多年生黑麦草花粉活力和柱头可授性1)
2015-03-10翟飞飞巨关升李伟刘俊祥韩蕾孙振元
翟飞飞 巨关升 李伟 刘俊祥 韩蕾 孙振元
(中国林业科学研究院林业研究所,北京,100091)
多年生黑麦草花粉活力和柱头可授性1)
翟飞飞 巨关升 李伟 刘俊祥 韩蕾 孙振元
(中国林业科学研究院林业研究所,北京,100091)
以多年生黑麦草(Loliumperenne)品种‘Pinnacle’为材料,用TTC染色法和花粉管体外萌发法测定花粉在4种贮藏条件下(4 ℃常态、4 ℃干燥、-20 ℃常态和-20 ℃干燥)的活力变化;采用联苯胺—过氧化氢法测定开花前和开花后不同时段的柱头可授性。结果表明:多年生黑麦草花粉不耐贮藏,相比较而言,4 ℃干燥有利于其保存,7 d后花粉活力可维持在5%左右;而柱头可授性在开花前1 d至开花后5 h较强,该时间内授粉有利于受精作用,可提高结实率。
多年生黑麦草;花粉活力;柱头活性;TTC染色;花粉管体外萌发
WithLoliumperennecommercial variety ‘Pinnacle’, we used TTC staining method and pollen tube germination method in vitro to measure pollen viability under four storage conditions of 4 ℃ and room humidity, 4 ℃ and dry condition, -20 ℃ and room humidity, and -20 ℃ and dry condition. And we used benzidine-hydrogen peroxide method to study stigma. The pollen ofLoliumperennewas not resistant to storage, and 4 ℃ with dry condition was beneficial to its conserve relatively. Pollen viability maintained about 5% after 7 d. Stigma receptivity was strong between 1 d before blooming and 5 h after blooming, and this period was beneficial to fertilization and enhancing fruiting rate of flowers.
多年生黑麦草是禾本科黑麦草属的多年生草本植物,是一种重要的冷季型草坪草,其株型美观、建坪速度快、耐践踏,广泛用于运动场和高尔夫球场[1]。但其存在抗寒、抗热性差[2-3],对水分要求高[4]等缺点,因此,培育抗性强的多年生黑麦草新品种具有重要意义:一方面可提高其在园林绿化中的运用,增加景观效果,另一方面可缓解国内对国外草种的过分依赖。在种子生产及杂交育种过程中,为了进行人工辅助授粉和杂交授粉,通常需要采集和保存花粉,使有活性的花粉到达适宜的接受柱头,从而解决花时和花期不遇问题,提高杂交育种工作成功率。
花粉活力及柱头可授性因植物种类而异,研究发现多数禾本科植物的花粉活力丧失较快,如水稻花粉在田间条件下3 min内即有50%失去活力,5 min内几乎全部丧失;小麦花粉放置5 h,授粉后结实率降低到6.4%;高粱花粉采下5 h,再授粉则不能获得任何种子;玉米花粉的活力在自然条件下仅能维持1~2 d[5-6]。多年生黑麦草是一种疏丛型植物,为异花授粉类型,且自交不亲和,目前,未见有关其花粉活力及柱头可授性方面的报道。文中对多年生黑麦草花粉活力及柱头可授性进行研究,旨在找到适合多年生黑麦草花粉的最佳贮藏条件及柱头最佳可授期,为以后的杂交育种工作合理有序进行提供参考,进而培育出多年生黑麦草新种质。
1 材料与方法
本试验材料为多年生黑麦草品种‘Pinnacle’。种子于7月16日播种于直径为10.0 cm的营养钵中,栽培基质V(泥炭)∶V(珍珠岩)=6∶1,每盆施0.5 g缓释肥(Osmocotte,M5-6,质量分数w(N)∶w(P)∶w(K)=14∶13∶13)。温室温度白天22~25 ℃,晚上16~19 ℃,相对湿度50%,最大光强为400 μmol·m-1·s-1。每7 d浇1次水。9月15日移栽至中国林业科学研究院玉泉山苗圃,并进行正常田间养护管理;第2年5月中旬开花后,取花粉、柱头进行试验。
于盛花期上午09:30—11:30,用镊子夹取露出颖壳、即将开裂的黄色花药置于直径为60 mm的小培养皿中。采集完后放入冰盒,立即带回实验室进行处理:4 ℃常态、4 ℃干燥、-20 ℃常态、-20 ℃干燥。干燥:在直径为90 mm的大培养皿中放入适量硅胶,将小培养皿置于大培养皿内,并用自封袋包裹进行干燥处理。常态:大培养皿内不加硅胶,小培养皿置于大培养皿内。然后,将干燥和常态处理的花粉放入冰箱4 ℃和-20 ℃条件下,进行不同温度保存。分别在保存1、2、4、7 d测定花粉活力。另外,在田间条件下,取下花药5、10 min和1 h后,分别测定花粉活力。
花粉活力的测定:TTC(2,3,5-氯化三苯基四氮唑)染色法采用高俊凤[7]的方法。在载玻片上滴一滴0.5% TTC溶液,用解剖针蘸取花药置于载玻片上的溶液中,将花药破碎,盖上盖玻片,将载玻片置于事先备好的放有湿润滤纸的培养皿中,然后连同培养皿放置于25 ℃的黑暗环境下培养。25 min后显微镜下观察,统计盖片中央5个视野中红色花粉所占比例,计算平均值并对数据进行统计分析。每种处理材料观察3个制片。试剂的配制,即称取0.50 g TTC放入烧杯中,加入少许95%酒精使其溶解,然后用蒸馏水稀释至100 mL。溶液避光保存,若发红则不能再用。
花粉管体外萌发法采用胡适宜[8]的方法。首先配制适合花粉萌发的培养基,即称10.0 g蔗糖、0.01 g硼酸(HBO3)、0.03 g硝酸钙(Ca(NO3)2·4H2O)、0.02 g硫酸镁(MgSO4·7H2O),0.01 g硝酸钾(KNO3)溶解于100 mL蒸馏水中,置于冰箱4 ℃保存备用。用滴管加两滴培养液于凹槽培养皿中,用解剖针蘸取花药置于载玻片上的溶液中,将花药破碎,盖上盖玻片,将载玻片置于事先备好的放有湿润滤纸的培养皿中,然后连同培养皿放置于25 ℃的环境下,室温下进行孵育,3 h后显微镜下观察花粉萌发情况。每种处理材料观察3个制片,盖片中央选5个视野,统计其萌发率,计算花粉萌发百分率。
柱头可授性的测定:采用联苯胺—过氧化氢法[9],联苯胺—过氧化氢反应液为V(1%联苯胺溶液)∶V(3%过氧化氢溶液)∶V(水)=4∶11∶22的混合液。在开花盛期,采开花后不同天数的花朵,将其柱头浸入含有联苯胺—过氧化氢反应液的凹面载玻片凹陷处。若柱头具有可授性,则表现过氧化物酶活性,使周围的反应液呈现蓝色并伴有大量气泡,反之则无。每天取5~10个柱头进行测定,并且在第1天取样时标记好其他开放的花。试剂的配制为1%联苯胺溶液,即称1.0 g联苯胺固体溶于100 mL 50%的酒精溶液,装入棕色试剂瓶备用;3%过氧化氢,即取10.0 mL 30%的过氧化氢和90.0 mL蒸馏水混合,装入试剂瓶中备用。反应液配好之后放入棕色试剂瓶内,并于冰箱4 ℃保存备用。
数据处理:采用Excel 2007软件进行数据统计和图表绘制;应用SPSS 18.0软件进行统计分析,采用单因素方差分析和Duncan多重比较检验不同处理间的差异。
2 结果与分析
2.1 短时间内花粉活力
由表1可知,两种测定方法所得结果较为一致,多年生黑麦草花粉采集5、10 min和1 h,其活性无显著差异;经TTC染色法测定的花粉活性均比花粉管体外萌发法高,前者最高为48.68%,后者最高仅为26.19%(表1)。可见,多年生黑麦草花粉活力在5 min之内丧失较快,具活力花粉的活性可维持一段时间,且在1 h之内变化不显著。
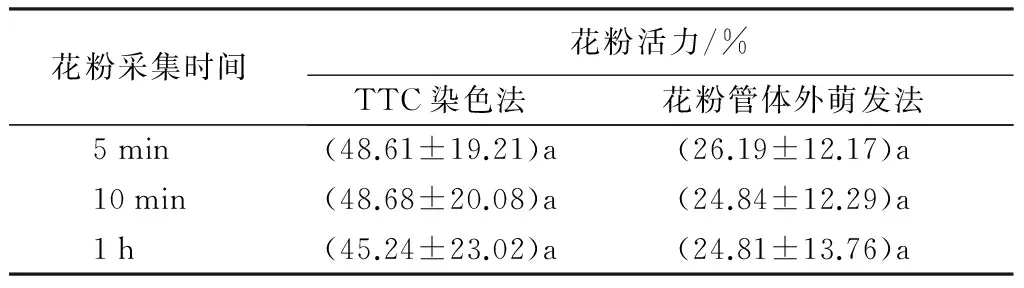
表1 短时间内花粉活力测定
注:表中数据为15次测定的平均值±标准误;同列相同小写字母表示同一方法、不同时间下无显著差异(P>0.05)。
2.2 不同贮藏条件对花粉活力的影响
由表2可知,随着时间延长,4种贮藏条件下多年生黑麦草花粉活力均下降,且TTC染色法比花粉管体外萌发法所测活力高。TTC染色法测定结果表明,花粉采集1 d后,-20 ℃(常态和干燥)下利于花粉活力保持。采集4 d后,4 ℃(常态和干燥)条件下,花粉活力下降不显著,而-20 ℃(常态和干燥)条件下则显著降低。7 d后,4种条件下花粉活力均显著下降,以-20 ℃常态下降幅度最大,为95.99%(相对于表1中1 h时花粉活力,下同),4 ℃干燥下降87.34%;且4 ℃干燥条件的花粉活力显著高于-20 ℃常态和-20 ℃干燥,为5.73%(P<0.05)。花粉管体外萌发法测定结果表明,采集2 d后,花粉活力下降不显著,4 d后仅-20 ℃干燥条件下花粉活力显著下降。7 d后,4种贮藏条件下的花粉活力均显著下降,其中-20 ℃(常态和干燥)下降幅最大,约为90%;4 ℃(常态和干燥)状态下花粉活力显著高于-20 ℃(常态和干燥)条件下的,分别为4.98%和5.70%。以上结果可知,4 ℃常态较利于多年生黑麦草花粉保存。
2.3 柱头可授性
由表3可知,多年生黑麦草柱头在开花前1 d已具较强可授性,即联苯胺—过氧化氢法染色颜色深,气泡数量多,柱头分泌大量黏液;开花后1 h柱头可授性较强,与开花前相当;开花后5 h,柱头分泌黏液减少,染色较深,气泡较多,可授性减弱;开花后1 d,柱头可授性明显降低,染色较深,气泡少量或较多,黏液分泌较少;开花后2 d,柱头过氧化氢酶活性减弱至无,已不再具有可授性。由以上结果可知,多年生黑麦草柱头在开花前1 d至开花后1 h,具有较强可授性。

表2 不同贮藏条件对花粉活力的影响
注:表中数据为15次测定的平均值±标准误;同行相同小写字母表示同一测定方法和同一贮藏条件、不同时间下无显著差异(P>0.05),同行不同小写字母表示同一测定方法和同一贮藏条件、不同时间下有显著差异(P<0.05);同列同一测定方法和同一时间、不同贮藏条件下相同大写字母表示无显著差异(P>0.05),同列同一测定方法和同一时间、不同贮藏条件下不同大写字母表示有显著差异(P<0.05)。

表3 多年生黑麦草柱头可授性测定
注:颜色变化,+为浅,++为较深,+++为深;气泡数量,+为少,++为较多,+++为多;柱头分泌黏液,+为柱头分泌黏液,++为柱头分泌较多黏液,+++为柱头分泌大量黏液。
3 结论与讨论
植物花粉活力大小、维持时间长短,既由自身遗传基因决定,又受环境因素影响,如温度、湿度、气压等[10]。刺五加的花粉活力可维持4 d,且花粉活力及下降速度受生境条件影响[11];锦带花在开花后6 d,仍有15.38%的花粉具有活力[12];连翘的花粉活力在开花初期不高,随后逐渐上升,开花第2 d后达到最高峰,为94.25%[13];而银杏的花粉活力在低温干燥条件下可维持1 a[14]。对于禾本科植物花粉而言,其活力丧失较快,贮藏条件不同于其他植物,花粉采集后,如何对花粉进行合理有效保存至关重要。本研究表明,多年生黑麦草花粉活力丧失较快,采下5 min后即有一半以上花粉失活,但是在短期(1 h)内活性基本不变;在采集花粉后1~24 h活力又丧失一半,活力丧失较快,还需对此段时间内花粉活力进行研究。多年生黑麦草花粉不耐贮藏,相比较而言,4 ℃干燥条件下花粉活力较高,此贮藏条件有助于提高多年生黑麦草的结实率。
TTC染色法是根据花粉粒呼吸作用强弱判断花粉活力的,花粉中呼吸酶活性可较准确反映其活力强弱,同时氧化还原染料能在呼吸酶作用下染色,所测花粉活力值较大[15]。成熟花粉具有较强生活力,在适宜条件下便能萌发和生长,花粉管体外萌发法是通过培养基培养来判断其活力的[16]。但是体外萌发法易受花粉萌发条件如培养时间和温度、营养物质、矿质元素等的影响[17],并且培养基和柱头存在一定差距,故所测结果偏低。本研究中,虽然TTC染色法测得花粉活力高于体外萌发法,但两者趋势一致,从不同角度反映花粉活力,结果说服力较强。
柱头可授期是花朵成熟过程中的一个重要时期,在很大程度上影响自花传粉率、开花不同阶段传粉成功率、各种传粉者的相对重要性、雄性和雌性功能之间的相互干扰、不同基因型花粉之间的竞争以及配子体选择的机会等[18]。柱头可授期因植物不同而有所差异,如甜樱桃(Cerasusavium)柱头可授期从开花前1 d开始可持续5~7 d[19],梨(Pyruscommunis)开花时柱头还没发育成熟,开花2 d后柱头才具有可授性[20];并且花朵年龄、一天中开花时间及柱头分泌物等均影响柱头可授性[21]。本研究中,多年生黑麦草柱头可授性持续时间较短:开花前1 d至开花后1 h内,柱头具有最强可授性;而在开花5 h内,具较强可授性。因此,对于多年生黑麦草,最佳授粉时期是在花朵开放后1 h内进行,最好在5 h内完成,这样可提高授粉成功率。
根据上述研究结果,多年生黑麦草开花后,花粉和柱头在短时间内均具有较强活性,但是多年生黑麦草自交不亲和,可能是花粉和柱头的SI蛋白相互作用,引起胞内游离Ca2+质量浓度迅速增加,激活了Ca2+依赖蛋白激酶,在蛋白激酶作用下,蛋白质发生磷酸化,一系列级联反应之后,激活相关基因表达阻止花粉萌发;引起细胞骨架肌动蛋白解聚,柱头细胞程序性死亡,最终导致自交不亲和[22]。因此,对多年生黑麦草花粉活力及柱头可授性进行研究,找到适合其花粉的贮藏条件以及最佳柱头可授期,可为杂交育种提供理论依据,有利于多年生黑麦草新品种选育。
[1] 翟飞飞,韩蕾,刘俊祥,等.人工低温胁迫下多年生黑麦草诱变株系的抗寒性研究[J].草业学报,2013,22(6):268-279.
[2] Gusta L V, Butler J D, Rajashekar C, et al. Freezing resistance of perennial turfgrass[J]. HortScience,1980,15(4):494-496.
[3] 石永红,万里强,刘建宁,等.多年生黑麦草高温半致死温度与耐热性研究[J].草业科学,2010,27(2):104-108.
[4] 万里强,石永红,李向林,等.高温干旱胁迫下三个多年生黑麦草品种叶绿体和线粒体超微结构的变化[J].草业学报,2009,18(1):25-31.
[5] 潘瑞炽,董愚得.植物生理学:下册[M].北京:高等教育出版社,1984:97.
[6] 李要民,陈良碧.不同温湿条件下贮藏的3种禾本科植物花粉活力变化(简报)[J].植物生理学通讯,1998,34(1):35-37.
[7] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006:183-184.
[8] 胡适宜.植物胚胎学实验方法(一):花粉生活力的测定[J].植物学通报,1993,10(2):60-62.
[9] 张香桂,倪万潮,沈新莲,等.棉花柱头可授性测定方法的研究[J].种子科技,2012(1):25-26.
[10] 陈灵鸷,杨春华,张文君,等.扁穗牛鞭草花粉活力及柱头可授性研究[J].中国草地学报,2009,31(6):59-63.
[11] 刘林德,张洪军,祝宁,等.刺五加花粉活力和柱头可授性的研究[J].植物学研究,2001,21(3):375-379.
[12] 刘林德,张萍,张丽,等.锦带花的花粉活力、柱头可授性及传粉者的观察[J].西北植物学报,2004,24(8):1431-1434.
[13] 铁军,金山,茹文明,等.连翘花粉活力和柱头可授性研究[J].山西大学学报:自然科学版,2008,31(4):604-607.
[14] 孙霞,刑世岩,路冬,等.银杏花粉活力研究[J].果树科学,1998,15(1):58-64.
[15] 胡春,刘左军,李富香,等.纯裂银莲花花粉活力测定方法的研究[J].植物研究,2013,33(5):582-586.
[16] 邢虎成,揭雨成,周清明,等.苎麻花粉活力测定方法研究[J].中国农学通报,2010,26(17):65-69.
[17] 孙坤,张玉娜,苏雪,等.中国沙棘花粉生活力检测方法比较研究[J].西北师范大学学报:自然科学版,2009,45(2):78-81.
[18] Dafni A. Pollination Ecology[M]. New York: Oxford Univ Press,1992:59-89.
[19] 王丽娟,刘林德,张莉,等.烟台甜樱桃柱头的可授性、形态特征与坐果率[J].植物学报,2011,46(1):44-49.
[20] Sanzol J, Rallo P, Herrero M. Stigmatic receptivity limits effective pollination period in ‘Agua de Aranjuez’ pear[J]. Journal of the American Society for Horticultural Science,2003,128:458-462.
[21] 叶要妹,张俊卫,齐迎春,等.百日草柱头可授性和花粉生活力的研究[J].中国农业科学,2007,40(10):2376-2381.
[22] 牛俊海,鲁晓民,汤继华,等.禾本科植物自交不亲和性及其分子生物学研究进展[J].分子植物育种,2006,4(2):269-274.
Pollen Viability and Stigma Receptivity ofLoliumperenne
Zhai Feifei, Ju Guansheng, Li Wei, Liu Junxiang, Han Lei, Sun Zhenyuan(Research Institute of Forestry, Chinese Academy of Forestry Sciences, Beijing 100091, P. R. China)/Journal of Northeast Forestry University,2015,43(3):75-78.
Loliumperenne; Pollen viability; Stigma receptivity; TTC staining method; Pollen tube germination method in vitro
1)国家“863”计划项目(2011AA100209)。
翟飞飞,女,1987年5月生,中国林业科学研究院林业研究所、国家林业局林木培育重点实验室(中国林业科学研究院),博士研究生。E-mail:lkyzff@163.com 。
韩蕾,中国林业科学研究院林业研究所、国家林业局林木培育重点实验室(中国林业科学研究院),研究员。E-mail:hl04192003@aliyun.com。
2014年8月5日。
S512.5; Q944.42
责任编辑:任 俐。
