武夷山典型常绿阔叶林物种组成与群落结构
2015-03-10杨云方徐海根方炎明伊贤贵温小荣徐鲜均
丁 晖,杨云方,徐海根,方炎明,陈 晓,杨 青,伊贤贵,徐 辉,温小荣,徐鲜均
1 环境保护部南京环境科学研究所,国家环境保护生物安全重点实验室,南京 210042 2 南京信息工程大学遥感学院,南京 210044 3 南京林业大学森林资源与环境学院,南京 210037 4 福建省武夷山生物研究所,武夷山 354300
武夷山典型常绿阔叶林物种组成与群落结构
丁 晖1, *,杨云方1,2,徐海根1,方炎明3,陈 晓4,杨 青4,伊贤贵3,徐 辉4,温小荣3,徐鲜均4
1 环境保护部南京环境科学研究所,国家环境保护生物安全重点实验室,南京 210042 2 南京信息工程大学遥感学院,南京 210044 3 南京林业大学森林资源与环境学院,南京 210037 4 福建省武夷山生物研究所,武夷山 354300
以福建武夷山典型常绿阔叶林1.44 hm2固定样地调查数据为基础,从群落物种组成、区系特征、径级结构和空间分布格局等4个方面分析该地区常绿阔叶林物种组成和群落结构,结果表明:样地内共有胸径≥1 cm的木本植物139种9,116株,隶属于34科65属;样地内有稀有种23种,偶见种46种,优势树种不明显。区系特征明显,科级与属级地理成分以温带性质较强。群落及重要值>3的12个植物种群径级结构呈倒“J”字型。从物种的空间分布格局来看,这12个植物种群和4个主要种群的大、中、小径木都表现出明显的集聚分布,但集聚度有所差异。大部分物种随着径级的增加集聚度变小。
典型常绿阔叶林;物种组成;径级结构;空间分布格局;武夷山
生物多样性是人类赖以生存和发展的物质基础。我国是世界上生物多样性最丰富的国家之一[1]。但是,在过去的几百年中人类使物种灭绝速率比地球历史上物种自然灭绝速率增加了1000倍[2- 3]。生物多样性正面临栖息地被破坏、过度利用、环境污染、外来物种入侵、气候变化等诸多威胁[4]。生物多样性保护已经成为国际社会最为瞩目的重大问题之一。森林作为生物多样性保护最为重要的陆地生态系统,对其生物多样性的保护就显得尤为重要。以动态监测样地为主的森林生物多样性监测受到越来越多的关注,为人们了解物种组成与群落结构提供了详实的数据。以CTFS(the Centre for Tropical Forest Science)生物多样性监测网络最具代表性。我国在2004年到2012年也已建成中国森林生物多样性监测网络,其监测站点已达12个,如吉林长白山[5- 6]、北京东灵山[7]、河南宝天曼、广东鼎湖山[8]、西双版纳和广西弄岗等5—25 hm2大型动态监测样地。
亚热带常绿阔叶林是全球亚热带大陆东岸湿润气候和季风气候条件下的产物[9],广泛分布在南北纬22°—40°之间,以我国分布面积为最大,也是我国典型的森林植被类型之一[10]。武夷山是我国生物多样性的关键区域之一[11],该地区分布着大面积的亚热带常绿阔叶林。近年来,关于亚热带常绿阔叶林的物种组成与群落结构方面的研究已多有报道[12- 15],但对武夷山地区常绿阔叶林植物种群相关报道仍然较少,还有待进一步研究。本文通过对武夷山典型常绿阔叶林1.44 hm2永久性森林动态样地调查,就物种组成、区系特征、径级结构和空间分布格局进行了分析和总结,以期为进一步认识武夷山常绿阔叶林生物多样性和物种共存机制提供参考,为制定保护措施提供科学依据。
1 研究区概况与样地设置
1.1 研究区及样地概况
图1 样地基准点地理位置Fig.1 The location of the plot datum point
研究地点位于武夷山市星村镇四新附近(图1),该区域属典型的亚热带季风气候,年平均气温8.5—18 ℃,相对湿度78%—84%,年降水量一般为1486—2100 mm,无霜期253—272 d,土壤以红壤和黄壤为主。常绿阔叶林是武夷山地区最典型、最具代表性和最占优势的生态系统,主要由壳斗科、樟科、木兰科和山茶科等常绿双子叶阔叶树种组成,是中国中亚热带季风气候区的地带性植被类型,也是亚热带陆地生态系统的重要组成部分,主要分布于海拔1200 m以下。
样地基准点地理坐标为27°35′37″ N,117°45′54″ E,海拔约470—520 m。样地为正方形,投影面积为1.44 hm2(120 m×120 m),由山脊向山脚延伸。样地总体上北高南低,坡度10—50°,东南坡向为主;样地内有两条山谷,由北至南向穿越样地。乔木层主要有少叶黄杞(Engelhardiafenzelii)、米槠(Castanopsiscarlesii)、南岭栲(Castanopsisfordii)、甜槠(Castanopsiseyrei)、格药柃(Euryamuricata)、矩叶鼠刺(Iteaoblonga)、罗浮栲(Castanopsisfabri);灌木层伴生种主要有弯蒴杜鹃(Rhododendronhenryi)、秀丽野海棠(Brediaamoena)、密花树(Rapaneaneriifolia);草本层主要有狗脊蕨(Woodwardiajaponica)、苔草(Carextristachya)、草珊瑚(Sarcandraglabra)、芒萁(Discranoptetrisdichotama);层间层主要有网脉酸藤子(Embeliarudis)、链珠藤(Alyxiasinensis)、流苏子(Coptosapeltadiffusa)。由于该地较早被划为自然生态林加以保护,群落典型,地带性物种成分占绝对优势,近期未受较大的人为干扰,种群天然更新状况良好。仅在沟谷等不稳定地形上,由于滑坡等影响,间或有林窗分布。
1.2 样地概况及调查方法
2011年7月,在研究区内选择群落外貌具有代表性的常绿阔叶林,设置1.44 hm2(120 m×120 m)的永久性固定样地,用全站仪将整个样地划分36个20 m×20 m的大样方。在每个20 m×20 m的大样方内细分为16个5 m×5 m的小样方,以样地西南角为原点,东西向为横轴(x),南北向为纵轴(y),对植物群落进行全面调查。记录每个小样方内胸径(DBH)≥1 cm的树木个体及其分枝的位置、胸径、树高、枝下高和生长状况等,DBH≥1 cm的分枝按照同样方法挂牌测量,并记录分枝的母株编号。位置以坐标值表示,坐标值直接用距离(m)表示。
进一步对调查数据进行统计分析,分别计算出物种数量S、多度与密度、盖度,进而得到各物种的重要值,计算公式见文献[16];群落多样性的测度选用样地的Marglef物种丰富度指数、均匀度指数和物种多样性指数3类,具体测度方法见文献[17];按世界种子植物科、属的分布区类型划分标准[18- 20]统计样地内科、属的分布区类型;种-面积曲线采用随机取样法,分别用对数模型和幂指数模型拟合种-面积曲线;种群空间分布格局的分析方法选用扩散系数、Lloyd的平均拥挤度和聚块指数、负二项参数测度典型常绿阔叶林主要种群分布格局,其测定方法见文献[21]。采用R 2.15.0软件及SigmaPlot10.0对数据进行分析和作图。
2 结果
2.1 植物区系特征
2.1.1 种类组成
样地内乔木层共出现维管植物34科65属139种,其中仅杉木1种裸子植物,其它都为被子植物(双子叶植物32科63属137种,仅毛竹1种单子叶植物)。属、种数占优势的科主要有冬青科(1属15种)、樟科(5属13种)、山茶科(6属12种)、壳斗科(4属12种)、山矾科(1属10种)、杜鹃花科(3属9种)、蔷薇科(5属8种)、茜草科(5属5种)、木兰科(2属7种)、杜英科(1属5种)、木犀科(2属4种)。样地内独立个体数为9,116株(包括1,896个分枝),单位面积个体数为6,331株/hm2。下文统计的数据均基于独立个体。
样地内重要值>1%的物种有29种(表1),这些物种的个体数和胸高断面积分别占样地总个体数和总胸高断面积的78.08%和88.03%。样地内所有物种的平均胸径为5.38 cm,最大胸径为66.25 cm(米槠)。平均胸径最大的物种是南酸枣,为18.51 cm,但它只有7个个体;山桐子、拐枣和紫树平均胸径也较大,个体数并不多。样地内所有个体的总胸高断面积为34.01 m2/hm2。其中,胸高断面积大于2 m2/hm2的只有4个物种,从大到小依次为少叶黄杞、米槠、罗浮栲和南岭栲,它们的胸高断面积分别只占总胸高断面积的9.66%、9.35%、7.45%和6.88%。而个体数最多的赤楠的平均胸径为2.80 cm,胸高断面积仅占总胸高断面积的1.41%。
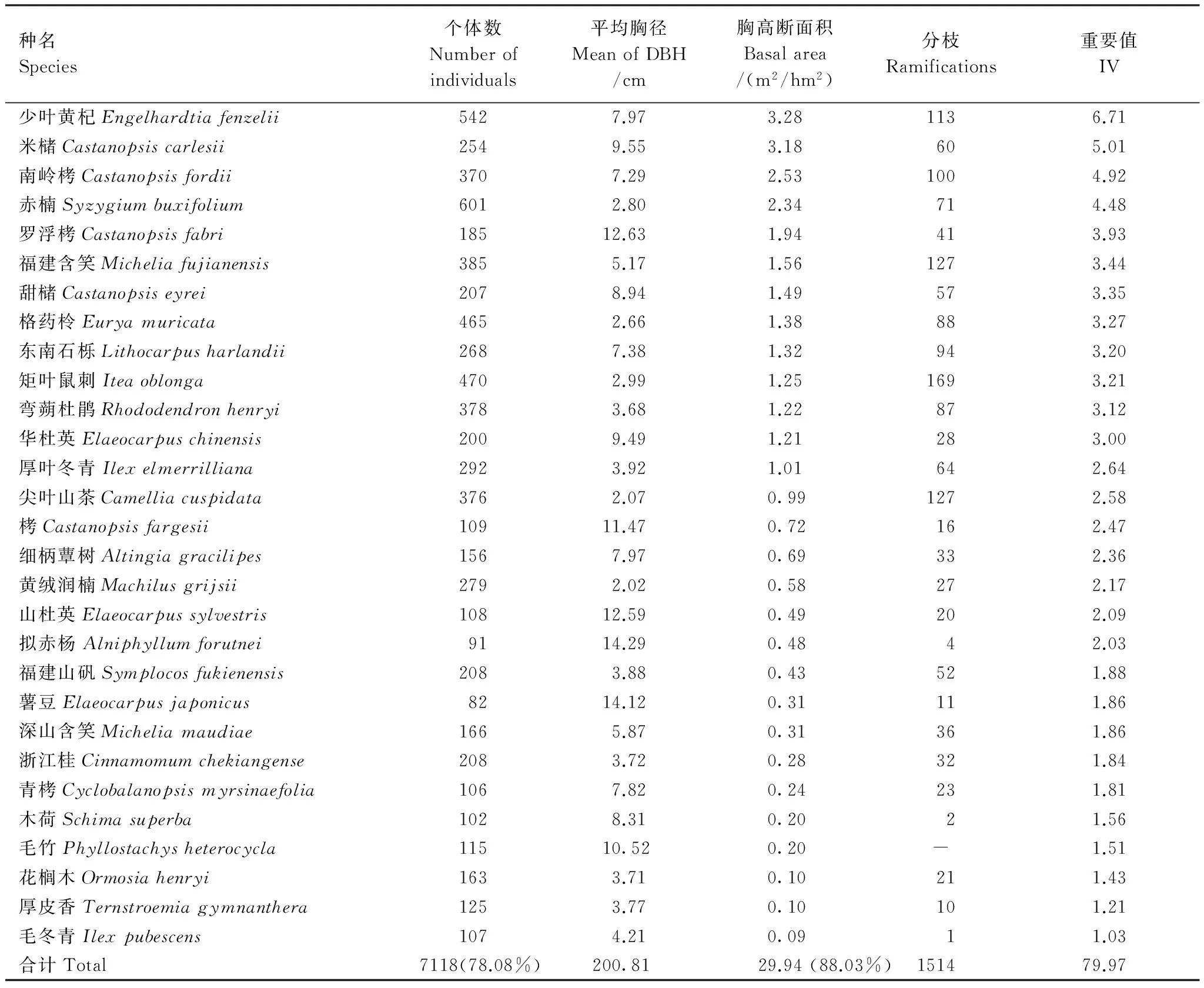
表1 样地中重要值≥1的优势物种组成Table 1 Species composition with important value≥1 in the plot
已有文献[22]将小于或等于1株/hm2的种定义为稀有种,1—10株/hm2的种定义为偶见种,大于100株/hm2的种为常见种。样地中共20个常见种,它们的个体数之和占样地总个体数的67.7%,其中个体数最多的赤楠(601株)占样地总个体数的6.6%,个体数最少的细柄蕈树(156株)占样地总个体数的1.7%。样地内有稀有种23种、偶见种46种,此69个物种的个体数之和只占样地总个体数的2.8%。
该样地物种丰富度指数Margalef指数R为15.14,说明群落中物种较丰富;Shannon-Wienner多样性指数H为5.60,反映群落不仅物种丰富,不同物种在群落内的分布较均匀。Simpson多样性指数D值为0.97,即群落的生态优势度值只有0.03,说明群落中没有绝对占优势的优势种,是较多的优势种形成了群落的共建种。物种均匀度指数Pielou指数为0.79,说明各个种的多度的均匀度值较大,即各物种个体数量的差异较小。总之,该群落的物种丰富度指数、均匀度指数和物种多样性指数说明样地内物种丰富,分布较均匀,多度没有集中于少数几个种,即较多的树种成了群落的优势种。
4) 所在楼层、公共交通、教育配套、生活配套对徐州市主城区的住宅价格影响不大.这可能是由于选择的研究区域为徐州市主城区,居民生活条件好,私家车拥有率高,学校分布均衡,基础设施完善.
2.1.2 分布区类型
由表2可知,科的分布区类型中,以泛热带分布最多(32.45%),如山樟科、茶科和山矾科等;其次是世界广布分布(26.50%),如蔷薇科、茜草科和木犀科等。总的来说,热带性质的科多于温带性质的科(18/7),这与武夷山地区地处中亚热带区系的特点相符。
属的分布区类型中,热带性质的属多于温带性质的属(39/24)。热带性质的属有39属,占60%,其中泛热带分布15属,占23.08%,如冬青属、卫矛属和红豆树属等,其次是热带亚洲分布10属,占15.38%,如青冈属、含笑属和木荷属等;旧世界热带分布6属,占9.23%,如五月茶属、野桐属和黄皮属等;热带亚洲和热带南美洲间断分布、热带亚洲至热带非洲分布及热带亚洲至热带非洲分布分别为3属、1属和4属,分别占4.62%、1.54%和6.15%。温带性质的属有24属,其中东亚和北美间断分布8属,占12.31%,如栲属、石栎属和漆属等;东亚分布7属,占10.77%,如山桐子属、刚竹属和南酸枣属等;旧世纪温带分布及地中海、西亚至中亚分布的各1属,占1.54%。中国特有属为2属:木兰属和杉木属。
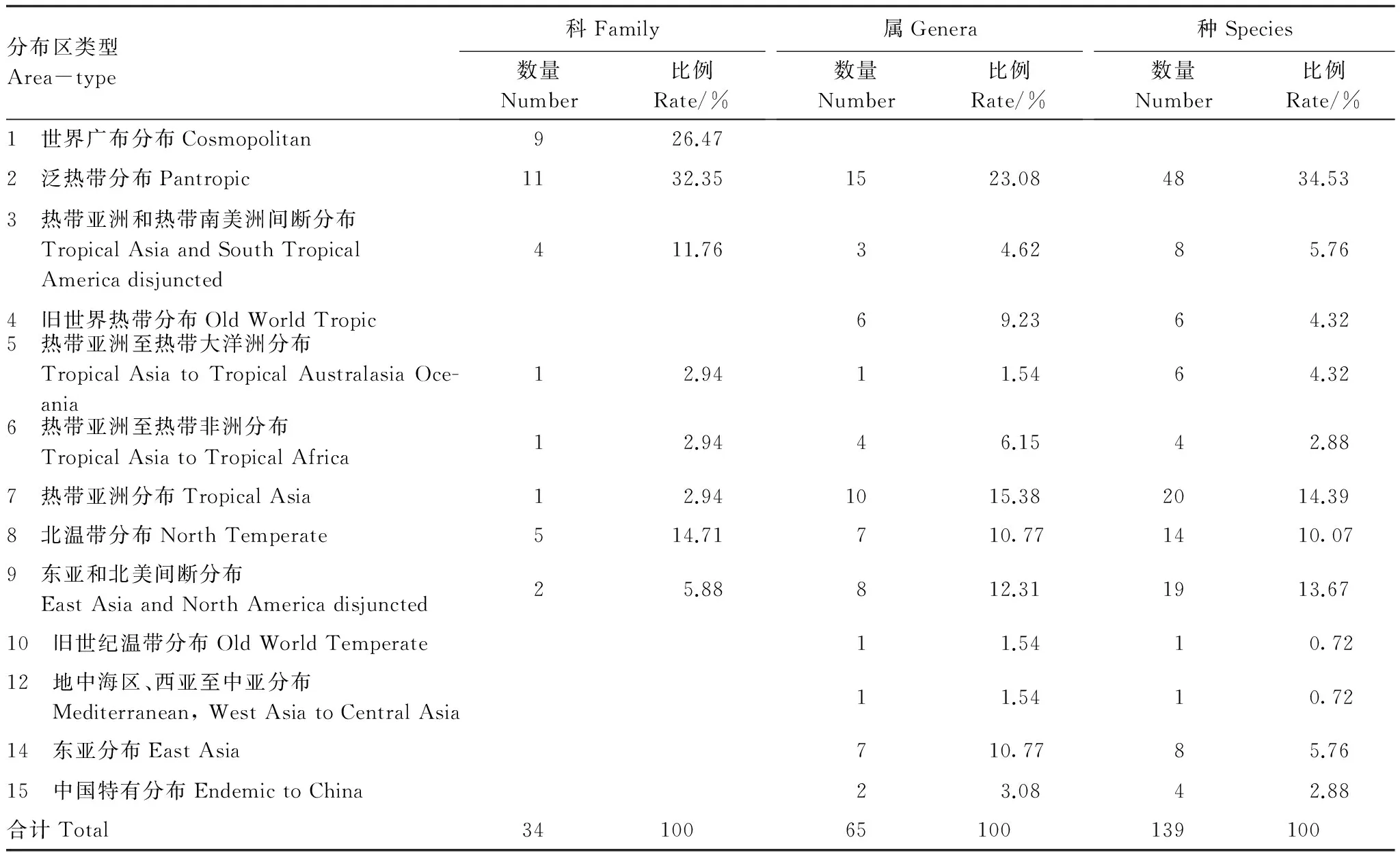
表2 样地中木本植物区系类型Table 2 The areal-types of woody plant in the plot
2.1.3 种-面积曲线
武夷山典型常绿阔叶林的种-面积曲线如图2所示,种-面积曲线在取样面积较小的初始阶段物种数迅速上升,当取样面积为0.48 hm2时,曲线斜率趋于稳定,物种数随取样面积增加缓慢,说明该植物群落物种十分丰富。面积在0.48 hm2以下时,物种随着面积增加而迅速增加,有111个物种出现,占样地总物种80%;面积在0.48—1.44 hm2之间,物种数增加比较缓慢;当取样面积达到1.28 hm2时,样地内的48个物种全部出现。种-面积曲线用取样面积(A)自然对数的线性方程拟合,拟合公式为:
S= 21.48lnA- 71.32 (R2= 0.978)
(1)
式中,S为物种数,A为样方面积。
2.3 径级结构
径级分布是反映林分数量特征的一个重要指标。根据样地内林木胸径径级分布的实际情况进行划分,共分为3级:小径木、中径木和大径木,小径木指的是1 cm≤DBH<6 cm的树木;中径木指的是6 cm≤DBH<20 cm的树木;大径木指的是DBH≥20 cm的树木。由表3可知,随着林木胸径的增大,样地内林木株数呈迅速减少的趋势,林木径级分布遵从异龄林典型的倒“J”型(图3),不同径级的个体数和物种数差异较大。整个样地内大、中、小径木分别占总株数的3.89%、22.21%和73.89%,小径木的比例较大,大径木所占比例较小。不同等级的物种数从多到少的顺序是小径木(130种)>中径木(90种)>大径木(37种)。
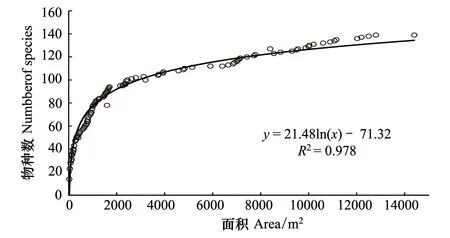
图2 样地种-面积曲线Fig.2 Species-area curve in the plot
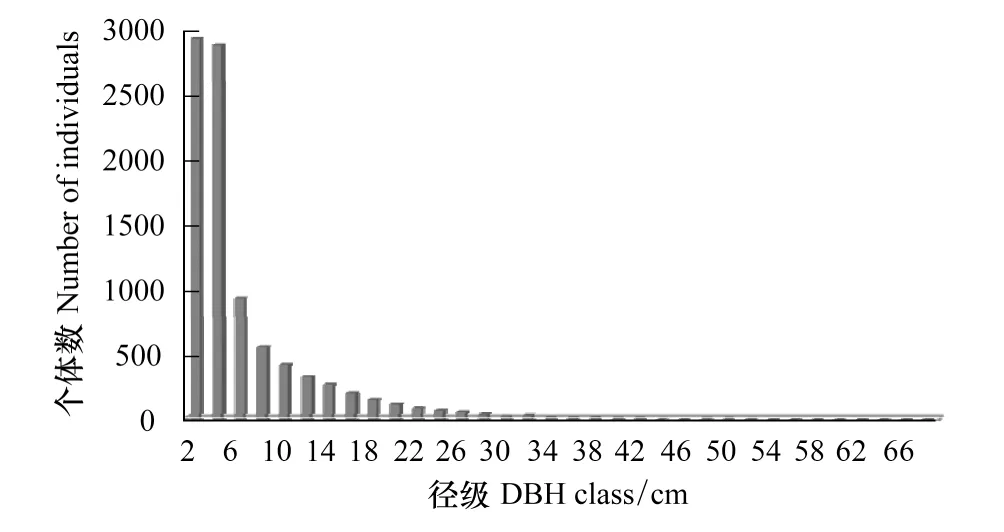
图3 样地木本植物径级分布Fig.3 Distributional patterns of DBH class in the plot
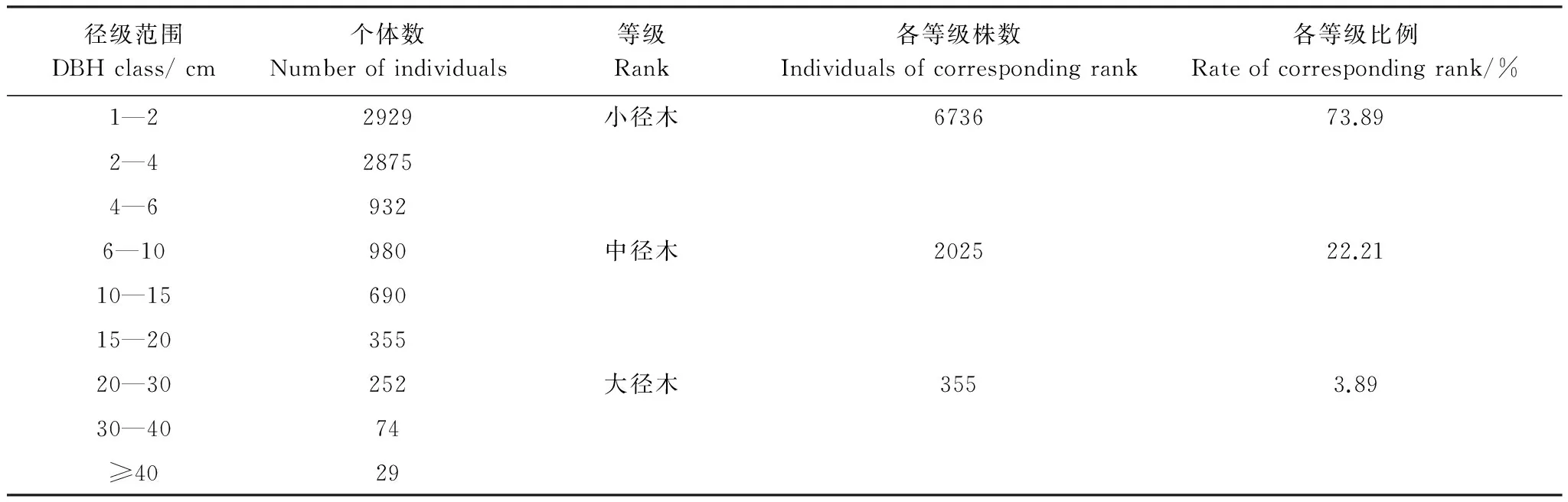
表3 DBH≥1 cm的林木径级分布Table 3 Size-class distribution of the woody palnts (DBH≥1cm) in the plot
根据大、中、小径木3级划分得到各主要树种的径级分布,列出重要值大于3的12个树种在胸径大小结构的一般特征,包括各主要树种的(小径木、中径木和大径木)的径级分布和绝对最大胸径以及平均胸径(表4)。绝对最大胸径指样地内该树种最大的一株树的胸径。从表4可知,以小径木株数的百分率来看,各树种从多到少依次为:格药柃(99.8)、矩叶鼠刺(99.6)、赤楠(96.0)、弯蒴杜鹃(95.8)、福建含笑(87.5)、南岭栲(78.6)、东南石栎(73.5)、少叶黄杞(70.0)、米槠(68.9)、甜槠(64.7)、华杜英(59.0)、罗浮栲(45.9)等。进一步对样地内重要值前4的少叶黄杞、米槠、南岭栲和赤楠的径级结构分析可以看出(图4),这4个种群的个体都集中在小径级,中径木和大径木的个体数相对较少。对于少叶黄杞和南岭栲,中径木的个体数都明显高于大径木的个体数;对于米槠,其中径木和大径木的个体数相当;而对于赤楠,小径木的个体数占总其总个体数的96%,中径木为4%,这可能与赤楠的生态学特性有关。总的来说,南岭栲和米槠种群在林木层和亚乔木层中占优势,赤楠种群在灌木层中占绝对优势。
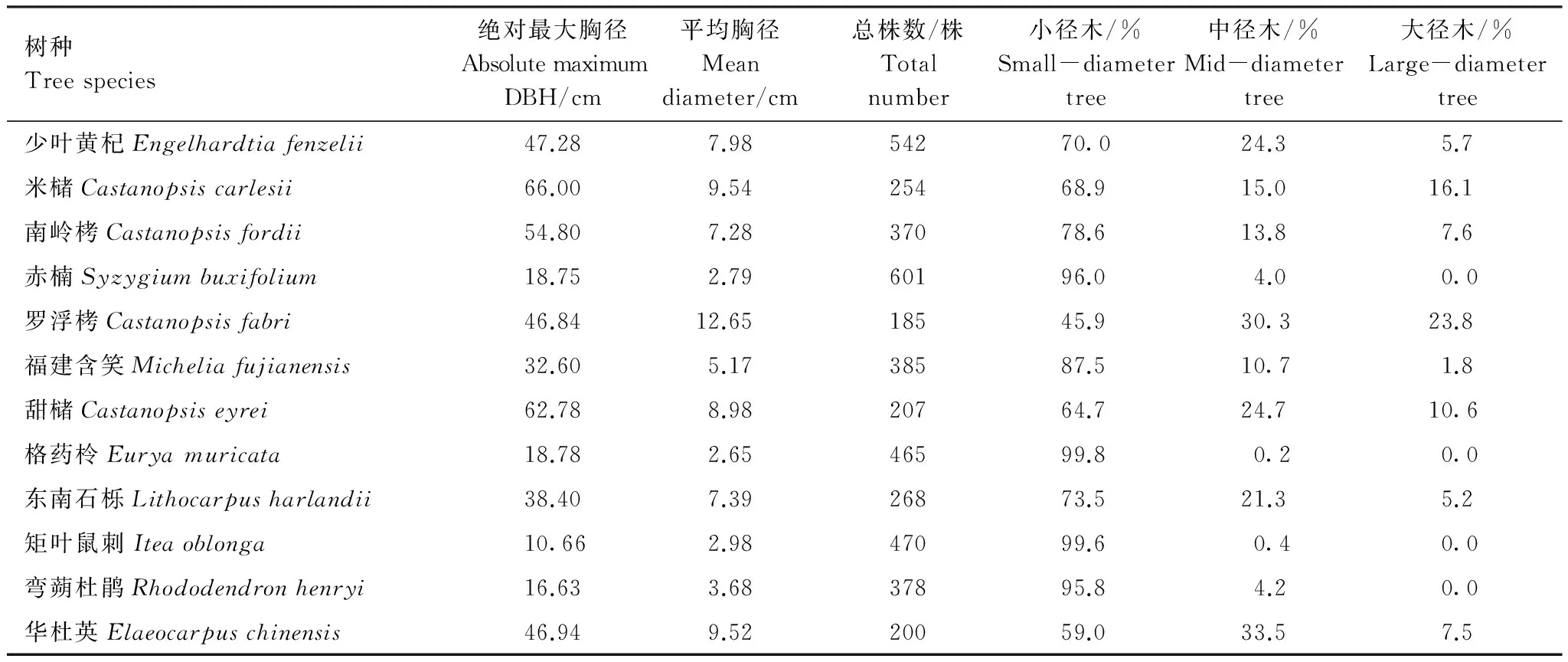
表4 重要值>3的12个树种径级分布Table 4 Size-class distribution of 12 species with importance value lager than 3
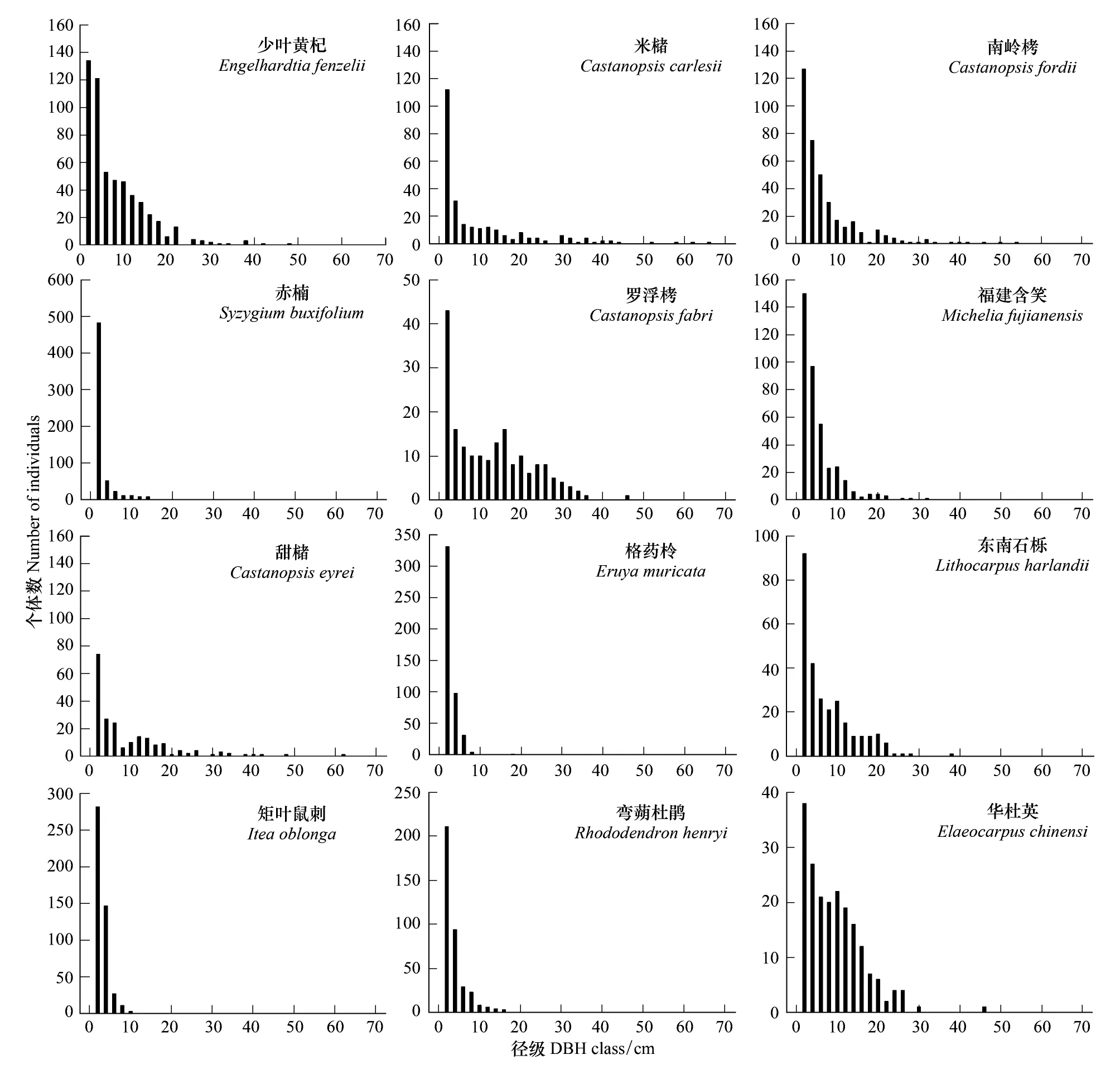
图4 样地中12个主要物种的径级分布Fig.4 Size-class distribution of twelve main species in the plot
2.4 空间分布格局
种群空间格局反映的是一定环境因子对个体行为、生存和生产的影响。不仅与物种的生物学特性和种间竞争有关,而且与生境有密切的联系。大多数自然种群都服从集群分布而不服从随机分布。对武夷山重要值>3的12个树种进行空间格局分析(表5,图5)。从表5中可以看出,少叶黄杞种群的方差/均值比为3.23,大于1,且t值检验差异极显著;平均拥挤度为3.37,大于均值0.94;聚块指数为3.58,大于1;负二项参数为0.39,在0—3范围内。以上结果均表明少叶黄杞种群为集群分布。同样,米槠等11个种群的方差/均值比均大于1,t值均大于0.48,差异极显著;平均拥挤度均大于各自的均值;聚块指数均大于1;负二项参数值均在0—3范围内。因此,这12个种群的分布格局都呈集群分布,但集群度有所差异,集群度从高到低依次为:甜槠、福建含笑、东南石栎、矩叶鼠刺、华杜英、格药柃、米槠、少叶黄杞、罗浮栲、赤楠、南岭栲和弯蒴杜鹃。从图5中可以看出,12个种群呈小斑块镶嵌式或有交错的镶嵌式分布,表现出小气候或小生境的集群分布。这种现象的主要原因是除受到种群本身的生态学特性和外界生态环境综合影响外,还与种子的成群散布有关。果实成熟后掉落在母株周围,只能靠重力作用进行传播,致使在母株周围形成大量的幼树,就造成了这些种群在一定地域的集群分布。

表5 重要值>3的12个树种的分布格局分析表Table 5 The distribution patterns′ analysis table of 12 species with importance value lager than 3
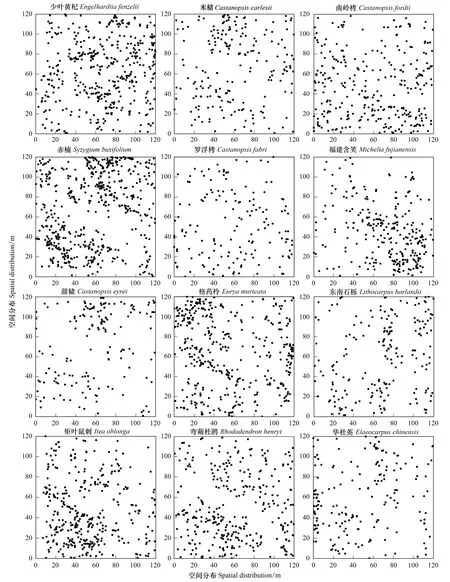
图5 重要值大于3的12个树种的空间分布格局Fig.5 Spatial distribution of 12 species with importance value lager than 3
为了进一步说明种子扩散是致使12个种群在一定地域集群分布的主要原因,对样地内4个主要树种的小、中、大径木的空间分布进行分析发现(表6,图6),这4个主要种群在不同等级的径级下都表现出了集群分布,集群度有所差异。少叶黄杞、米槠、南岭栲和赤楠种群的集群度随径级从高到底的顺序分别为:小径木>大径木>中径木、小径木>中径木>大径木、小径木>中径木>大径木、中径木>小径木。少叶黄杞、米槠和南岭栲3个种群的小径木的集群度最高。这是因为集群分布能更好地改变小生境,小径木对不良环境比单独个体抗性更强,更有利于小径木围绕母株聚集分布[23]。而赤楠种群的集群度并不是小径木最高的理想情况,主要是因为空间格局除受种子扩散影响外,还受到种间竞争的影响。样地内赤楠种群的个体总体上较小,平均胸径只有2.79 cm,小径木的比例占其总个体数的96%,其集群度的差异受其他种群干扰的影响。
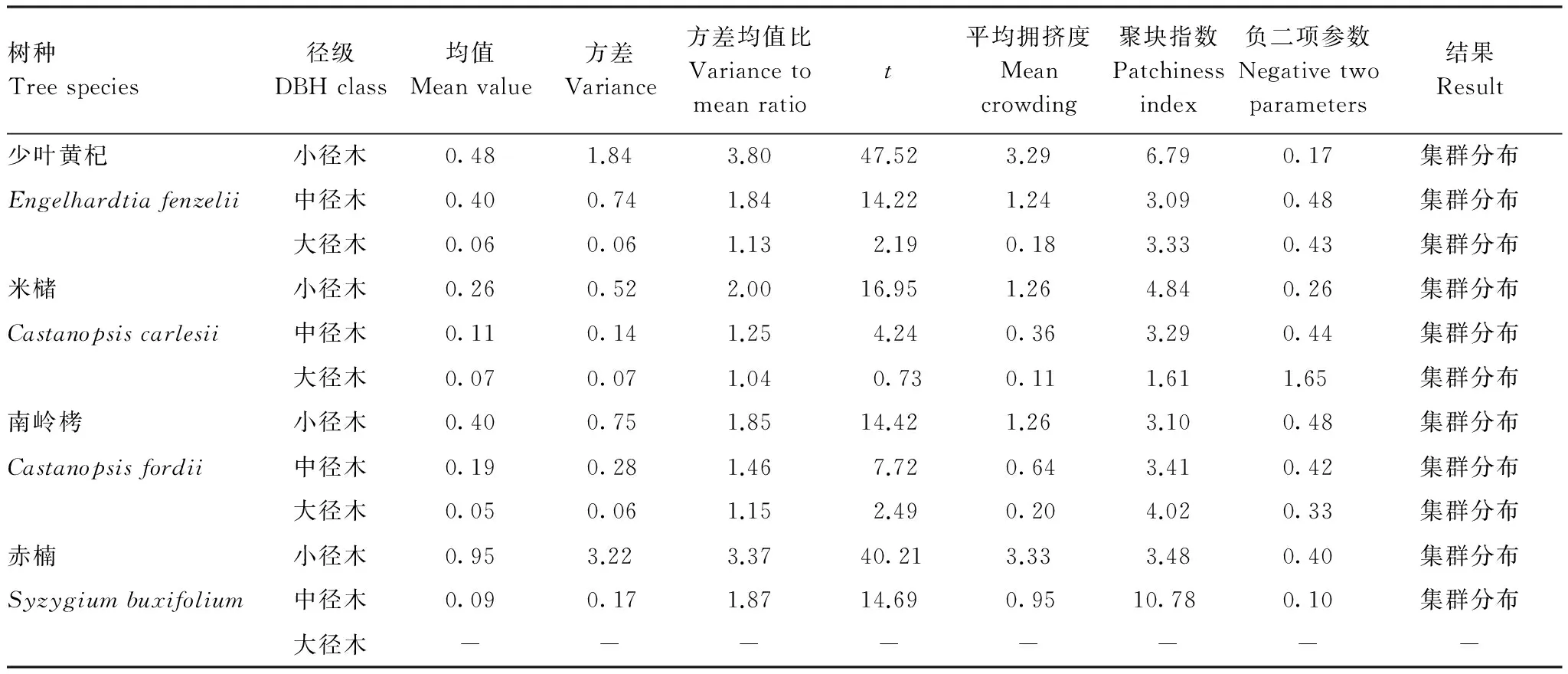
表6 4个主要树种的格局分析Table 6 The patterns analysis of 4 main species
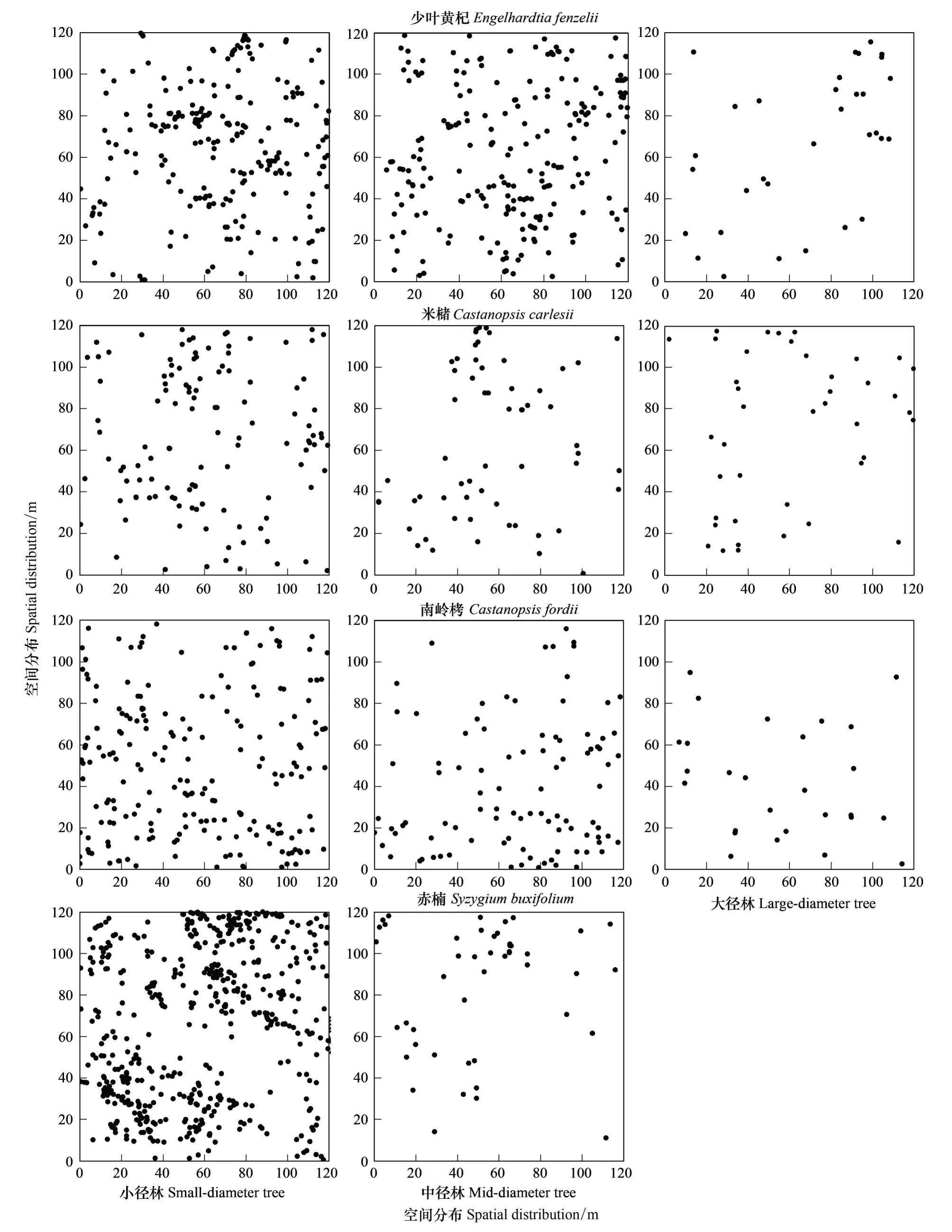
图6 4个主要树种在不同径级的空间分布格局Fig.6 Spatial distribution of 4 main species at different size-classes
3 讨论
监测种群生长、死亡和更新动态是一个非常重要和复杂的生态学过程。本文所建设的1.44 hm2样地相比BCI的50 hm2、古田山24 hm2、浙江天童20 hm2和小兴安岭9 hm2[24]样地很小,但是根据宋永昌[25]关于中亚热带常绿阔叶林最小取样面积为400—800 m2,Hubbell26]和祝燕等[12]关于小面积条件下研究个体数大于100株/hm2的植物种群才满足监测要求,武夷山样地中有139个物种,其中20个物种大于100株/hm2,已足以监测样地内主要物种的动态变化。通过对武夷山典型常绿阔叶林样地物种组成与群落结构的分析,可以发现:样地内共有维管束植物34科65属139种,其中仅杉木1种裸子植物,其余都为被子植物,样地内有国家Ⅲ级保护植物乐东拟单性木兰;区系特征明显,泛热带成分占主要成分;优势树种不明显。
与浙江天童20 hm2和古田山24 hm2常绿阔叶林样地相比,在物种组成和群落结构等方面有很多共同之处:树木个体密度相近,武夷山、天童和古田山样地每公顷树木个体数分别为6331、6627和5863株/hm2;科和属的分布区类型相近,3样地科和属的分布区类型热带性质均高于温带性质成分,常绿阔叶树占绝对优势;小径级树木占绝对优势,武夷山样地DBH<6 cm的个体数占总个体数的73.89%,天童样地DBH<5 cm的个体数占总个体数的70.3%,古田山样地DBH<4 cm的个体数占总个体数的69.07%;武夷山样地与古田山样地,4个主要优势种群都表现一定程度的集聚分布,且都随径级的增大集聚度呈降低的趋势。这些与三地同属中亚热带常绿阔叶林,地理位置和植被类型较为接近有很大的联系。除以上共性外,三地在物种组成上也有明显的差异。天童和古田山样地都有其较为明显的优势树种,而武夷山样地出优势树种不明显的特征;3样地都有一定数量的稀有种,武夷山、天童和古田山样地稀有种数量(比例)分别为23种(16.5%)、55种(36.2%)和59种(37.1%)。
种群空间格局反映的是一定环境因子对个体行为、生存和生产的影响。一般来说,种群分布的状态和形式,可分为随机分布、集群分布和均匀分布3种类型。集群分布体现了有利于个体间生存的空间关系,而均匀分布体现了种内某些个体会对其他个体的生存产生不利影响的关系,随机分布则意味着种内个体间相互独立[27]。通过选择武夷山典型常绿阔叶林群落中重要值大于3的12个植物种群间和4个主要树种不同径级的空间结构研究,发现研究区域内这12个植物群落和这4个主要种群的大、中、小径木基本上呈集群分布,这与郑元润[21]、吴宁[28]和闰淑君[29]的研究结果一致,即绝大数自然种群都服从集群分布而不服从随机分布。这可能与种子散布的限制或生物学特性有关。4个主要树种的集聚度总体上表现出从小径木到中径木直到大径木逐渐降低的趋势,这与祝燕[12]、He[30]等的研究结果一致。
在本研究中,基于武夷山典型常绿阔叶林1.44 hm2动态监测样地的调查,较全面地介绍了样地内的物种组成与群落结构,探讨了重要值大于3的12个植物种群和4个主要种群大、中、小径木的空间格局,对解决生物多样性形成和维持机制、生物多样性保护等重要生态学问题和实践问题具有重要的意义。地形是影响植物科利用土壤各种养分的重要因素,本文缺少分析地形和环境异质性对物种空间格局的影响,有必要进一步进行种群空间分布格局研究。
致谢:南京林业大学李蒙、严靖、沈超、左美银、黄建等同学和南京森林警察学院杜莹老师参加野外调查工作,环境保护部南京环境科学研究所姚纲博士、雷军成博士帮助写作,特此致谢。
[1] Xu H G, Wu J, Liu Y, Ding H, Zhang M, Wu Y, Xi Q, Wang L L. Biodiversity congruence and conservation strategies: A national test. BioScience, 2008, 58(7): 632- 639.
[2] Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Biodiversity Synthesis. Washington DC: World Resources Institute, 2005: 3- 3.
[3] Pimm S L, Russell G J, Gittleman J L, Brooks T M. The future of biodiversity. Science, 1995, 269(5222): 347- 350.
[4] 吴军. 《生物多样性公约》的产生背景和主要内容. 绿叶, 2011, (9): 47- 54.
[5] 郝占庆, 张健, 李步杭, 叶吉, 王绪高, 姚晓琳. 长白山次生杨桦林样地: 物种组成与群落结构. 植物生态学报, 2008, 32(2): 251- 261.
[6] 郝占庆, 李步杭, 张健, 王绪高, 叶吉, 姚晓琳. 长白山阔叶红松林样地(CBS): 群落组成与结构. 植物生态学报, 2008, 32(2): 238- 250.
[7] 刘海丰, 李亮, 桑卫国. 东灵山暖温带落叶阔叶次生林动态监测样地: 物种组成与群落结构. 生物多样性, 2011, 19(2): 232- 242.
[8] 叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 2008, 32(2): 274- 286.
[9] 吴征镒. 中国植被. 北京: 科学出版社, 1980: 306- 356.
[10] 何永涛, 曹敏, 唐勇, 杨国平. 滇中地区常绿阔叶林树种多样性比较研究. 山地学报, 2000, 18(4): 322- 328.
[11] 陈昌笃. 论武夷山在中国生物多样性保护中的地位. 生物多样性, 1999, 7(4): 320- 326.
[12] 祝燕, 赵谷风, 张俪文, 沈国春, 米湘成, 任海保, 于明坚, 陈建华, 陈声文, 方腾, 马克平. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构. 植物生态学报, 2008, 32(2): 262- 273.
[13] 杨庆松, 马遵平, 谢玉彬, 张志国, 王樟华, 刘何铭, 李萍, 张娜, 王达力, 杨海波, 方晓峰, 阎恩荣, 王希华. 浙江天童20ha常绿阔叶林动态监测样地的群落特征. 生物多样性, 2011, 19(2): 215- 223.
[14] 陈辉, 刘玉宝, 吴承祯, 游斌. 闽北次生常绿阔叶林主要树种空间分布格局及其应用研究. 应用与环境生物学报, 1999, 5(6): 561- 565.
[15] 毕肖锋, 彭华贵, 黄忠良, 曹洪麟. 南岭大顶山常绿阔叶林群落结构及其物种多样性. 生态科学, 2005, 24(2): 113- 116.
[16] 孙儒泳, 李庆芬, 牛翠娟, 娄安如. 基础生态学. 北京: 高等教育出版, 2002: 140- 143.
[17] 张金屯. 数量生态学 (第二版). 北京: 科学出版社, 2011: 248- 249.
[18] 吴征镒, 周浙昆, 李德铢, 彭华, 孙航. 世界种子植物科的分布区类型系统. 云南植物研究, 2003, 25(3): 245- 257.
[19] 吴征镒. 《世界种子植物科的分布区类型系统》的修订. 云南植物研究, 2003, 25(5): 535- 538.
[20] 吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991, (增刊): 1- 139.
[21] 郑元润. 不同方法在沙地云杉种群分布格局分析中的适用性研究. 植物生态学报, 1997, 21(5): 480- 484.
[22] Hubbell S P, Foster R B. Commonness and rarity in a neotropical forest: implications for tropical tree conservation // Conservation Biology: the Science of Scarcity and Diversity. Sunderland: Sinauer Press, 1986: 205- 231.
[23] 宋萍, 洪伟, 吴承祯, 封磊, 范梅兰, 朱慧, 林勇明, 张琼. 珍稀濒危植物桫椤种群结构与动态研究. 应用生态学报, 2005, 16(3): 413- 418.
[24] 徐丽娜, 金光泽. 小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构. 生物多样性, 2012, 20(4): 470- 481.
[25] 宋永昌. 植被生态学. 北京: 华东师范大学出版社, 2001.
[26] Hubbell S P. Two decades of research on the BCI Forest Dynamics Plot: where we have been and where we are going // Losos E C, Leigh E G Jr, eds. Tropical Forest Diversity and Dynamism: Findings From a Large-Scale Plot Network. Chicago: University of Chicago Press, 2004: 8- 30.
[27] 宋于洋, 李园园, 张文辉. 梭梭种群不同发育阶段的空间格局与关联性分析. 生态学报, 2010, 30(16): 4317- 4327.
[28] 吴宁. 贡嘎山麦吊杉群落优势种群的分布格局及相互关系. 植物生态学报, 1995. 19(3): 270- 278.
[29] 闰淑君, 洪伟, 吴承祯, 毕晓丽, 蓝斌. 武夷山天然米槠林优势种群结构与分布格局. 热带亚热带植物学报, 2002, 10(1): 15- 21.
[30] He F L, Legendre P, LaFrankie J V. Distribution patterns of tree species in a Malaysian tropical rain forest. Journal of Vegetation Science, 1997, 8(1): 105- 114.
Species composition and community structure of the typical evergreen broad-leaved forest in the Wuyi Mountains of Southeastern China
DING Hui1,*,YANG Yunfang1,2, XU Haigen1, FANG Yanming3, CHEN Xiao4, YANG Qing4, YI Xiangui3, XU Hui4, WEN Xiaorong3, XU Xianjun4
1NanjingInstituteofEnvironmentalSciences,MinistryofEnvironmentalProtection,StateEnvironmentalProtectionKeyLaboratoryonBiosafety,Nanjing210042,China2InstituteofRemoteSensing,NanjingUniversityofInformationScience&Technology,Nanjing210044,China3CollegeofForestResourcesandEnvironment,NanjingForestUniversity,Nanjing210037,China4WuyishanBiologicalInstituteofFujianProvince,Wuyishan354300,China
Subtropical evergreen broad-leaved forest typically forms in subtropical areas of the eastern coasts of continents which usually experience moist and monsoon climate conditions. This kind of forest is one of several representative vegetation types in China, which has the largest area covered by this kind of vegetation in the world. Even though subtropical evergreen broad-leaved forest is one of the key areas of biodiversity in China and is characteristic of much of the Wuyi Mountains, few studies have reported the species composition and community structure of this mountain range. Recently, dynamic monitoring of forest biodiversity has attracted increased attention of scientists and is known to provide detailed data for understanding the species composition and the community structure of these forests. In July, 2011, a 1.44 hm2(120 m × 120 m) permanent forest plot (27°35′37″N, 117°45′54″E, alt. 470—520 m) was established in the Wuyi Mountains for long-term monitoring of plant diversity. Parameters for each individual woody plant with DBH ≥ 1 cm was recorded, including the number of individuals in the plot, position, diameter at breast height (DBH), plant height, height under living branch, forest health condition and so on. Using our field data, the species composition and community structure of this plot were analyzed based on the species composition, floristic characteristics, size class structure and spatial patterns of the area. A total of 9,116 individuals (1896 branches of trees with trunks in the plots were also included) were found, belonging to 139 species, 65 genera and 34 families. Of the 139 species found, only one was a gymnosperm (Cunninghamialanceolata(Lamb.) Hook.) and only one was a monocot (Phyllostachysedulis(Carrière) J. Houzeau); the rest were dicots. Twenty-three rare species (≤ 1 individual / hm2) and 46 accidental species (1—10 individuals / hm2) were found in the plot, individuals of which together accounted for 2.8% of the total individuals in the plot. For common species (>100 individuals / hm2),SyzygiumbuxifoliumHook. & Arn. was most common with 601 individuals or 6.6% of the individuals in the plot, whileAltingiagracilipesHemsl. had the fewest individuals (156, 1.7%) for a common species. No species appeared to be dominant in the plant community, as suggested by the abundance, basal area, the average DBH and importance values of the 139 species. However, the floristic characteristics of the plant community were very important; strong temperate aerial-type species were found at both the family- and genus-level, and a few tropical aerial-type species were intermixed. The size distribution of 12 species with importance value that higher than 3 followed a reverse “J” shape. An evident difference was observed between the number of individuals and species that present with different size classes. For the entire plot, only 3.89% of all individuals had a large DBH (> 20 cm), while 73.89% of all individuals had a small DBH (1 cm ≤ DBH < 6cm). In addition, spatial pattern analysis showed the 12 tree species with high importance values and the large-diameter, mid-diameter and small-diameter trees of the four main species (EngelhardiafenzeliiMerr.,Castanopsiscarlesii(Hemsl.) Hayata,CastanopsisfordiiHance,S.buxifolium) were aggregated, and aggregation intensity decreased as DBH increased.
typical evergreen broad-leaved forest; species composition; size class structure; spatial patterns; Wuyi Mountains
国家科技支撑计划课题(2012BAC01B01)
2013- 05- 05;
日期:2014- 04- 11
10.5846/stxb201305050924
*通讯作者Corresponding author.E-mail: nldinghui@sina.com
丁晖,杨云方,徐海根,方炎明,陈晓,杨青,伊贤贵,徐辉,温小荣,徐鲜均.武夷山典型常绿阔叶林物种组成与群落结构.生态学报,2015,35(4):1142- 1154.
Ding H,Yang Y F, Xu H G, Fang Y M, Chen X, Yang Q, Yi X G, Xu H, Wen X R, Xu X J.Species composition and community structure of the typical evergreen broad-leaved forest in the Wuyi Mountains of Southeastern China.Acta Ecologica Sinica,2015,35(4):1142- 1154.
