项目
2013-05-30;
日期:2014-07-18
10.5846/stxb201305301247
*通讯作者Corresponding author.E-mail: gxali@126.com
李春干, 代华兵.不同因子驱动下通过不同途径发生的红树林斑块数量和面积变化量的计量方法.生态学报,2015,35(6):1713-1726.
Li C G, D B.Two methods for measuring the changes in patch-number and area of mangrove driven by various factors through different ways.Acta Ecologica Sinica,2015,35(6):1713-1726.
