蚯蚓对瓦氏黄颡鱼摄食达氏鳇卵的干扰实验
2015-03-10石小涛白艳勤胡运燊陈求稳刘德富
石小涛, 路 波, 白艳勤, 胡运燊, 罗 佳, 陈求稳,3, 乔 晔, 刘德富,*
1 三峡大学,三峡库区生态环境教育部工程研究中心, 宜昌 443002 2 水利部水工程生态效应与生态修复重点实验室, 武汉 430079 3 中国科学院生态环境研究中心, 北京 100085
蚯蚓对瓦氏黄颡鱼摄食达氏鳇卵的干扰实验
石小涛1,2,3, 路 波1, 白艳勤1, 胡运燊1, 罗 佳1, 陈求稳1,3, 乔 晔2, 刘德富1,*
1 三峡大学,三峡库区生态环境教育部工程研究中心, 宜昌 443002 2 水利部水工程生态效应与生态修复重点实验室, 武汉 430079 3 中国科学院生态环境研究中心, 北京 100085
自然水域中中华鲟(Acipensersinensis)卵常受到敌害鱼类捕食的威胁,瓦氏黄颡鱼Pelteobagrusvachelli(Richerdson)是中华鲟卵的重要敌害鱼类。以中华鲟卵的敌害鱼类瓦氏黄颡鱼为研究对象,选择环毛蚓(Pheretimatschiliensis)为瓦氏黄颡鱼的饵料,以达氏鳇卵(Kalugasturgeon)替代中华鲟卵进行室内实验,通过研究适合瓦氏黄颡鱼摄食的环毛蚓规格,以及环毛蚓干扰瓦氏黄颡鱼摄食达氏鳇卵投放比例,探讨中华鲟卵的保护方法。在水温为(20.0±1.5) ℃的条件下,研究了3种不同体长规格[体长(15.3±1.4) cm、(12.0±2.1) cm、(7.7±0.5) cm]实验鱼个体分别摄食3种不同体长规格环毛蚓(体高为0.3cm,体长分别为8、4、2cm)和达氏鳇卵的摄食率;研究了体长为4cm的环毛蚓对3种不同体长实验鱼摄食达氏鳇卵的干扰效果。实验结果表明:3种体长规格瓦氏黄颡鱼均摄食达氏鳇卵;3种体长规格瓦氏黄颡鱼共同摄食的替代饵料规格为体长4cm及以下的环毛蚓;适宜干扰实验所选体长实验鱼摄食达氏鳇卵的饵料为体长4 cm的环毛蚓;当环毛蚓与达氏鳇卵数量比例为1∶1时,可实现达氏鳇卵的保护。研究证明在实验条件下可实现环毛蚓对中华鲟卵敌害鱼类——瓦氏黄颡鱼实施摄食干扰,提示下一步可以尝试选择环毛蚓作为替代中华鲟卵的饵料,进行干扰食卵鱼类摄食中华鲟卵的野外验证试验。
瓦氏黄颡鱼; 环毛蚓; 达氏鳇卵; 摄食
葛洲坝建成后阻断了中华鲟(Acipensersinensis)繁殖洄游通道,导致中华鲟产卵场缩减和中华鲟补给资源量急剧下降[1]。此外,中华鲟卵在孵化过程中还遭受到敌害鱼类捕食,进一步导致中华鲟补给资源量下降[2]。2002年的调查表明,90%的中华鲟卵资源量被底栖性敌害鱼类所摄食,敌害鱼类主要包括圆口铜鱼(Coreiusguichenoti)、铜鱼(Coreiusheterodon)和瓦氏黄颡鱼Pelteobagrusvachelli(Richerdson)等[1- 2]。2011年的调查发现,在中华鲟产卵时期中华鲟产卵场中瓦氏黄颡鱼的分布资源量比例增加,占总食卵鱼类70%以上[2]。因此,瓦氏黄颡鱼可能成为了中华鲟卵的主要敌害鱼类。
瓦氏黄颡鱼又称江黄颡鱼,隶属鲇形目(Sihrriforms)、鲿科(Bagridea)、黄颡鱼属(PelteobagrusBleeker),是长江水系的重要经济鱼类[3]。目前,关于瓦氏黄颡鱼的研究,主要集中在对人工养殖瓦氏黄颡鱼的生理和行为研究,如天然饵料缺失对瓦氏黄颡鱼鱼苗的培育存在影响[4],温度和光照影响瓦氏黄颡鱼幼鱼对黑白底质颜色的偏好和集群行为[5]。关于长江中瓦氏黄颡鱼野生个体的研究主要集中在其资源量调查上,而对其摄食习性报道甚少。为了进一步了解瓦氏黄颡鱼对中华鲟卵的捕食效应并寻找潜在的危害减缓措施,有必要针对长江葛洲坝下游江段瓦氏黄颡鱼的摄食行为展开研究。
鱼类摄食饵料的适口性和摄食偏好是鱼类摄食习性的重要内容[6- 9],如路波等[10]在比较瓦氏黄颡鱼对多种饵料的选择性后发现蚯蚓是瓦氏黄颡鱼的最喜食饵料。同时,有研究报道鱼类在面对多种饵料时一般选择丰度相对较高的饵料[11- 13]。因此本研究以环毛蚓(Pheretimatschiliensis)为替代饵料,探讨替代饵料和目标饵料的相对数量比例对瓦氏黄颡鱼摄食的影响,以期通过替代饵料干扰瓦氏黄颡鱼的摄食实现鲟鱼卵的保护。
1 材料与方法
1.1 实验材料及其驯化
瓦氏黄颡鱼为渔民在葛洲坝下游中华鲟产卵场捕获的野生个体。实验前暂养30 d,暂养在黑色圆形水槽中,水槽直径2 m,深1 m。实验鱼用浓度为4—8 mg/L的高锰酸钾溶液消毒5 min后放入暂养水槽。暂养水为曝气3 d后的自来水,每3日更换1次,水温(20.0±1.0) ℃,溶氧>7 mg/L,氨氮<0.01 mg/L, 室外自然光,暂养7 d。为防止实验鱼对某种饵料适应,实验鱼暂养期间不投喂饵料。
1.2 实验设计
实验鱼被放置在深蓝色水族箱进行实验,水族箱为(宽0.4 m×深0.4 m×长0.6 m)。实验水深0.2 m,水温为(20.0±1.5) ℃。实验鱼根据体长分为3种规格:(15.3±1.4) cm (大)、(12.0±2.1)cm (中)、(7.7±0.5) cm (小)。实验选择体长(8.3±0.4) cm(以下以8 cm代表该体长数据)、(4.5±0.7) cm(4 cm代表该体长数据)、(2.1±0.6) cm(2 cm代表该体长数据)的环毛蚓和中华鲟卵替代品——达氏鳇(Kalugasturgeon)卵[卵径为(0.43±0.03) cm]作为瓦氏黄颡鱼的饵料进行实验,实验选择达氏鳇卵作为中华鲟卵的替代品,是因为中华鲟卵资源量不易获得和达氏鳇卵与中华鲟卵理化性质相似。研究主要进行单因素对瓦氏黄颡鱼摄食行为的影响。瓦氏黄颡鱼摄食适宜饵料和达氏鳇卵的实验,实验选择同一体长规格实验鱼被投喂同一种饵料为一个实验组,3种体长规格实验鱼,4种饵料(3种体长规格环毛蚓+达氏鳇卵),共12个实验组。单独投喂环毛蚓时,每次实验选择3组不同体长实验鱼同时投喂同种体长规格环毛蚓,3次重复,每次重复5尾实验鱼,共135尾实验鱼;单独投喂达氏鳇卵时,投饵量以实验鱼平均体重的3%—5%为准,大规格体长实验鱼被投喂25粒(1.535±0.046) g,中规格体长实验鱼被投喂20粒(1.220±0.037) g,小规格体长实验鱼被投喂15粒(0.915±0.052) g,每次选择3组不同体长实验鱼同时进行实验,每组3次重复,每次重复5尾实验鱼,共45尾。根据瓦氏黄颡鱼摄食不同体长环毛蚓的实验结果,发现试验所选体长瓦氏黄颡鱼均摄食4 cm体长规格的环毛蚓,且相比2 cm体长环毛蚓达到相同饱和度花费寻觅饵料时间短。因此本研究选择4 cm体长的环毛蚓干扰瓦氏黄颡鱼摄食达氏鳇卵的实验,同样选择以上3种不同体长大小实验鱼,饵料以4 cm体长环毛蚓和达氏鳇卵为主,进行了2种不同投放数量比例的实验组,3种体长规格实验鱼同时被投喂的两种投放数量比例相同的饵料,同样分别被投喂两组不同投放数量比例的饵料。每组实验重复3次,每次重复5尾实验鱼,实验鱼共90尾。实验组达氏鳇卵和4 cm体长环毛蚓的数量比例分别为(达氏鳇卵10粒) 1∶2 (4 cm体长环毛蚓5粒)和(达氏鳇卵10粒) 1∶1(4 cm体长环毛蚓10条)。实验期间,将实验鱼放入水族箱中适应24h后,由于瓦氏黄颡鱼是夜间摄食型鱼类,因此,在室外自然光下选择在16:00—18:00期间放置饵料进行实验。在次日8:00收拾残饵,并将每组实验鱼摄食饵料的数量百分比记为实验鱼的摄食率。
1.3 统计分析方法
记录数据为每次重复瓦氏黄颡鱼对每种饵料摄食的数量百分比,并统计3次重复摄食每种饵料的数量平均百分比,即瓦氏黄颡鱼的摄食率。数据均由SPSS11.5软件和Origin8.1进行统计处理,并均采用One-way ANOVA检验。统计值用平均值±标准值(mean±SD)表示,差异显著水平P<0.05,极显著水平P<0.01。
2 结果
2.1 3种规格瓦氏黄颡鱼对不同大小环毛蚓的摄食率
实验中仅大规格体长实验鱼摄食体长为8 cm的环毛蚓,而中小规格体长实验鱼均不摄食体长8 cm的环毛蚓(图1)。实验鱼对体长为4 cm和2 cm的环毛蚓的摄食率均不存在显著性差异 (P>0.05)(图1)。环毛蚓体长2 cm时,虽然实验鱼均摄食,但综合考虑2 cm环毛蚓体长规格获取的难易程度和瓦氏黄颡鱼达到饱和程度比4 cm环毛蚓体长规格觅食时间长,因此下一步实验确定选择体长4 cm的环毛蚓为干扰瓦氏黄颡鱼摄食达氏鳇卵的饵料。
2.2 3 种规格瓦氏黄颡鱼对达氏鳇卵的摄食率
在达氏鳇卵饱足投喂下,发现3种体长规格实验鱼均摄食达氏鳇卵,且3种体长实验鱼之间的摄食率不存在显著性差异(P>0.05) (图2)。研究结果表明,实验所选7 cm体长以上的瓦氏黄颡鱼均能够摄食达氏鳇卵,下一步可以选择达氏鳇卵作为中华鲟卵替代品开展实验。
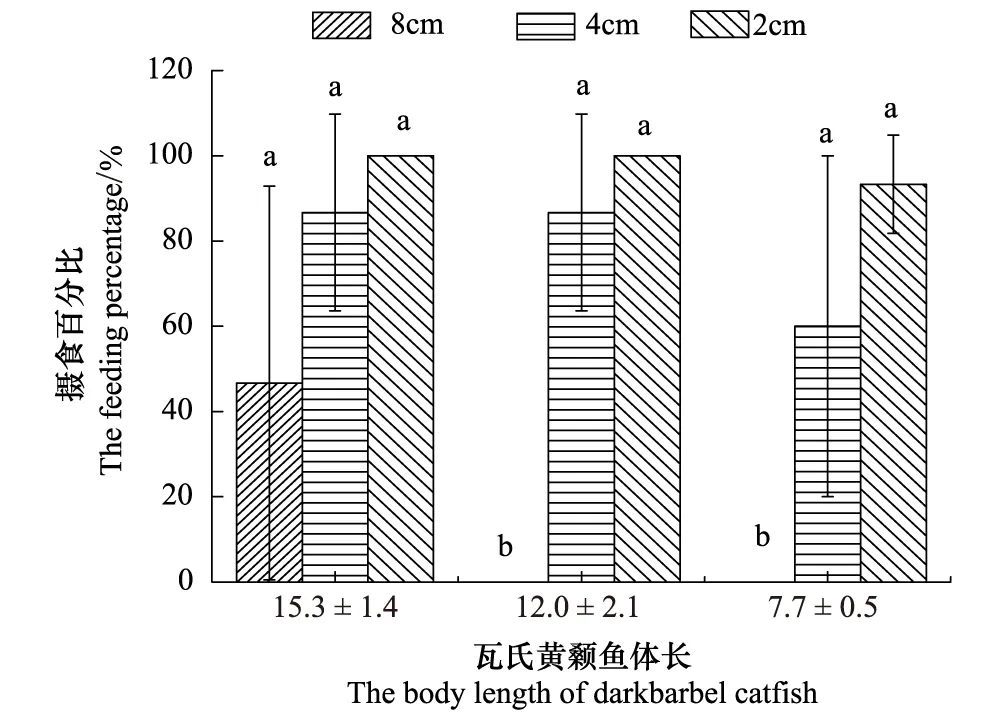
图1 瓦氏黄颡鱼对环毛蚓的摄食率Fig.1 The feeding rate on earthworm by darkbarl catfish图中柱上不同字母代表各体长组实验鱼对不同体长规格环毛蚓摄食率的差异显著性(P<0.05),相同字母代表差异不显著性(P>0.05)
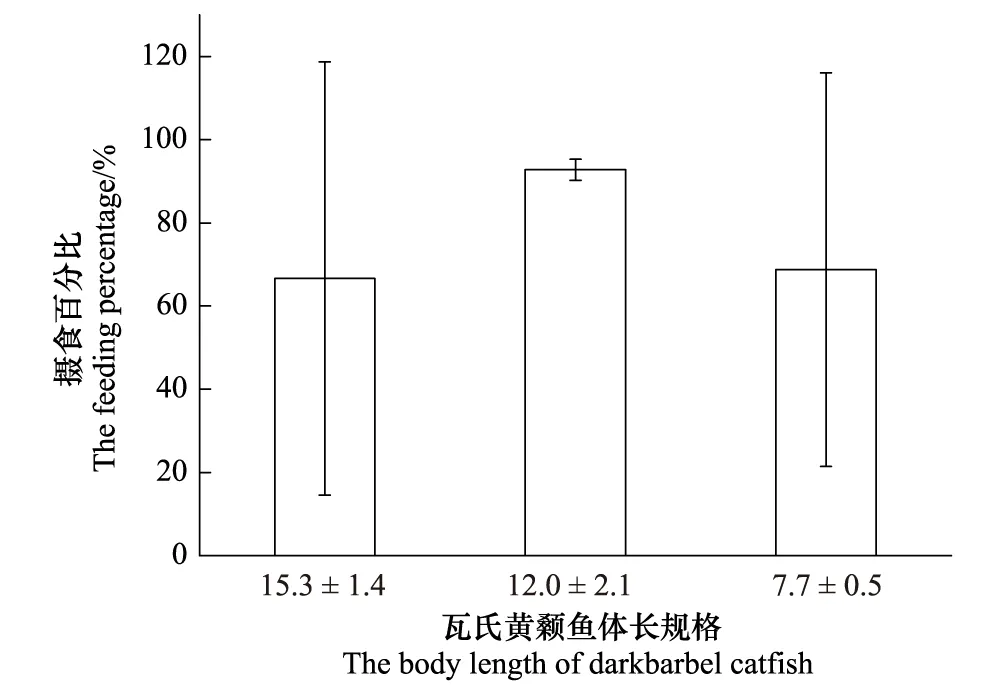
图2 瓦氏黄颡鱼对达氏鳇卵的摄食率Fig.2 The feeding rate on eggs of Kaluga by darkbarbel catfish
2.3 不同比例环毛蚓和达氏鳇卵对3种规格瓦氏黄颡鱼摄食偏好的影响
研究结果发现,达氏鳇卵和环毛蚓数量比例为1∶2时,瓦氏黄颡鱼对环毛蚓的摄食率高于达氏鳇卵,但3种体长瓦氏黄颡鱼的对环毛蚓的摄食率不显著高于达氏鳇卵(P>0.05)(图3)。当达氏鳇卵和环毛蚓数量比例为1∶1时,发现3种体长实验鱼对环毛蚓的摄食率显著高于达氏鳇卵,对达氏鳇卵的摄食率接近于0,且存在显著性差异[小规格体长实验鱼(P<0.05);中规格体长实验鱼(P<0.05);大规格体长实验鱼(P<0.05)](图4)。
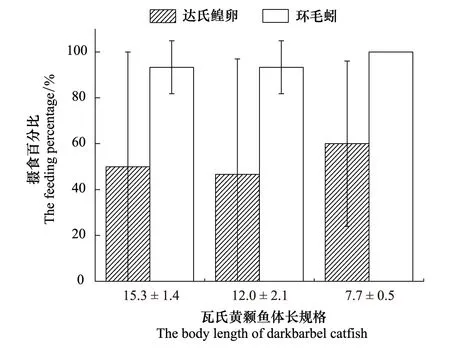
图3 环毛蚓和达氏鳇卵数量比为1∶2下瓦氏黄颡鱼的摄食率Fig.3 The feeding rate of darkbarbel catfish under 1∶2 of quantity ratio between Earthworms and eggs of Kaluga

图4 环毛蚓和达氏鳇卵数量比为1∶1下瓦氏黄颡鱼的摄食率Fig.4 The feeding rate of darkbarbel catfish under 1∶1 of quantity ratio between Earthworms and eggs of Kaluga
3 讨论
3.1 瓦氏黄颡鱼饵料的适口性
鱼类摄食的饵料适口与否主要取决于摄食饵料种类(饵料的营养成份如蛋白质、脂肪、维生素或消化难易程度)和摄食饵料规格(生物饵料的体长、体宽和人工饵料的颗粒体积等)[13- 16]。如在同样体长的鳙(Aristichthysnobilis)、鲮(Cirrhinusmolitorella)、鲢(Hypophthalmichthysmolitrix)、草鱼(Ctenopharyngodonidellus)、团头鲂(Megalobramaamblycephala)同时存在时,鳜鱼(Sinipercachuasti)优先摄食鳙和鲮,因为鳙和鲮的粗蛋白含量较其它几种饵料高[17];瓦氏黄颡鱼在多种饵料丰度相同条件下,偏好摄食蛋白含量较高的生物饵料如环毛蚓、河虾(Macrobranchiumnipponense)和麦穗鱼(Pseudorasboraparva)等[10]。本研究发现不同体长瓦氏黄颡鱼对达氏鳇卵摄食率很高,进一步表明瓦氏黄颡鱼喜欢摄食蛋白含量高的生物饵料[18]。
鱼类摄食既受饵料种类影响,又受到饵料规格限制。适宜规格的饵料可以减少摄取食物消耗的时间,提高食物中营养物质和能量的转化效率,缩短暴露于捕食者面前的时间,减少被捕食的风险,同时可以节省体能,提高鱼类成活率[19]。鱼类摄食何种规格的饵料主要受到口裂或口宽的限制,且不同体长或不同发育阶段鱼类的口裂或口宽存在差异,因此不同体长鱼类摄食饵料的规格存在差异[6,9,11,20- 23]。如随着鱼体全长的增加,大口黑鲈(Micropterussalmoides)摄食的饵料长度而逐步增加:大口黑鲈仔鱼(全长5.8—8.1 mm)摄食饵料长度为100—555 μm或55—200 μm,稚鱼(全长8.8—23.0 mm)摄食饵料长度为650—830 μm,幼鱼(全长23.6—30.3 mm)摄食饵料长度为1250—1350 mm[21]。鳜鱼的适口饵料规格也随着体长的增加而增大,其胃含物中大规格体长鱼类占有比例逐渐增多[9]。本研究中,随着瓦氏黄颡鱼体长的增加,其偏好摄食的环毛蚓体长规格也增加,佐证了鱼类摄食饵料规格范围随体长的变化而变化的观点。
3.2 环毛蚓对瓦氏黄颡鱼摄食达氏鳇卵的干扰效果
鱼类对饵料的摄食偏好除受到摄食饵料种类影响和饵料规格的限制外,还受到鱼类栖息地环境因素的影响,其主要环境因素之一为摄食饵料在栖息环境中存在的相对丰度[23- 26]。一般鱼类在多种饵料共存条件下选择丰度较高的饵料,这样可以减少搜寻和捕获饵料时的活动时间,减少被敌害鱼类捕食的风险,并获得更多净能量[19]。因此,鱼类在多种喜好饵料的密度存在差别时,鱼类是以饵料密度较高的喜好饵料为主。如轮虫和卤虫无节幼体皆适宜11日龄的牙鲆(Paralichthysolivaceus)仔鱼摄食,当两者密度存在差别时,牙鲆仔鱼对密度较高的饵料摄食量较大[27]。在白鲦(Hemiculterleucisculus)资源量丰富时,鳜鱼对白鲦的摄食量最大,而在麦穗鱼(Pseudorasboraparva)、鲫鱼(Carassiusauratus)、沙塘鳢(Odontobutisobscurus)等鱼类资源量丰富时,鳜鱼则对这些鱼类资源量摄食量较大[9]。在中华鲟产卵期间,中华鲟卵被食卵鱼类大量摄食,可能是由于中华鲟卵资源量在中华鲟产卵场相对其它饵料更为丰富的缘故。本研究选择4 cm体长环毛蚓来干扰实验鱼摄食达氏鳇卵,发现随着环毛蚓投喂的数量相对达氏鳇卵的比例增大,瓦氏黄颡鱼选择摄食环毛蚓的概率增大,当比例达到1∶1时,瓦氏黄颡鱼对达氏鳇卵存在极小的的摄食率,进一步证明增大鱼类某一偏好摄食饵料的密度可实现对其它偏好饵料的保护。
3.3 研究意义
本研究探讨了不同体长瓦氏黄颡鱼皆适宜摄食的环毛蚓体长规格,试验了干扰瓦氏黄颡鱼摄食达氏鳇卵的环毛蚓投放数量比例。本研究中实验对象瓦氏黄颡鱼是中华鲟卵敌害鱼类,达氏鳇卵在一定程度上模拟了中华鲟卵,因此实验结果具备为中华鲟卵的保护策略提供借鉴的参考价值,为下一步实施野外中华鲟卵保护实验提供了科学依据。研究发现在达氏鳇卵单独存在条件下,7 cm体长以上的瓦氏黄颡鱼均可以摄食达氏鳇卵,推断长江葛洲坝下游中华鲟产卵场内体长7 cm以上的瓦氏黄颡鱼均存在摄食中华鲟卵的可能性,但何种规格危害最大有待进一步探讨。本研究发现4 cm体长环毛蚓与达氏鳇卵数量比例为1∶1时,瓦氏黄颡鱼几乎不摄食达氏鳇卵,表明4 cm体长环毛蚓可能作为鲟鱼卵的替代品实现鲟鱼卵如中华鲟卵的保护。本研究提示在中华鲟产卵时期,通过向中华鲟产卵场投喂替代饵料如4cm体长环毛蚓并提高其相对中华鲟卵的数量百分比,有望实现对中华鲟卵的保护,但有待野外实验验证。此外,本研究对瓦氏黄颡鱼饵料的适口性进行了探讨,发现瓦氏黄颡鱼随着体长的增加摄食环毛蚓的体长规格适宜范围也增加,该研究结果可以为瓦氏黄颡鱼的人工养殖技术改进提供依据,养殖者可以根据瓦氏黄颡鱼的生长时期提供相适宜规格的饲料以提高瓦氏黄颡鱼的生产效率。
[1] 虞功亮, 刘军, 许蕴玕, 常剑波. 葛洲坝下游江段中华鲟产卵场食卵鱼类资源量估算. 水生生物学报, 2002, 26(6): 591- 599.
[2] 罗佳, 姜伟, 陈求稳, 刘德富, 唐锡良, 涂志英, 石小涛. 葛洲坝下中华鲟产卵场食卵鱼资源量的调查和分析. 淡水渔业, 2013, 43(5): 28- 31.
[3] 李明锋. 瓦氏黄颡鱼研究进展及前景展望. 现代渔业信息, 2011, 26(1): 5- 12.
[4] 李芹, 刁晓明. 不同饵料对瓦氏黄颡鱼稚鱼生长和消化酶活性的影响. 水生态学杂志, 2009, 2(1): 98- 102.
[5] 王武, 李伟纯, 马旭洲, 何丽, 叶斐菲. 水温与光照对瓦氏黄颡鱼幼鱼行为的影响. 生态学杂志, 2008, 27(5): 791- 796.
[6] 周德勇, 叶佳林, 王卫民, 刘正文. 太湖梅梁湾红鳍原鲌(Cultrichthys erythropterus)食性与个体大小的关系: 对生物调控与管理的启示. 湖泊科学, 2011, 23(5): 796- 800.
[7] 贺舟挺, 薛利建, 金海卫. 东海北部近海棘头梅童鱼食性及营养级的探讨. 海洋渔业, 2011, 33(3): 265- 273.
[8] 尚玉昌. 行为生态学. 北京: 北京大学出版社, 1998.
[9] Li W. Zhang T, Ye S, Liu J, Li Z. Feeding habits and predator-prey size relationships of mandarin fish Siniperca chuatsi (Basilewsky) in a shallow lake, central China. Journal of Applied Ichthyology, 2013, 29(1): 56- 63.
[10] 路波, 白艳勤, 罗佳, 刘德富, 石小涛, 陈求稳, 黄应平. 葛洲坝下游中华鲟产卵场瓦氏黄颡鱼摄食活动的试验研究. 生态学杂志, 2013, 32(10): 2728- 2734.
[11] Alp A, Yegen V, Apaydin Y M, Uysal R, Bicen E, Yagci A. Diet composition and prey selection of the pike, Esoxlucius, in Civril Lake, Turkey. Journal of Applied Ichthyology, 2008, 24(6): 670- 677.
[12] 季强. 异齿裂腹鱼食性的初步研究. 水利渔业, 2008, 28(3): 51- 53.
[13] 韩志泉, 郭雪清, 田雨, 孙晓雯. 淡水鱼所需五种动物性饵料脂肪酸组成比较. 首都师范大学学报: 自然科学版, 2001, 22(1): 73- 79.
[14] Tonn W M, Holopainen I J, Paszkowski C A. Piscivory and recruitment: mechanisms structuring prey populations in small lakes. Ecology, 1992, 73(3): 951- 958.
[15] Claessen D, Van Oss C, de Roos A M, Persson L. The impact of size-dependent predation on population dynamics and individual life history. Ecology, 2002, 83(6): 1660- 1675.
[16] Wysujack K, Mehner T. Can feeding of European catfish prevent cyprinids from reaching a size refuge. Ecology of Freshwater Fish, 2005, 14(1): 87- 95.
[17] 梁银铨, 刘友亮, 崔希群. 鳜对饵料鱼类选择性的营养生理分析. 水利渔业, 1995, (1): 9- 11.
[18] 高露姣, 夏永涛, 黄艳青, 师伟, 庄平, 张涛, 宋超. 俄罗斯鲟鱼卵与西伯利亚鲟鱼卵的营养成分比较. 海洋渔业, 2012, 34(1): 57- 63.
[19] 石小涛, 陈求稳, 庄平, 刘德富, 黄应平. 提高摄食-反捕食能力导向的鱼类野化训练方法述评. 生态学杂志, 2012, 31(12): 3235- 3240.
[20] Lazzaro X. A review of planktivorous fishes: Their evolution, feeding behaviours, selectivities, and impacts. Hydrobiologia, 1987, 146(2): 97- 167.
[21] 张韵桐, 夏金华, 杨正华. 大口黑鲈仔幼鱼的饵料选择和生长的研究. 湛江水产学院学报, 1992, 12(1): 19- 24.
[22] 张波, 孙耀, 郭学武, 王俊, 唐启升. 黑鲪的最大摄食率与温度和体重的关系. 海洋水产研究, 1999, 20(2): 82- 85.
[23] 童玉和, 郭学武. 两种岩礁鱼类的食物竞争实验. 中国水产科学, 2009, 16(4): 541- 549.
[24] 杨纪明. 渤海鱼类的食性和营养级研究. 现代渔业信息, 2001, 16(10): 10- 19.
[25] 郭学武, 唐启升. 鱼类摄食量的研究方法. 海洋水产研究, 2004, 25(1): 68- 78.
[26] Dorner H, Hulsmann S, Holker F, Skov C, Wagner A. Size-dependent predator-prey relationships between pikeperch and their prey fish. Ecology of Freshwater Fish, 2007, 16(3): 307- 314.
[27] 龚小玲, 鲍宝龙, 苏锦祥. 牙鲆仔鱼在混合饵料期的摄食能力及饵料选择性. 上海水产大学学报, 2004, 13(1): 30- 35.
Earthworms can be used to deter predation on sturgeon eggs by darkbarbel catfish
SHI Xiaotao1,2,3, LU Bo1, BAI Yanqin1, HU Yunshen1, LUO Jia1, CHEN Qiuwen1,3, QIAO Ye2, LIU Defu1,*
1EngineeringResearchCenterofEco-environmentinThreeGorgesReservoirRegion,MinistryofEducation,ChinaThreeGorgesUniversity,Yichang443002,China2KeyLaboratoryofEcologicalImpactsofHydraulic-projectsandRestorationofAquaticEcosystem,MinistryofWaterResources,Wuhan430079,China3ChinaResearchCenterforEco-EnvironmentalSciences,ChineseAcademyofSciences,Beijing100085,China
The eggs of Chinese sturgeon (Acipensersinensis), an important native fish species in China, are extremely vulnerable to predation by other fishes in the wild. In some cases, more than 90% of Chinese sturgeon eggs are consumed by predators. Darkbarbel catfish [Pelteobagrusvachelli(Richerdson)] are major predators of Chinese sturgeon eggs. Thus, it is critical to gain a better understanding of the feeding behaviors of Darkbarbel catfish to develop practical ways to protect Chinese sturgeon eggs from predation. In this study, we explore the use of bait as a potential tool to redirect Darkbarbel catfish′s feeding away from Chinese sturgeon eggs. In a laboratory experiment, Darkbarbel catfish were fed earthworms (Pheretimatschiliensis) of different sizes, and we observed their feeding habits and preference between earthworms and Kaluga sturgeon eggs. Darkbarbel catfish were captured from a known Chinese sturgeon spawning area at the lower reaches of Gezhou Dam, and fed in the experimental tanks for 30 days to adapt to the experimental environment before beginning feeding trials. The experimental environment was supplied with water at a constant (20.0 ± 1.5) ℃ temperature under natural light conditions. Darkbarbel catfish were divided into 3 groups according to standard body length[ (15.3 ± 1.4) cm, (12.0 ± 2.1) cm, (7.7 ± 0.5) cm] and fed a diet of either different-sized earthworms (body height of 0.3 cm; body length of 8 cm, 4 cm or 2 cm, respectively) or Kaluga sturgeon eggs. The results showed that earthworms of body length of 8 cm can only be eaten by fish in the largest size, whereas earthworms 2 cm and 4 cm in length can be eaten by fish of all sizes. Kaluga sturgeon eggs were eaten by Darkbarbel catfish of all sizes. There were no significant differences between the feeding rates of Darkbarbel catfish on earthworms 2 or 4 cm in length. We also examined whether medium-sized earthworms (body length of 4 cm) could be used to distract Darkbarbel catfish when feeding on Kaluga sturgeon eggs. Darkbarbel catfish were simultaneously offered Kaluga sturgeon eggs and earthworms in different quantities. The results showed that the liklihood of Darkbarbel catfish eating Kaluga sturgeon eggs was close to 0 when the ratio of the quantity of earthworms to Kaluga sturgeon eggs is 1∶1, while half of the Kaluga sturgeon eggs were eaten when the ratio of Kaluga sturgeon eggs to earthworms is 2∶1. The findings indicated that 1) Darkbarbel catfish of all size groups will eat Kaluga sturgeon eggs and earthworms; 2) earthworms with body length equal to or shorter than 4 cm could be appropriate as bait for Darkbarbel catfish of all sizes; 3) the ratio of earthworms to Kaluga sturgeon eggs is an important determinant in the feeding preference of Darkbarbel catfish; 4) earthworms could potentially be used as a bait to feed Darkbarbel catfish to mitigate predations on sturgeon eggs; 5) Darkbarbel catfish prefer to eat earthworms rather than Kaluga sturgeon eggs when the ratio of earthworms to eggs reaches or exceeds 1∶1. Considering the similarity between Chinese sturgeon and Kaluga sturgeon eggs, this study could help inform conservation strategies for the Chinese sturgeon in Yangtze River. Further verifications in the field and on Chinese sturgeon eggs are now needed.
darkbarbel catfish; earthworms; kaluga sturgeon eggs; feeding
楚天学者基金(KJ2010B002); 国家自然科学基金(51009082, 51210105017); 水利部公益性行业科研专项(201201030); 水利部水工程生态效应与生态修复重点实验室开放基金; 三峡库区生态环境教育部工程研究中心开放基金(KF2013- 03)
2013- 09- 14;
日期:2014- 07- 14
10.5846/stxb201309142275
*通讯作者Corresponding author.E-mail: fishlab@163. com
石小涛, 路波, 白艳勤, 胡运燊, 罗佳, 陈求稳 乔晔, 刘德富.蚯蚓对瓦氏黄颡鱼摄食达氏鳇卵的干扰实验.生态学报,2015,35(4):1045- 1050.
Shi X T, Lu B, Bai Y Q, Hu Y S, Luo J, Chen Q W, Qiao Y, Liu D F.Earthworms can be used to deter predation on sturgeon eggs by darkbarbel catfish.Acta Ecologica Sinica,2015,35(4):1045- 1050.
