南沙群岛美济礁海域夏季浮游动物群落特征
2015-03-10杜飞雁王雪辉林昭进
杜飞雁,王雪辉,林昭进
1 中国水产科学研究院南海水产研究所,广州 510300 2 广东省渔业生态环境重点开放实验室,广州 510300 3 农业部南海渔业资源环境科学观测实验站,广州 510300
南沙群岛美济礁海域夏季浮游动物群落特征
杜飞雁1,2,3,*,王雪辉1,2,林昭进1,2
1 中国水产科学研究院南海水产研究所,广州 510300 2 广东省渔业生态环境重点开放实验室,广州 510300 3 农业部南海渔业资源环境科学观测实验站,广州 510300
浮游动物是珊瑚礁生态系统中的重要组成部分,但国内相关研究相对较少。利用2012年7月在南沙群岛美济礁海域开展的13个站次的海洋生物调查数据,对美济礁浮游动物的群落特征进行研究,表明:(1) 美济礁浮游动物种类组成丰富,共出现15个类群138种(类)浮游动物; (2) 优势种组成复杂、区域变化明显,单一种的优势度不高; (3) 浮游动物平均密度和湿重生物量分别为117.70 个/m3、69.01 mg/m3,浮游幼虫在总密度中所占比例最大; (4) 浮游动物非常丰富,多样性程度较高。平均丰富度、多样性指数和均匀度分别为4.93、3.33和0.67;(5) 浮游动物可划分为2个群落,分别为潟湖-礁坪区群落和向海坡群落;(6) 受珊瑚礁不同生物地貌带的空间异质性和水动力条件的影响,美济礁浮游动物群落区域差异明显。潟湖区浮游动物多样性较高,生物量最低;礁坪区浮游动物数量大,但分布不均匀,且多样性水平最低;向海坡区浮游动物种类最多、多样性非常丰富,但栖息密度最低。
南沙群岛;美济礁; 浮游动物;群落特征
分布于热带海洋的珊瑚礁具有极丰富的生物多样性和生态服务功能,是最重要的生态系统之一。浮游动物在珊瑚礁生态系统的物质循环和能量流动过程中起重要作用,是珊瑚礁吸收外来营养源的中间环节,摄食浮游动物和大的颗粒有机物是许多礁栖动物,包括鱼类、海葵、海百合、腹足类以及珊瑚虫的主要异养性营养路径。开展浮游动物研究对探索热带珊瑚礁生态系统的结构与功能具有十分重要的意义[1]。国内有关珊瑚礁浮游动物研究相对较少,仅有陈清潮、张谷贤、章淑珍、尹健强和柯志新等,对中沙群岛黄岩岛环礁、南沙群岛9个环礁潟湖、渚碧礁、雷州半岛灯楼角和三亚珊瑚礁浮游动物的种类组成、昼夜变化、数量等群落特征进行了研究[1- 7]。
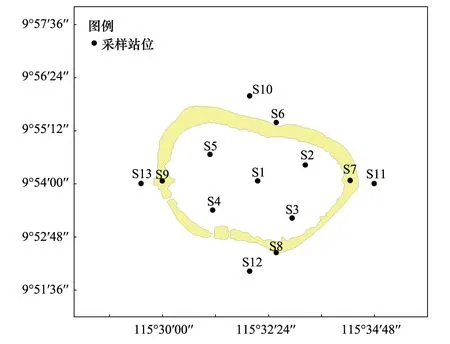
图1 调查站位设置示意图Fig.1 Sketch map of sampling stations
美济礁(9°52′—9°56′N,115°30′—115°35′E)为椭圆形准封闭型环礁,有口门存在,潟湖与礁外深海之间能进行海水交换。美济礁东西长约9 km,南北宽约6 km,礁顶面积45.31 km2、礁坪14.69 km2,潟湖30.62 km2,潟湖最大水深约27 m。美济礁浮游动物仅在南沙群岛的大面调查中有所涉及,但专题研究目前尚未见报道。美济礁具有礁前向海坡、礁坪和潟湖典型的生物地貌分带[8],而3个生物地貌分带区的海洋生物群落特征目前尚不清楚。本文利用2012年7月美济礁海域的调查数据,对潟湖、礁坪和向海坡3个区域浮游动物的群落特征进行较为深入的研究,以期有助于对热带珊瑚礁生态系统结构的全面认识。
1 材料与方法
1.1 站位设置及取样方法
2012年7月14日在南沙美济礁海域设置13个测站对浮游动物进行了采样,站位分布见图1,各调查站位地理坐标见表1。使用浅水I型浮游生物网(网长145 cm,网口内径50 cm,网口面积0.2 m2,网目大小0.505 mm)采样,每站自底层至表层垂直拖曳1网。样品的处理、保存、计数等均按《海洋调查规范》[9]进行。温度和盐度使用YSI 556进行现场测定,现场采水后,经0.45 um孔径醋酸纤维滤膜过滤、丙酮萃取后使用Turner-Design 10型荧光计测定叶绿素a含量。
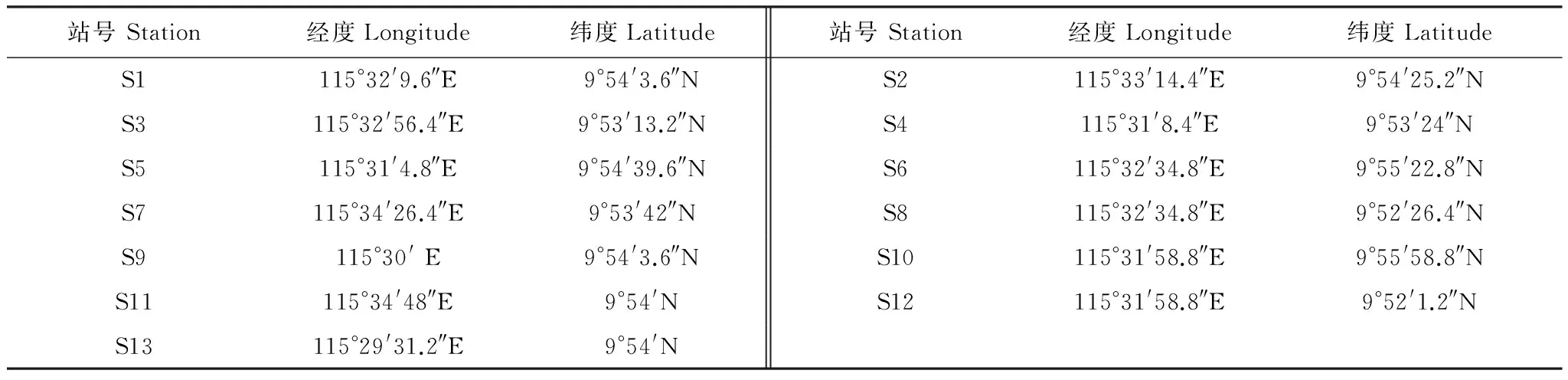
表1 调查站位地理坐标Table 1 Geographical positions of the sampling stations
1.2 数据统计及分析
采用优势度计算浮游动物优势种[10],其计算式为:
Y=(ni/N) ×fi
式中,ni为第i种的个体数量;N为某站总浮游动物个体数;fi为某种生物的出现频率。选用种类更替率(R)进行种类组成的区域变化研究[11],其计算式为:
R=[(a+b-2c)/(a+b-c)]×100%
式中,a与b分别为相邻2个区域的种数,c为相邻2个区域共同的种数。
采用Margalef种类丰富度指数D、Shannon-Wiener 指数H′、Pielou均匀度J和多样性阈值(Dv)来研究浮游动物多样性。Margalef种类丰富度指数D计算式为:
D=(S-1)/lnN;
Shannon-Wiener指数:
Pielou均匀度:
J=H′/log2S;
式中,S为样品中的种类总数,Pi为第i种的个体数(ni)与总个体数(N)的比值(ni/N);Dv=H′×J。
浮游动物群落结构的多变量分析通过PRIMER6.0软件,采用非参数多变量群落结构分析方法,为减少机会种对群落结构的干扰,首先根据丰度四次方根转换计算的Bray-Curtis相似性矩阵,应用组间平均聚类(Group average cluster)分析和非度量多维标度排序MDS(Non-metri multi-dimensional scaling)分析来研究群落结构[12- 13]。
通过SPSS18软件完成浮游动物群落与环境因子的双变量Spearman相关性分析,通过Suffer 8.0 完成图件绘制。
2 结果与分析
2.1 种类组成和生态类型
研究区域内共出现浮游动物138种(类),分属于腔肠动物水螅水母类、管水母类、栉水母类、枝角类、桡足类、端足类、磷虾类、十足类、等足类、介形类、翼足类、毛颚类、有尾类、海樽类和浮游幼虫等15个类群。其中以桡足类种类数最多,达42种。潟湖区出现浮游动物68种,礁坪区37种,向海坡出现种类最多,有101种。从各区域种类组成的相似性来看,潟湖和礁坪区的种类相似性为31.2%,潟湖与向海坡的相似性为29%,礁坪区与向海坡的相似性为20%,3个区域的共有种仅有19种。表明,美济礁浮游动物种类组成有明显的区域差异。
美济礁远离海岸,位于热带的高温高盐海域。调查期间海水维持高温高盐状态,水温在28.62—31.86 ℃之间,平均29.47 ℃,盐度变化范围为34.03—34.23,平均34.13。浮游动物生态类型呈明显的暖水性,大致有大洋性暖水种、沿岸暖水种和暖水广布种3种类型。大洋性暖水种主要有爪室水母Chelophyesappendiculata、小哲水蚤Nannocalanusminor、普通波水蚤Undinulavulgaris、达氏筛哲水蚤Cosmocalanusdarwinii、奥氏胸刺水蚤Centropagesorsinii、中隆剑水蚤Oncaeamedia和微型箭虫Sagittaminima等。沿岸暖水种有嵴状镰螅水母Zancleacostata、双生水母Diphyeschamissonis、拟细浅室水母Lensiasubtiloides、球型侧腕水母Pleurobrachiaglobosa、肥胖三角溞Evadnetergestina、异尾宽水蚤Temoradiscaudata、汤氏长足水蚤Calanopiathompsoni、小纺锤水蚤Acartianegligens、弱箭虫Sagittadelicata和长尾住囊虫Oikopleuralongicauda等。暖水广布种有四叶小舌水母Liriopetetraphylla、针刺真浮萤Euconchoeciaaculeata和肥胖箭虫Sagittaenflata等[1,14- 15]。此外,还出现了正型莹虾Lucifertypus和角锚真哲水蚤Rhincalanuscornutus等暖流指示种,反映出美济礁受外海暖流和沿岸水的共同影响。
2.2 优势种
优势种是具有控制群落和反映群落特征的种类。以优势度Y≥0.02为划分标准,研究区域内浮游动物优势种由鱼卵、长尾住囊虫、长尾类幼体、奥氏胸刺水蚤、住囊虫、粗壮箭虫Sagittarobusta、短尾类溞状幼体和单胃住筒虫Fritillariahaplostoma等8种组成。优势种的生态类型以大洋性暖水种和沿岸暖水种为主,浮游幼虫的优势较为明显。除鱼卵和长尾住囊虫的优势度分别为0.27和0.12,高于0.10外,其它均低于0.10。美济礁浮游动物优势种组成较为复杂,且单一种的优势度不高,与热带海区的生物群落特征相符合[16]。
美济礁浮游动物优势种组成有明显的区域差异,潟湖和礁坪区的优势种均有8种,虽然第一优势种有所不同,分别为长尾住囊虫和鱼卵,但优势种组成的相似性高达77.8%。而向海坡区的优势种组成与潟湖和礁坪区有明显差异,主要表现在以下3个方面:(1) 向海坡区优势种组成明显较为复杂,有羽长腹剑水蚤Oithonaplumifera、双尾纽鳃樽Thaliademocratica、住囊虫和长尾类幼体等15种组成。(2) 向海坡优势种组成与潟湖和礁坪区的组成差异较大,与潟湖区和礁坪区优势种的相似性分别只有27.8%和21%。(3) 3个区域中以向海坡单一种的优势度最低,各优势种的优势度最为接近,全部优势种的优势度均低于0.08(表2)。
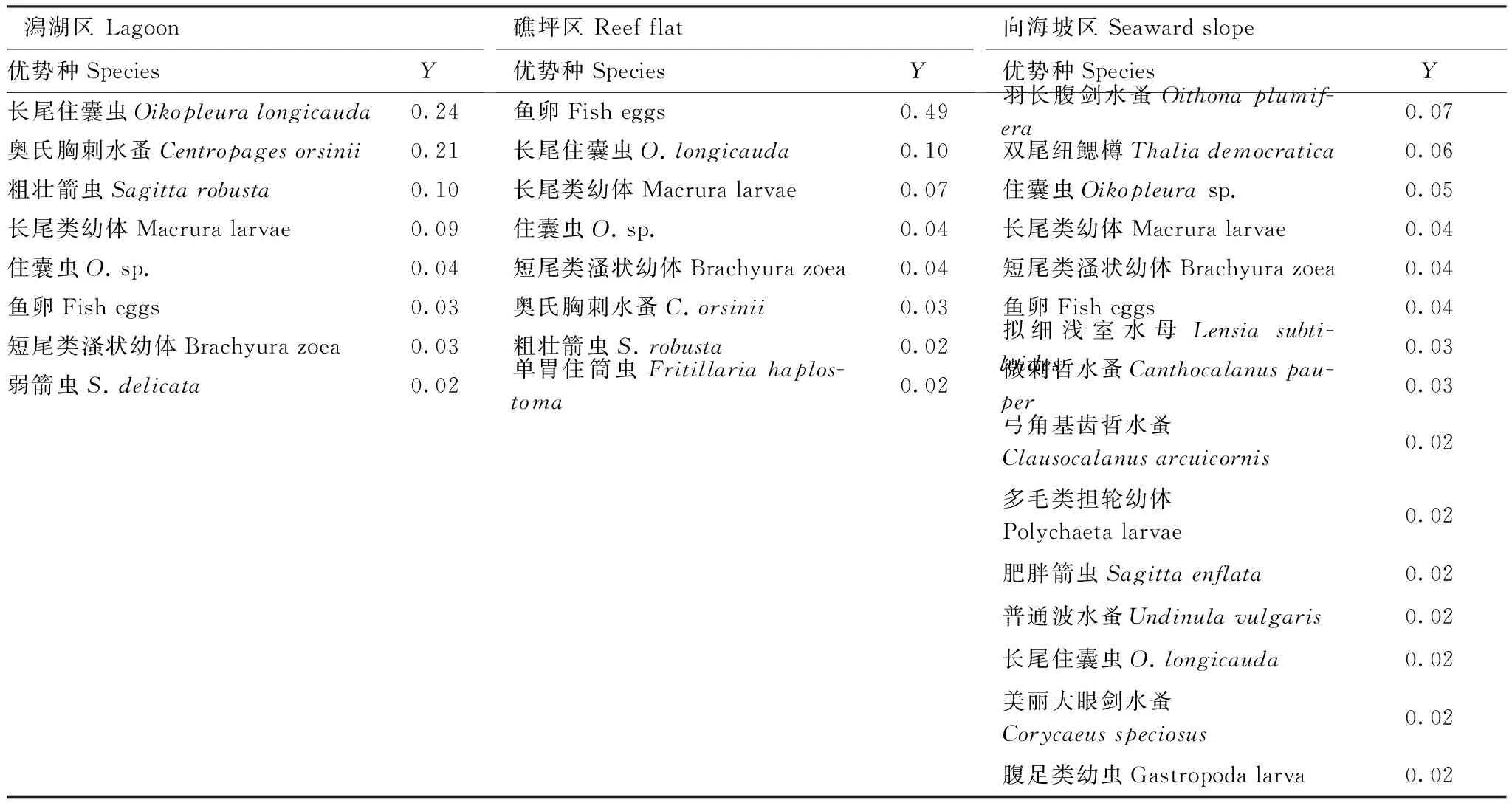
表2 美济礁不同区域浮游动物优势种组成Table 2 Composition of the dominant zooplankton in Meiji coral reef in summer
2.3 密度与生物量
美济礁海域夏季浮游动物平均密度为117.70 个/m3、平均湿重生物量为69.01 mg/m3。礁坪区密度和生物量分别为198.29 个/m3和126.05 mg/m3,潟湖区为100.50 个/m3、38.20 mg/m3,向海坡为58.63 个/m3和50.49 mg/m3。如图2和图3所示,浮游动物平面分布不均匀,礁坪区的密度和生物量明显高于向海坡和潟湖区,是美济礁海域浮游动物高数量区。密度以向海坡最低,生物量则以潟湖区最低。
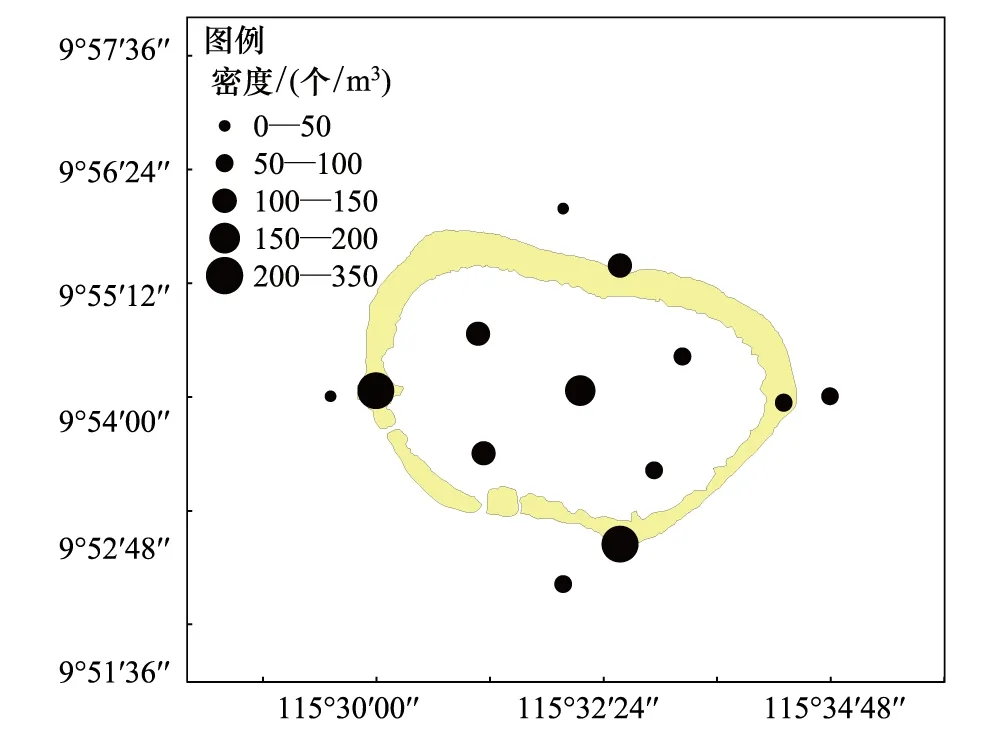
图2 美济礁夏季浮游动物密度平面分布Fig.2 Horizontal distribution of zooplankton abundance
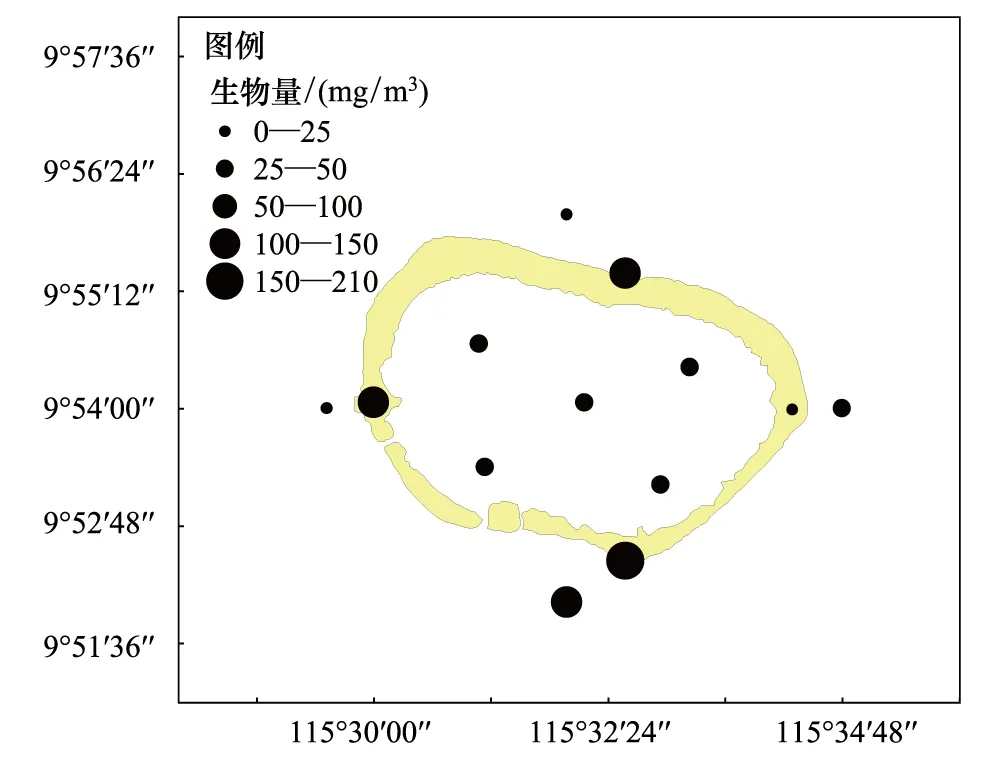
图3 美济礁夏季浮游动物湿重生物量平面分布Fig.3 Horizontal distribution of zooplankton biomass
从浮游动物的数量组成来看,美济礁浮游动物以浮游幼虫为主,其密度为52.32 个/m3,占总数量的44.5%,主要分布于礁坪区;有尾类次之,密度为28.02,占23.8%,也主要分布于礁坪区;桡足类密度为22.07 个/m3,占18.8%,居第三位,主要分布于潟湖区;其余13个类群的数量较低,累积占总量的12.9%。
潟湖区、礁坪区和向海坡区浮游动物数量的类群组成有明显的差异。潟湖区浮游动物密度仅次于礁坪区,但出现类群数最低,有8个类群。各类群中,以桡足类为主,其平均数量为33.25 个/m3,占潟湖区总量的33.1%;此外,有尾类、浮游幼虫和毛颚类的数量也较高。礁坪区出现12个类群,其中磷虾类和等足类仅在礁坪区出现。各类群中,以浮游幼虫数量占绝对优势,其密度高达128.48 个/m3,占64.8%;有尾类次之,占23.1%。向海坡类群组成最为复杂,除磷虾类、等足类和介形类外,其余13个类群均有出现。其中,枝角类、十足类、翼足类和多毛类只在向海坡出现。各类群中,仍以浮游幼虫为主,其密度为17.38 个/m3,占29.8%;此外,桡足类和有尾类数量也较高,分别占总量的28.8%的11.1%。
潟湖区浮游动物湿重生物量在26.95—45.50 mg/m3之间变化,平面分布的差异较小(SD=9.97);向海坡区生物量在17.13—115.95 mg/m3之间变化,除南部的S12站生物量达到115.95 mg/m3外,其余各站数量均低于50 mg/m3,平面分布的差异较大(SD=45.78);礁坪区是高生物量区所在,北、西和南侧3个测站的生物量均超过100 mg/m3,南侧S10站的生物量高达209.83 mg/m3。而东侧S7站的生物量只有24.68 mg/m3,密度也在礁坪区最低,为79.00 个/m3。礁坪是珊瑚虫及礁栖动物的主要栖息场所,据报道渚碧礁礁坪区浮游动物的数量由于捕食作用,明显低于潟湖区[1]。据在美济礁的现场观测和实地走访表明,美济礁东侧的珊瑚生长和生态环境状况明显优于其它区域。珊瑚虫及礁栖动物的捕食作用,可能是礁坪东侧浮游动物数量明显偏低的直接原因。
2.4 多样性特征
美济礁海域浮游动物种类非常丰富,平均丰富度为4.93,S11站高达10.05。其中向海坡区丰富度高达8.72,礁坪区丰富度最低,为2.76;潟湖区丰富度为3.62。多样性指数均值为3.33,仍以S11站最高,向海坡(4.79)> 潟湖区(2.99) >礁坪区(2.31)。均匀度均值为0.67,以S12站最高,向海坡(0.84)>潟湖区(0.65)>礁坪区(0.53)(表3)。
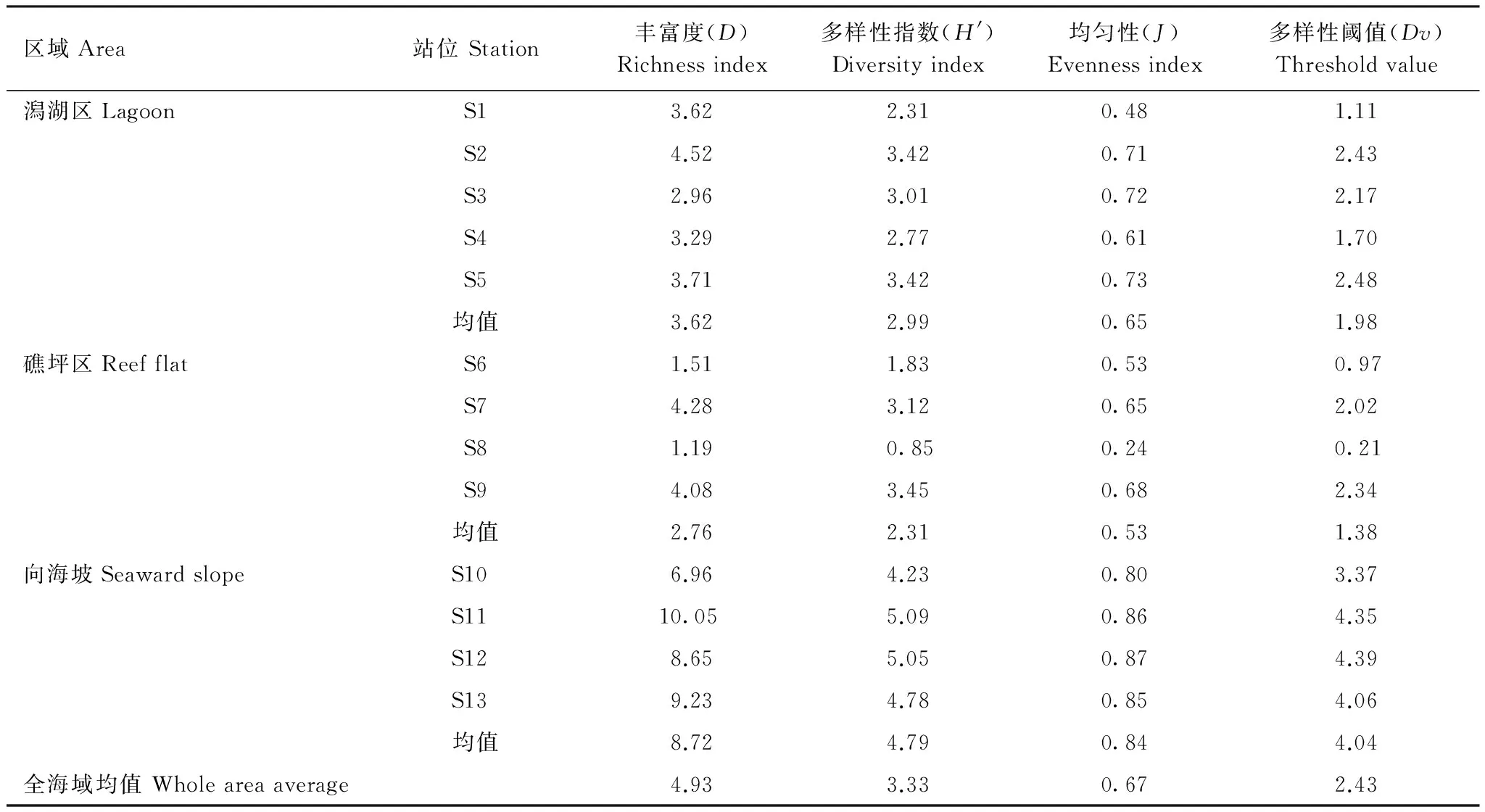
表3 美济礁浮游动物多样性指数、均匀度和丰富度Table 3 Indices of diversity,evenness and richness of zooplankton in Meiji coral reef
根据陈清潮等[10]给出的热带海区生物多样性阈值,对美济礁浮游动物多样性水平进行评价。全海域平均多样性阈值为2.43,浮游动物多样性较好水平。其中,向海坡多样性阈值为4.04,多样性非常丰富;潟湖区为1.98,多样性水平较好;而礁坪区为1.38,多样性程度一般。
2.5 群落格局
聚类分析和MDS分析表明,夏季南沙美济礁海域浮游动物可划分为2个群落。潟湖和礁坪区的S1、S2、S3、S4、S5、S6、S7、S8和S9站组成群落Ⅰ,向海坡的S10、S11、S12和S13站形成群落Ⅱ。MDS分析的协强系数stress=0.07,表明该结果较好地反映了样本间的相似关系(图4)。One-way ANOSIM检验表明,各群落间的差异极为显著(R=0.865,P=0.001)。
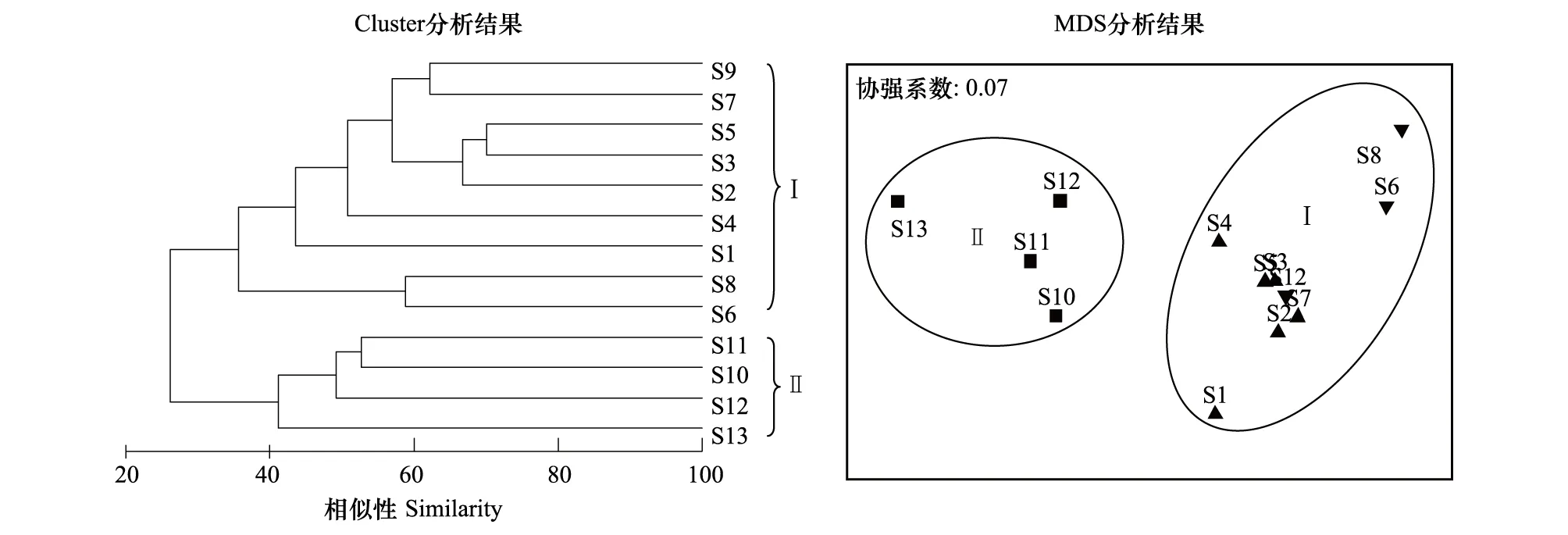
图4 南沙美济礁浮游动物群落结构的聚类分析和序列分析Fig.4 Cluster and MDS ordination of zooplankton in Meiji coral reef, Nansha Islands
从聚类分析和MDS分析结果可以看出,向海坡与礁坪区和潟湖区浮游动物的组成存在明显的差异,独自形成向海坡群落。而礁坪区的浮游动物虽然与潟湖区有一定的差异,但仍有较高的相似性,共同组成潟湖和礁坪群落。
2.6 与环境因子的关系
将浮游动物丰富度、多样性指数、均匀度、密度和生物量与现场同步实测的海水水温、盐度、pH、DO、悬浮物和叶绿素a数据,进行相关分析,结果见表4。
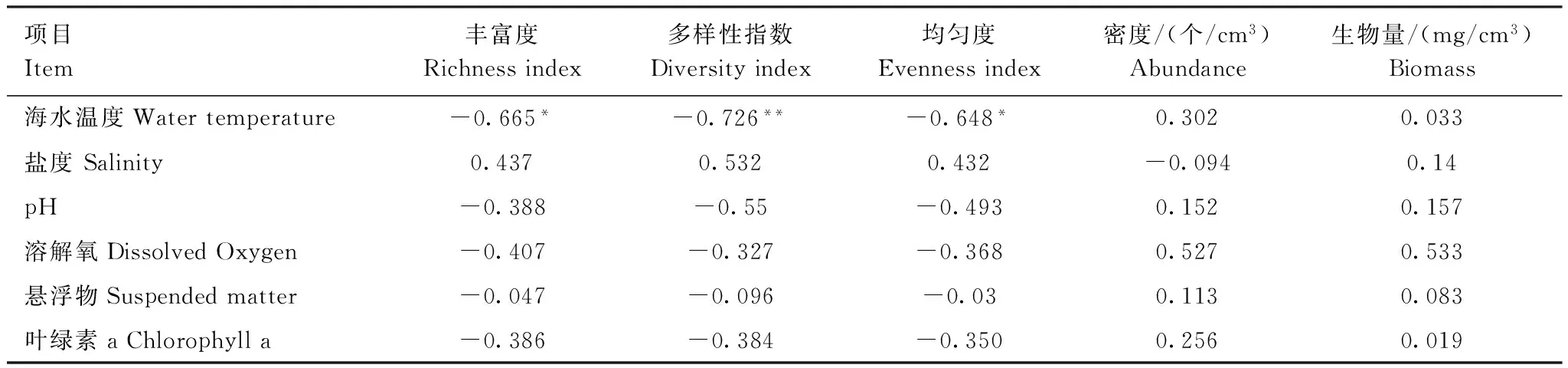
表4 浮游动物与环境因子相关系数表Table 4 Correlation coefficient of environmental factors to zooplankton
** 在置信度(双测)为 0.01 时,相关性是显著的;* 在置信度(双测)为 0.05 时,相关性是显著的
相关分析结果表明,各环境因子中除海水温度外,其它各因子与浮游动物的相关性不显著。海水温度与浮游动物丰富度、多样性指数呈非常显著的负相关,与丰富度和均匀度呈显著的负相关。美济礁向海坡区浮游动物的丰富度、多样性指数和均匀度最高,而此区域的海水温度为29.01,明显低于潟湖区的29.55和礁坪区的30.23。海水温度最高的礁坪区浮游动物多样性指数和均匀度均最低。
3 讨论
3.1 浮游幼虫在群落中占重要地位
珊瑚礁复杂多样的生境栖息着种类繁多、生活方式多样的礁栖无脊椎动物和脊椎动物,这些动物的幼虫成为珊瑚礁浮游动物群落的重要组成部分[5]。浮游幼虫在珊瑚礁浮游动物群落中占有重要地位,也是珊瑚礁浮游动物群落的特征之一[17- 18]。美济礁浮游幼虫具有种类、数量大和优势地位突出的特点,在浮游动物群落中占重要地位。本次调查共出现浮游动物138种,其中浮游幼虫出现24种,数量仅次于桡足类居第二位。研究区内37.5%的优势种为浮游幼虫,分别为鱼卵、长尾类幼体和短尾类溞状幼体。其中,鱼卵为美济礁海域浮游动物的第一优势种。浮游幼虫的密度居各类群的首位,占总数量的44.5%,主要分布于礁坪区和向海坡海域。
3.2 多样性水平
美济礁礁盘外的礁前向海坡区域浮游动物的丰富度、多样性指数和均匀度均明显高于潟湖区。渚碧礁潟湖区浮游动物多样性指数也明显低于礁坪区和岛礁外开阔海区,浮游植物多样性指数也低于外部开阔海域[1,5,19]。表明南沙海域的珊瑚礁潟湖可能不是珊瑚礁区生物多样性水平最高的区域。由于珊瑚礁区域海洋生物研究资料相对较少,这方面的研究还有待进一步深入。
3.3 浮游动物的区域差异明显
南沙群岛珊瑚礁的地貌形态受海洋水动力因素所控制,其中波浪是最主要的动力因素。波浪自礁坪外缘向潟湖方向传播,越过整个礁坪时能量逐渐衰减,形成波能分布带。不同波能带内水动力条件不同,适宜不同的珊瑚生长,发育成不同形态的动力地貌单元[20]。美济礁具有礁前向海坡、礁坪和潟湖典型的生物地貌分带。向海坡区和外礁坪区是波能的主要损耗带,水动力最强,是波浪的破碎区;礁坪区属于中波能带,水动力适宜,是珊瑚的主要生长带;潟湖区属低波能带,水动力最弱、风浪最小,由于礁坪的阻挡,波浪的影响大为削弱。此外,浮游生物与海流的依存关系密切,受水动力条件的影响明显。地貌和水动力条件的差异,也造成了其间海洋生物群落的差异。在特定的某一礁区范围内,珊瑚礁生物多样性表现出水平分带和垂直分层的特征,礁前向海坡、礁坪和潟湖的生物在种类和数量上都有明显的差异[21]。
从目前的研究结果来看,美济礁3个区域浮游动物的种类组成相似性低,优势种组成、数量和多样性的区域差异明显。美济礁潟湖区浮游动物多样性较好,生物量最低;礁坪区浮游动物数量大但分布不均匀,且多样性水平最低;向海坡区浮游动物组成最为丰富、多样性非常丰富,但密度最低。聚类分析也表明,美济礁浮游动物的区域差异明显。而南沙渚碧礁潟湖和礁坪区浮游动物数量、优势种和多样性也存在明显的差异,太平洋和印度洋环礁的潟湖与邻近海域浮游动物的种类组成有明显的不同[1,5,17- 18,22- 23]。从相关分析来看,除海水温度外,其余环境因子对美济礁浮游动物影响不大,而海水温度的差异也主要是因3个区域水深条件不同而造成的。因此,美济礁向海坡、礁坪和潟湖3个区域空间异质性和水动力条件的差异,是造成浮游动物区域差异的主要原因。向海坡紧接外海,坡度陡峭、生境复杂,水深的梯度变化非常显著,并易形成上升流区,有利于多种类型的浮游动物生长,故向海坡浮游动物种类组成最为丰富、多样性程度也非常丰富。另外,向海坡区为高波能区,水动力最强,水体紊动强度大。高紊动强度影响浮游生物生长,导致浮游动物数量减少[24- 25 ]。因此,在3个区域中,向海坡区的浮游动物数量最低。
致谢:样品鉴定得到了厦门大学黄加祺教授和许振祖教授的帮助,中国水产科学研究院南海水产研究所邱永松研究员帮助写作,刘华雪和梁新负责外业调查和样品采集,课题组其他成员提供海水环境基础数据,并帮助分析,特此致谢。
[1] 尹健强, 陈清潮, 谭烨辉, 张建林. 南沙群岛渚碧礁春季浮游动物群落特征. 热带海洋学报, 2003, 22(6): 1- 8.
[2] 陈清潮, 尹健强. 黄岩岛环礁的浮游动物 // 南海海区综合调查研究报告(一). 北京: 科学出版社, 1982: 273- 277.
[3] 陈清潮, 张谷贤, 尹健强. 南沙群岛及其邻近海区调查研究报告(一). 上卷. 北京: 科学出版社, 1989: 99- 100.
[4] 章淑珍, 李纯厚. 南沙群岛珊瑚礁潟湖生态系统小型浮游动物的营养作用 // 南沙群岛珊瑚礁潟湖化学与生物学研究. 北京: 科学出版社, 1997: 64- 69.
[5] 尹健强, 黄良民, 李开枝, 熊兰兰. 南沙群岛珊瑚礁浮游动物多样性与群落结构. 生物多样性, 2011, 19(6): 685- 695.
[6] 尹健强, 黄晖, 黄良民, 李开枝, 练健生. 雷州半岛灯楼角珊瑚礁海区夏季的浮游动物. 海洋与湖沼, 2008, 39(2): 131- 138.
[7] 柯志新, 黄良民, 谭烨辉, 尹健强. 三亚珊瑚礁分布海区浮游生物的群落结构. 生物多样性, 2011, 19(6): 696- 701.
[8] 赵焕庭, 宁朝景, 朱袁智. 南沙群岛“危险地带”腹地珊瑚礁的地貌与现代沉积特征. 第四纪研究, 1992, 12(4): 368- 377.
[9] 国家技术监督局. 海洋调查规范—海洋生物调查. 北京: 中国标准出版社, 1991.
[10] 陈清潮, 黄良民, 尹健强, 张谷贤. 南沙群岛海区浮游动物多样性研究 // 南沙群岛及其邻近海区海洋生物多样性研究I. 北京: 海洋出版社, 1994: 42- 50.
[11] 连光山, 林玉辉, 蔡秉及. 大亚湾浮游动物群落的特征 // 大亚湾海洋生态文集(II). 北京: 海洋出版社, 1990: 180- 186.
[12] 韩洁, 张志南, 于子山. 渤海中、南部大型底栖动物的群落结构. 生态学报, 2004, 24(3): 531- 537.
[13] 袁伟, 张志南, 于子山, 邓可. 胶州湾西北部海域大型底栖动物群落研究. 中国海洋大学学报, 2006, 36(增刊): 91- 97.
[14] 郑重, 李少菁, 许振祖. 海洋浮游生物学. 北京: 海洋出版社, 1984: 1- 653.
[15] 郑重. 我国海洋浮游桡足类的生态习性与分布 // 郑重文集. 北京: 海洋出版社, 1987: 101- 133.
[16] 尹健强, 陈清潮, 张谷贤, 黄良民, 李开枝. 南沙群岛海区上层浮游动物种类组成与数量的时空变化. 科学通报, 2006, 51(增II): 129- 138.
[17] Goswami S C, Goswami U. Diel variation in zooplankton in Minicoy lagoon and Kavaratti Atoll (Lakshadweep Islands). Indian Journal of Marine Sciences, 1990, 19(2): 120- 124.
[18] Carleton J H, Doherty P J. Tropical zooplankton in the highly-enclosed lagoon of Taiaro Atoll (Tuamotu Archipelago, French Polynesia). Coral Reefs, 1998, 17(1): 29- 35.
[19] 宋星宇, 黄良民, 钱树本, 尹健强. 南沙群岛邻近海区春夏季浮游植物多样性研究. 生物多样性, 2002, 10(3): 258- 268.
[20] 孙宗勋, 赵焕庭. 南沙群岛珊瑚礁动力地貌特征. 热带海洋, 1996, 15(2): 53- 60.
[21] 于登攀, 邹仁林. 鹿回头库礁造礁石珊瑚物种多样性的研究. 生态学报, 1996, 16(5): 469- 475.
[22] Gerber R P. Species composition and abundance of lagoon zooplankton at Eniwetak Atoll, Marshall Islands. Atoll Research Bulletin, 1981, (247): 1- 22.
[23] Goswami S C. Production & zooplankton community structure in the lagoon & surrounding sea at Kavaratti Atoll (Lakshadweep). Indian Journal of Marine Sciences, 1983, 12(1): 31- 35.
[24] Visser A W, Stips A. Turbulence and zooplankton production: insights from PROVESS. Journal of Sea Research, 2002, 47(3): 317- 329.
[25] 朱宜平, 张海平, 李飞鹏, 陈玲. 水动力对浮游生物影响的围隔研究. 环境科学, 2010, 31(1): 70- 75.
The characteristics of summer zooplankton community in the Meiji coral reef,Nansha Islands, South China Sea
DU Feiyan1,2,3,*, WANG Xuehui1,2, LIN Zhaojin1,2
1SouthChinaSeaFisheriesResearchInstitute,ChineseAcademyofFisheryScience,Guangzhou510300,China2KeylaboratoryofFisheryEcologyandEnvironment,GuangdongProvince,Guangzhou510300,China3ScientificObservingandExperimentalStationofSouthChinaSeaFisheryResourcesandEnvironments,MinistryofAgriculture,Guangzhou510300,China
Zooplankton is an important component of the coral reef ecosystem, but studies on this component are insufficient in China. Based on 13 samples collected in July 2012, the characteristics of zooplankton community in the Meiji coral reef waters were studied. It was found that the reef waters are rich in zooplankton species and a total number of 138 species belonging to 15 taxa was recorded. The dominant species were fish eggs,Oikopleuralongicauda, Macrura larvae,Centropagesorsinii,Oikopleurasp.,Sagittarobusta, Brachyura zoea, andFritillariahaplostoma. According to habitats of the species, they could be categorized into warm-water coastal fauna and warm-water oceanic fauna. There was no obvious species dominating throughout three types of habitats in the reef waters and the species compositions and dominant species varied with habitats. The dominant species compositions in seaward slope were the most complex and 15 species were recorded. The main dominant species wereOithonaplumifera,ThaliademocraticaandOikopleurasp. in seaward slope. The similarities of species compositions of seaward slope to lagoon and to reef flat were only 27.8% and 21%, respectively, whereas, the similarity of species compositions between lagoon and reef flat was as high as 77.8%. The reef waters are rich in zooplankton diversities. The average indexes of species richness, diversity and evenness were 4.93, 3.33 and 0.67, respectively, and the overall threshold value of diversity was 2.43. The threshold values of seaward slope, lagoon and reef flat were 4.04, 1.98 and 1.38, respectively, showing that the zooplankton diversity was the highest in the seaward slope waters. The mean abundance and wet biomass of zooplankton were 117.70 ind/m3and 69.01 mg/m3, respectively, and the zooplankton abundance was mainly composed of larvae. The abundance of larvae was 52.32 ind/m3, taking up 44.5% of total zooplankton abundance. The zooplankton community in the reef waters could be classified into the lagoon-reef flat group and the seaward slope group. The test of One-way ANOSIM showed that compositions of the two groups were significantly different (R= 0.865,P= 0.001). Due to topographical heterogeneity and differences in hydrodynamic conditions in the reef waters, the regional variation of the zooplankton community was obvious. Being the low wave energy zone, the hydrodynamic changes, winds and waves of the lagoon is the least. In the lagoon waters, the zooplankton diversity was higher, but the biomass was the lowest. Whereas in reef flat, due to medium wave energy and moderate hydrodynamic changes, the zooplankton abundance and biomass was higher, with uneven distributions, and the diversity was lower. Adjoining the open ocean, seaward slope is steep in topography, drastic change in water depth, and upwelling can occur there. The diversified environment conditions are fit for the growth of a variety of zooplankton species. Therefore, the species richness and diversity of zooplankton in the seaward slope was the highest. However, located in the high wave energy zone, the hydrodynamic and water turbulence in the seaward slope are the strongest. The conditions do not favor zooplankton growth and the zooplankton abundance there was the lowest.
Nansha Islands; Meiji coral reef; zooplankton; community characteristics
中央级科研院所基本科研业务费专项资金(中国水产科学研究院南海水产研究所)资助项目(2012TS02); 农业部财政重大专项(NFZX2013); 国家科研支撑计划课题(2013BAD13B06)
2013- 05- 02;
日期:2014- 04- 11
10.5846/stxb201305020892
*通讯作者Corresponding author.E-mail: feiyanegg@163.com
杜飞雁,王雪辉,林昭进.南沙群岛美济礁海域夏季浮游动物群落特征.生态学报,2015,35(4):1014- 1021.
Du F Y, Wang X H, Lin Z J.The characteristics of summer zooplankton community in the Meiji coral reef, Nansha Islands, South China Sea.Acta Ecologica Sinica,2015,35(4):014- 1021.
