三峡水库秀丽白虾生长与繁殖生物学特征研究
2015-03-03陈思宝廖传松赵修江叶少文李钟杰张堂林刘家寿
陈思宝廖传松赵修江叶少文李钟杰张堂林刘家寿
(1. 中国科学院水生生物研究所, 淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 中国科学院大学, 北京 100049;3. 中国长江三峡集团公司科技环保部, 北京 100038)
三峡水库秀丽白虾生长与繁殖生物学特征研究
陈思宝1,2廖传松1,2赵修江3叶少文1李钟杰1张堂林1刘家寿1
(1. 中国科学院水生生物研究所, 淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 中国科学院大学, 北京 100049;3. 中国长江三峡集团公司科技环保部, 北京 100038)
摘要:2012年11月至2013年10月逐月对三峡水库木洞江段秀丽白虾的生长和繁殖生物学特征进行了研究。结果显示秀丽白虾是一年生的虾类, 寿命为12—14个月。雌性个体的平均体长(37.72±7.92) mm显著大于雄性(37.08±6.59) mm, 具有明显的雌雄异形现象。秀丽白虾的渐近体长(L∞)为 59.33 mm, 生长系数(K)是 1.6/年, 生长方程为:Lt=59.33[–e–1.6(t+0.081)]。雌虾繁殖季节从4月上旬持续至9月下旬, 繁殖高峰期为4月中旬至6月中旬, 由越冬虾和当年虾两个世代的繁殖群体组成。抱卵雌虾的体长为27.07—53.71 mm, 平均体长(L50)为 37.06 mm。雌雄性比月变化为 1.11—2.36, 平均为 1.46, 群体中雌性个体数量显著多于雄性(P <0.001)。绝对繁殖力(FA)为 31—294粒, 相对繁殖力(FR)为(115±22)粒/g。绝对繁殖力与体长呈幂函数关系(FA=0.0031L2.8632, r=0.72, n=106), 而与体重呈直线关系(FA=104.63W + 9.9534, r=0.79, n=106)。卵径(D)平均值为(1215±102) μm。为合理保护和利用三峡水库秀丽白虾种群资源, 建议加强虾类捕捞管理, 繁殖期间(4—9月)禁止捕捞虾类或限制捕捞强度。
三峡水库; 秀丽白虾; 生长; 繁殖生物学
秀丽白虾(Exopalaemon modestus Heller, 1862)隶属于甲壳纲(Crustacea), 十足目(Decapoda), 长臂虾科(Palaemonidae), 白虾属(Exopalaemon), 主要分布在中国、朝鲜半岛和西伯利亚等地区[1]。秀丽白虾是我国淡水水域的主要经济虾类, 其产量仅次于日本沼虾(Macrobrachium nipponensis)[2], 巢湖[3,4]、太湖[5]等大型湖泊中资源丰富。在水域生态系统中,秀丽白虾既是捕食者又是消费者, 在食物链中发挥着重要作用[6]。目前有关秀丽白虾种群生态学的研究报道较少, 主要集中在湖泊中[4,5,7—10]。施炜纲等[5]、温周瑞等[8,9]、赖伟等[10]先后研究了太湖秀丽白虾的生长、繁殖和食性, Oh等[7]研究了朝鲜半岛南部入海口的Young-am湖秀丽白虾的生长和繁殖特征。
秀丽白虾是三峡水库重要经济虾类之一, 广泛分布于库区干流、库湾及支流中, 在渔业资源中占有重要的地位。然而, 以往水库渔业与水生生物学的研究主要围绕鱼类资源展开, 虾类资源方面却鲜有报道或研究, 仅有张亮[11]利用稳定同位素分析了三峡水库各江段日本沼虾、秀丽白虾的营养级差异。目前, 虾类的捕捞存在不合理现象, 长江上游的禁渔期为2—4月, 忽略了虾类的繁殖期在4—9月期间, 容易对虾类资源造成破坏。作者于 2012年 11月至2013年10月逐月对三峡水库秀丽白虾的生长和繁殖生物学特征进行了详细研究, 以期为合理利用和保护库区秀丽白虾资源提供基础资料。
1 材料与方法
1.1 研究区域概况
2012年11月至2013年10月逐月(下旬)采样,采样点设置在三峡水库上游的木洞镇, 位于长江干流和木洞小河的交汇处(106.84°E, 29.58°N)(图 1),距离三峡大坝约600 km。该江段水流较急, 且水位受水库周期性蓄水调节影响(水位在145—175 m运行)。
图1 采样点在三峡水库的位置Fig. 1 The location of sampling sites in the Three Gorges Reservoir of China
1.2 虾类采样和数据采集
采集样本的渔具为地笼, 网目(2a)为0.7 cm, 同时使用HOBO温度记录仪监测水温变化。每月测定秀丽白虾 100—200尾, 使用电子游标卡尺(精确到0.01 mm)、电子天平(精确到0.001 g)分别测定体长(L)、体重(W)。体长为眼柄基部至尾节末端的直线长度, 体重是干纱布吸去体表水分后的湿重。根据肉眼观测, 鉴定雌雄[2]。繁殖期间, 计数抱卵雌虾,确定雌虾抱卵率, 测量体长、体重后, 保存在 10%福尔马林溶液中, 带回实验室备检。清水冲洗后, 用镊子刮取抱卵雌虾腹部的虾卵, 在解剖镜下测定每尾样本的卵粒总数N、卵径D(用长和宽的平均值表示, 精确到1 μm)。
1.3 生长和繁殖的研究方法
根据秀丽白虾的体长频率分布, 运用FiSAT Ⅱ (FAO-ICLARM Fish Stock Assessment Tools, Version 1.2.2)软件中 ELEFAN(Electronic Length Frequency Analysis)来 分析其生长[12], 获 得VBGF(Von Bertalanffy Growth Function)生长方程: Lt= L∞[1–e–K(t–t0)] (1)
由图5可知,本文提出的模型最有利于对抗开关攻击。本文所提模型减慢了信任上升速度,并加快了信任值的下降速度。当节点j展开攻击以后,需要在很多周期内连续伪装为正常节点才能把自身信任提升到一个较高的水平。这使得节点j难以把自身信任水平维持在高处,其他节点可以更快地识别与隔离此类节点。对整个仿真周期(0~300)进行统计,本文所提模型比TEMBB与EMNT的平均信任值分别下降了18.5%和16.4%。
log10(–t0) = –0.392–0.275 log10L∞–1.038log10K[13](2)
式中 Lt为 t龄虾的体长, L∞是渐近体长, K(/年)是VBG 生长系数, t0是Lt=0时的理论年龄(月龄)。
体长(L)与体重(W)关系通过公式 W=aLb进行回归拟合, 用t检验比较b值是否与3(即等速生长状态下的b值)有显著性的差异。
根据雌虾抱卵率(抱卵雌虾占全部雌虾的百分率)判断繁殖期和繁殖高峰期。绝对繁殖力(FA)为抱卵雌虾的抱卵总数(N), 相对繁殖力用单位体重的卵粒数(FR=N/W)表示。
用逻辑斯蒂回归模型计算抱卵雌虾的比例, 50%的抱卵个体平均体长即为种群的平均抱卵大小(L50)[14]。
P = 1/{1+exp [–(a+bL)]} (3)
L50=–a/b (4)
式中a、b是参数。
性比为雌、雄性个体数量的比值, 使用卡方检验检(χ2test)检测秀丽白虾的雌雄性比是否符合1︰1[15]。
数据分析在SPSS V22.0和R V3.1.1软件中进行,以P<0.05作为数据达到差异显著性的标准。
2 结果
三峡水库木洞江段周年采样期间, 共采集1790尾秀丽白虾, 其中雌虾1062尾(含抱卵雌性106尾)、雄虾728尾。
2.1 种群结构和生长
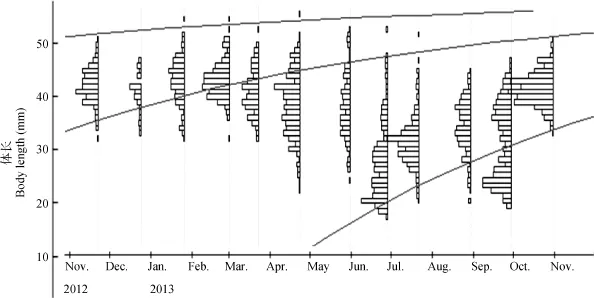
图2 ELEFAN估算三峡水库秀丽白虾周年生长曲线Fig. 2 The growth curves estimated with ELEFAN for E. modestus in the Three Gorges Reservoir
从秀丽白虾种群的体长频率分布月变化(图 2)推测其为一年生的淡水虾类, 寿命约为 12—14个月。2012年11月到2013年5月和2013年6月至10月分别为一个世代种群, 其中 2013年 6月到 9月为新老种群世代交替过程, 新出生虾类的数量和个体大小逐月增长, 而上一世代的虾类数量则逐渐减少, 直至9月份世代交替结束。ELEFAN分析表明秀丽白虾的渐近体长(L∞)为 59.33 mm, 生长系数(K)是1.6, 估算的生长方程为: Lt= 59.33[–e–1.6(t+0.081)]。
雌、雄个体体长均值月变化如图3所示, 秀丽白虾的体长均值从11月到2月逐渐增长(除1月份外),在2月达到峰值, 然后逐渐降低(雌虾除4月份外), 雄虾在6月降到最低值, 而雌虾7月处于最低值, 之后体长均值逐渐上升(除9月份外, 由于繁殖个体死亡, 新生群体增加)。2012年11月至2013年3月及2013年7月、9月雌雄个体的体长大小没有显著性差异(Mann-Whitney test, P>0.05, 表1), 其余月份雌性个体的体长均显著性地大于雄性(Mann-Whitney test, P<0.05, 表1)。比较秀丽白虾雌、雄全部个体的大小, 发现雌性个体的平均体长是(37.72±7.92) mm (mean±SD, n=1062),显著性大于雄性个体平均体长(37.08±6.59) mm (n=728) (Mann-Whitney Test, W=571557, P=0.02)。
2.2 体长与体重关系
秀丽白虾体长与体重关系如下:
雌性: W =1.2185×10–5L3.042(n=1062, r2=0.973)
雄性: W =1.2849×10–5L3.018(n=728, r2=0.985)
t检验表明, 雌、雄个体的b值均显著性地大于3 (P<0.05), 秀丽白虾雌、雄的生长式型均为正异速生长(图4)。
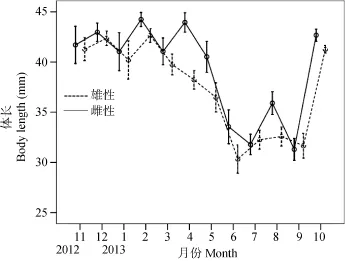
图3 三峡水库秀丽白虾体长逐月变化Fig. 3 The monthly change in the body length of E. modestus in the Three Gorges Reservoir
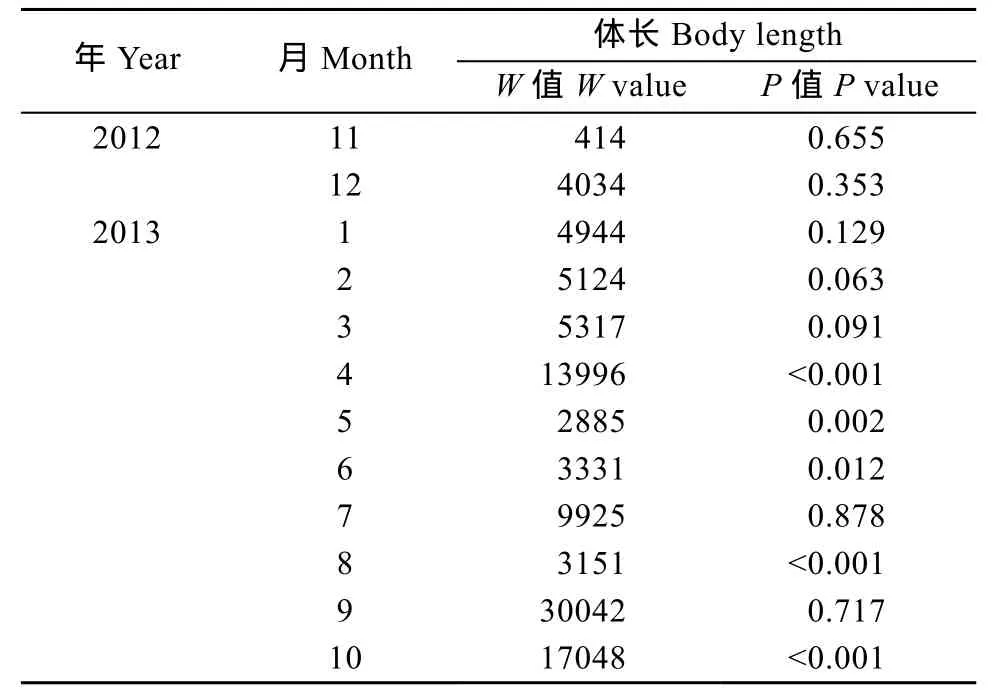
表 1 三峡水库秀丽白虾雌、雄个体体长差异的 Mann-Whitney检验分析Tab. 1 The sexual differences in the body length of E. modestus tested with Mann-Whitney method in the Three Gorges Reservoir
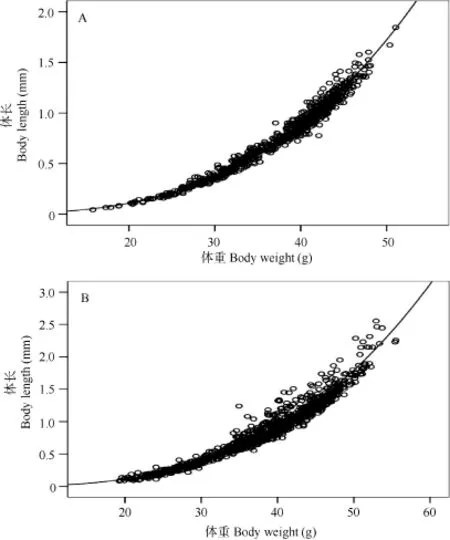
图4 三峡水库秀丽白虾雄性(A)和雌性(B)体长-体重关系Fig. 4 The relationship between the body length and the body weight of male (A) and female (B) E. modestus in the Three Gorges Reservoir
2.3 繁殖生物学
繁殖期与繁殖群体组成 经过周年采样观察,木洞江段出现抱卵雌虾的时间为4月上旬至9月下旬, 由越冬虾(前一年出生种群)和当年虾(当年出生的种群)两个世代的繁殖群体组成, 繁殖期间的水温变化范围为17.1—34.3 ℃(年平均水温19.4 ℃ ,图5)。
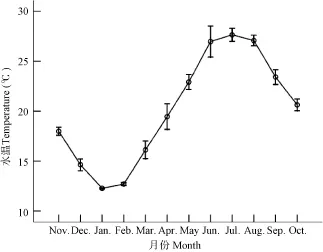
图5 三峡水库木洞江段水温月变化Fig. 5 The monthly variation of water temperature in the Mudong section in the Three Gorges Reservoir
4月上旬开始出现抱卵雌虾, 此时抱卵雌虾主要由越冬虾组成。繁殖高峰期出现在 4月中旬至 6月中旬, 雌虾抱卵率均在50%以上, 5月下旬雌虾抱卵率最高达到66.2%。在6月下旬至7月下旬抱卵率逐步下降, 7月下旬的雌虾抱卵率仅为17%, 即越冬虾的繁殖期高峰期结束。随着当年虾性成熟并加入繁殖群体中, 8月下旬又达到了繁殖高峰期, 抱卵率为49.5%, 9月下旬繁殖活动结束(表2)。
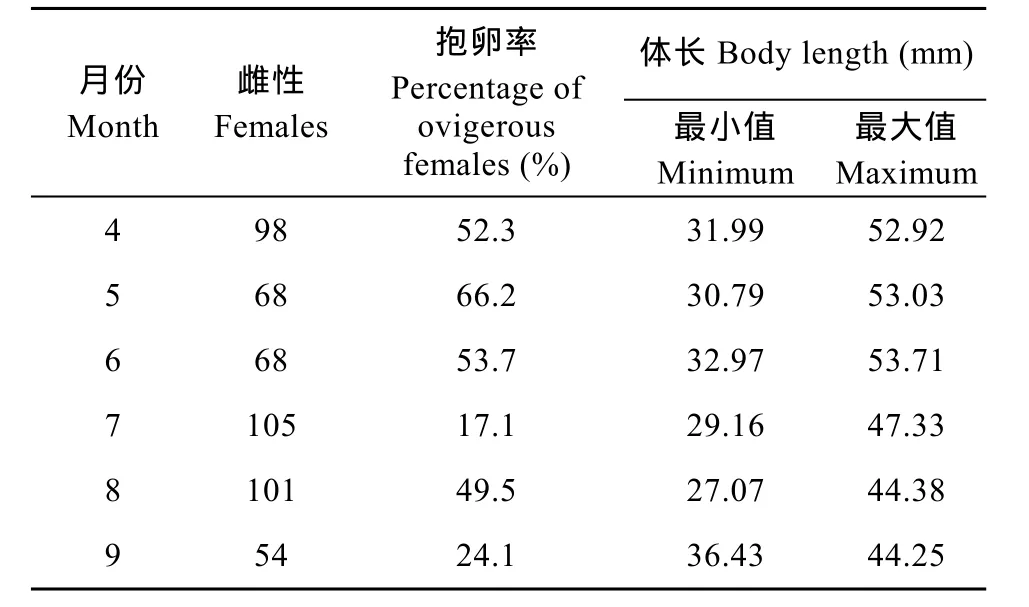
表2 三峡水库秀丽白虾雌虾抱卵率及体长组成Tab. 2 Percentage of ovigerous females and their minimum and maximum body lengths in the Three Gorges Reservoir
抱卵雌虾的最小体长是27.07 mm, 最大体长是53.71 mm, 雌虾体长与抱卵比例的逻辑斯蒂方程式为: P = 1/[1+exp (8.969–0.242L)], 估算出木洞江段秀丽白虾雌性个体抱卵时的平均体长(L50)为37.06 mm (图6)。
性比 三峡水库木洞江段秀丽白虾性比在不同月份间具有明显差异(图 7), 在 1.11—2.36之间,年平均性比为 1.46, 雌性数量显著多于雄性数量(χ2test, χ2=51.20, df=1, P<0.001, n=1790)。
繁殖力与卵径 秀丽白虾的绝对繁殖力(FA)为(127±53)粒(n=106), 相对繁殖力(FR)为(115±22)粒/g (n =106), 卵径(D)为(1215±102) μm (n=4171)。秀丽白虾的绝对繁殖力为 31—294粒, 随着体长和体重增加而增加(图 8), 与体长呈幂指数相关(FA= 0.0031L2.8632, r=0.72, n=106), 而与体重呈直线相关(FA=104.63W+9.9534, r=0.79, n=106)。
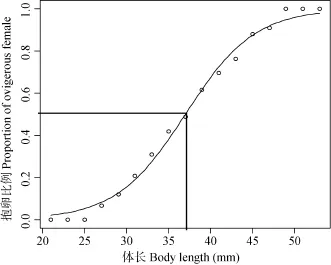
图6 逻辑斯蒂函数拟合三峡水库秀丽白虾雌虾抱卵体长Fig. 6 The logistic function describing the relationship between the proportion of ovigerous females and the body lengths L50corresponded to 50% of ovigerous females in the Three Gorges Reservoir
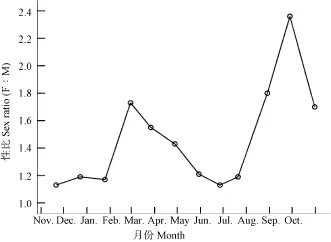
图7 三峡水库秀丽白虾性比(雌︰雄)月变化Fig. 7 The monthly variation in the sex ratio of E. Modestus in the Three Gorges Reservoir
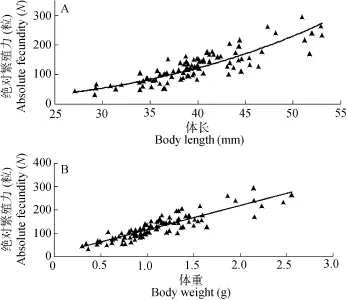
图8 三峡水库秀丽白虾体长(A)、体重(B)与绝对繁殖力的关系Fig. 8 The relationship between the absolute fecundity and the size (body length: A, body weight: B) of E. modestus in the Three Gorges Reservoir
3 讨论
3.1 生长和繁殖
长臂虾科的虾类寿命差异较大[7,16—19], 很多虾类的生命周期较短, 如 Palaemon xiphias的寿命为14—17月[16], 太平长臂虾(P. pacificus)的寿命为12—15月, 锯齿长臂虾(P. serrifer)为13—16月[17]。三峡水库木洞江段秀丽白虾种群结构表明, 6月至9月处于世代交替过程, 繁殖群体在繁殖结束后逐渐死亡, 新出生的群体快速生长。结合秀丽白虾体长频率月变化, 分析其寿命约为12—15月, 与朝鲜半岛南部入海口的 Young-am 湖的研究结果(13—16月)相似[7]。研究认为[7,16]虾类的生长和寿命与纬度(水温)有显著性的关系, 低纬度的种群, 性成熟早、寿命短。三峡水库秀丽白虾生长系数(K=1.6)显著大于Young-am 湖[7]生长系数(雌性为0.58, 雄性为0.62)。
三峡水库木洞江段秀丽白虾4月上旬出现抱卵雌虾, 持续至9月下旬, 与其他水域研究结果[7,8,10]相似, Young-am湖[7]秀丽白虾出现抱卵雌虾的时间是5月至9月, 太湖[8,10]繁殖期是4月下旬至9月下旬。研究表明[20,21]长臂虾科种类的繁殖周期与温度、光照、食物等因素有关, 其中水温与虾类卵巢发育密切相关, 决定虾类的产卵次数[22]。长臂虾科中有些种类在繁殖期间可以产卵多次, P. xiphias抱卵次数可以达到2—6次[16], 日本沼虾(M. nipponensis)可以产卵4—6批[23]。三峡水库秀丽白虾繁殖群体由越冬虾和当年虾组成, 7月下旬越冬虾的繁殖期基本结束, 8月至9月的繁殖群体主要为当年虾, 通过对秀丽白虾抱卵雌虾的卵粒鉴定, 同一批抱卵雌虾的卵粒具有两个不同的发育期。根据繁殖期间的水温,可以推断秀丽白虾的产卵次数至少可以达到 2次,还需要室内实验来深入研究。
三峡水库秀丽白虾种群中雌性个体数量显著多于雄性, 性比为 1.46, 与其他研究的结果是一致的[7,8,10]。在秀丽白虾产量丰富的太湖[7,10]都发现雌雄性比为1.4左右, 在Young-am 湖[7], 秀丽白虾雌性个体数量也显著性多于雄性个体数量。性比可能与种群的生长、死亡、行为等因素有关[21]。根据Oh等[7]的研究, 秀丽白虾的雌性生长率显著大于雄性。Berglund[24]认为雄性的生长率小于雌性, 有助于减少其生长耗能以及降低被捕食的压力, 从而导致种群中雌性的比例较多。Kim[25]认为性比的差异可能是由于雌、雄死亡率不同, 或者种群行为特征(如迁移)具有差异。长臂虾科的种类具有明显的雌雄异形现象[16,22,25—29], 雌虾比雄虾具有更大的个体。三峡水库秀丽白虾雌性体长(37.72 mm)显著地大于雄性(37.08 mm), 与其他水域的研究结果一致[7, 8]。
3.2 种群资源管理
秀丽白虾是三峡水库渔业资源重要的组成部分,为保护和利用种群资源, 根据其生活史特征提出以下建议: (1) 繁殖期间禁捕, 控制起捕规格。要增加世代补充量, 必须保证足够的亲虾进行产卵繁殖以及保护幼虾[20]。三峡水库禁渔期(2月1日至4月30日)过后的5月至8月捕捞强度高, 而秀丽白虾正处于产卵高峰期, 大量亲虾被捕捞, 破坏了种群结构,为保护库区虾类资源, 5—8月份也应禁止捕捞秀丽白虾, 至少应该限制捕捞量。三峡水库地笼网目多为 0.7 cm, 制定合理的渔具网目规格可以有效地避免捕捞幼虾, 应将网目大小改为1.5 cm及以上, 确保秀丽白虾起捕体长控制到35 mm以上。(2) 发展优质高效的水库渔业。目前库区鱼类资源已经呈现衰退趋势, 渔获物小型化、低龄化现象明显[30,31],急需恢复鱼类资源。已有研究表明虾类可以作为黄颡 鱼 (Pelteobagrus fulvidraco)[32]、 鳜 (Siniperca chuatsi)[33]天然饵料。结合三峡水库虾类资源和小型鱼类资源, 发展食鱼(虾)性鱼类放养技术, 形成一种优质高效的水库渔业模式。
[1] Holthuis L B. FAO Species Catalogue. Volume 1. Shrimps and Prawns of the World. An Annotated Catalogue of Species of Interest to Fisheries [M]. FAO Fisheries Synopsis, 1980, 1—271
[2] Li X Z, Liu R Y, Liang X Q, et al. Fauna Sinica. Invertebrata Vol. 44. Crustacea. Decapoda. Palaemonoidea [M]. Beijing: Science Press. 2007, 61—64, 128—131 [李新正, 刘瑞玉, 梁象秋, 等. 中国动物志. 无脊椎动物.第44卷. 十足目. 长臂虾总科. 北京: 科学出版社. 2007, 61—64, 128—131]
[3] Tan Q K, Zhu Q S. Freshwater shrimps of Chaohu Lake and the surrounding waters [J]. Journal of Anhui Normal University (Natural Science), 1982, (3): 80—85 [谈奇坤, 朱清顺. 巢湖及其周围水域的虾类. 安徽师范大学报 (自然科学版), 1982, (3): 80—85]
[4] Tan Q K, Wen Y Q, Dong X Y, et al. The current situation and use of freshwater shrimps in Chaohu Lake [J]. Freshwater Fisheries, 1985, (3): 10—13 [谈奇坤, 温耀群,董秀英, 等. 巢湖虾类资源现状及其利用意见. 淡水渔业,1985, (3): 10—13]
[5] Shi W G, Yan X M, Bing X W. Biology and feeding habit of Exopalaemon modestus (Heller) in Taihu Lake [J]. Journal of Lake Science, 1995, 7(1): 69—76 [施炜纲, 严小梅, 邴旭文. 太湖秀丽白虾生物学与食性的研究. 湖泊科学, 1995, 7(1): 69—76]
[6] Xu J, Zhang M, Xie P. Stable isotope changes in freshwater shrimps (Exopalaemon modestus and Macrobrachium nipponensis): trophic pattern implications [J]. Hydrobiologia, 2008, 605(1): 45—54
[7] Oh C W, Suh H L, Park K Y, et al. Growth and reproductive biology of the freshwater shrimp Exopalaemon modestus (Decapoda: Palaemonidae) in a lake of Korea [J]. Journal of Crustacean Biology, 2002, 22(2): 357—366
[8] Wen Z R, Xie P, Xun J. reproductive biology of Exopalaemon modestus in Taihu Lake [J]. Journal of Yangtze University (Natural Science Edition), 2012, 9(4): 23—40 [温周瑞, 谢平, 徐军. 太湖秀丽白虾繁殖生物学研究. 长江大学学报(自然科学版), 2012, 9(4): 23—40]
[9] Wen Z R, Xie P. Growth of Exopalaemon modestus in Taihu Lake [J]. Fisheries Science, 2013, 40(3): 139—142 [温周瑞,谢平. 太湖秀丽白虾生长研究. 水产科技情报, 2013, 40(3): 139—142]
[10] Lai W, Deng X H, Chen B L, et al. Investigations on the characteristics of population and reproductive habit of fresh water prawn, Exopalaemon modestus Heller in Taihu Lake [J]. Journal of East China Normal University (NaturalScience Edition), 1983, (3): 81—87 [赖伟, 邓雪怀, 陈炳良,等. 太湖秀丽白虾种群及生殖习性研究. 华东师范大学学报(自然科学版), 1983, (3): 81—87]
[11] Zhang L. A stable carbon and nitrogen isotope study of fishes from the Three Gorges riverine section of the Yangtze River [D]. Thesis for Doctor of Science. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan. 2007 [张亮. 长江三峡江段鱼类碳、氮稳定性同位素研究. 博士学位论文, 中国科学院水生生物研究所, 武汉. 2007]
[12] Gayanilo F C, Sparre P, Pauly D. FAO-ICLARM Stock Assessment Tools II (FiSAT II). Revised version. User’s guide [M]. FAO Computerized Information Series (Fisheries). No.8, Revised version. Rome, 2005, 1—168
[13] Pauly D. Theory and Management of Tropical Multiespecies Stocks: A Review, with Emphasis on the Southeast Asian Demersal Fisheries [M]. Manila: International Centor for Living Aquatic Resources Management, 1979, 1—35
[14] Somerton D A. A computer technique for estimating the size of sexual maturity in crabs [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37(10): 1488—1497
[15] Cohen F P A, Takano B F, Shimizu R M, et al. Life cycle and Population structure of Aegla paulensis (Decapoda: Anomura: Aeglidae) [J]. Journal of Crustacean Biology, 2011, 31(3): 389—395
[16] Guerao G, Pérez-Baquera J, Ribera C. Growth and Reproductive Biology of Palaemon xiphias Risso, 1816 (Decapoda: Caridea: Palaemonidae) [J]. Journal of Crustacean Biology, 1994, 14(2): 280—288
[17] Ito M, Watanabe S, Murano M. Growth and reproduction of Palamemon pacificus and P. Serrifer [J]. Nippon Suisan Gakkaishi, 1991, 57(7): 1229—1239 [In Japanese with English summary]
[18] Forster G R. The biology of the common prawn, Leander serratus Pennant [J]. Journal of the Marine Biological Association of the UK, 1951, 30(2): 333—360
[19] Jefferies D J. The moulting behaviour of Palaemonetes varians (Leach) (Decapoda, Palaemonidae) [J]. Hydrobiologia, 1964, 24(3–4): 457—488
[20] Kim S, Hong S. Reproductive biology of Palaemon gravieri (Decapoda: Caridea: Palaemonidae) [J]. Journal of Crustacean Biology, 2004, 24(1): 121—130
[21] Béguer M, Bergé J, Girardin M, et al. Reproductive biology of Palaemon longirostris (Decapoda: Palaemonidae) from Gironde Estuary (France), with a comparison with other european populations [J]. Journal of Crustacean Biology, 2010, 30(2): 175—185
[22] Wear R G. Incubation in British decapod Crustacea, and the effects of temperature on the rate and success of embryonic development [J]. Journal of the Marine Biology Association of the UK, 1974, 54:745—762
[23] Sun J Y, Zhang D Y, Duan Z H, et al. The population reproduction of freshwater shrimp (Macrobrachium nipponense) [J]. Journal of Lake Science, 1998, 10(2): 72—77 [孙建贻, 张道源, 段中华, 等. 洪湖日本沼虾的种群繁殖生物学. 湖泊科学, 1998, 10(2): 72—77]
[24] Berglund A. Sex dimorphism and skewed sex ratios in the prawn species Palaemon adspersus and P. squilla [J]. Oikos, 1981, 36: 158—162
[25] Kim S. Population structure, growth, mortality and size at sexual maturity of Palaemon gravieri (Decapoda: Caridea: Palaemonidae) [J]. Journal of Crustacean Biology, 2005, 25(2): 226—232
[26] Béguer M, Bergé J, Girardin M, et al. Growth modeling and spatio-temporal variability in the body condition of the estuarine shrimp Palaemon longirostris in the gironde (Sw France) [J]. Journal of Crustacean Biology, 2011, 31(4): 606—612
[27] Alexandra C. Growth of the prawn palaemon longirostris (Decapoda, Palaemonidae) in Mira River and Estuary, Sw Portugal [J]. Journal of Crustacean Biology, 2003, 23(2): 251—257
[28] Paschoal L R P, Guimarães F J, Couto E C G. Relative growth and sexual maturity of the freshwater shrimp Palaemon pandaliformis (Crustacea, Palaemonidae) in northeastern of Brazil (Canavieiras, Bahia) [J]. Iheringia. Série Zoologia, 2013, 103(1): 31—36
[29] Sabri B, Osman S, Ozcan O. Seasonal growth and reproduction biology of the Baltic prawn, Palaemon adspersus (Decapoda: Palaemonidae), in the southern Black Sea [J]. Journal of the Marine Biological Association of the UK, 2009, 89(3): 509—519
[30] Wang K, Duan X B, Liu S P, et al. Survey on temporal and spatial distribution of fish in the Daning River [J]. Acta Hydrobiologica Sinica, 2009, 33(3): 516—527 [王珂, 段辛斌, 刘绍平, 等. 三峡库区大宁河鱼类的时空分布特征.水生生物学报, 2009, 33(3): 516—527]
[31] Wu Q, Duan X B, Xu S Y, et al. Studies on fishes resources in the Three Gorges Reservoir of the Yangtze River [J]. Freshwater Fisheries, 2007, 37(2): 70—75 [吴强, 段辛斌,徐树英, 等. 长江三峡库区蓄水后鱼类资源现状. 淡水渔业, 2007, 37(2): 70—75]
[32] Xiao T Y, Zhang H Y, Wang X Q, et al. Biological characteristics of Pelteobagrus fulvidraco in Dongting Lake [J]. Chinese Journal Zoology, 2003, 38(5): 83—88 [肖调义,章怀云, 王晓清, 等. 洞庭湖黄颡鱼生物学特征. 动物学杂志, 2003, 38(5): 83—88]
[33] Li W, Zhang T L, Ye S W, et al. Feeding habits and predator-prey size relationships of mandarin fish Siniperca chuatsi (Basilewsky) in a shallow lake, central China [J]. Journal of Applied Ichthyology, 2012, 29: 56—63
THE GROWTH AND THE REPRODUCTIVE BIOLOGY OF EXOPALAEMON MODESTUS (HELLER, 1862) (CRUSTACEA, DECAPODA, PALAEMONIDAE) IN THE THREE GORGES RESERVOIR OF CHINA
CHEN Si-Bao1,2, LIAO Chuan-Song1,2, ZHAO Xiu-Jiang3, YE Shao-Wen1, LI Zhong-Jie1, ZHANG Tang-Lin1and LIU Jia-Shou1
(1. State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Department of S & T and Enviroment Protection, China Three Gorges Corporation, Beijing 100038, China)
We conducted monthly study on the growth and the reproductive biology of shrimp Exopalaemon modestus in the Three Gorges Reservoir from November 2012 to October 2013. The typical life span of the shrimp was 12—14 months. E. modestus showed a clear sexual dimorphism in the body length, and the females were significantly larger in size than the males. The Von Bertalanffy growth function was estimated to be Lt= 59.33[–e–1.6(t+0.081)] based on the frequency of the body length. The ovigerous females were monitored between early April and late October, and we found that the peak reproductive period was from mid April to mid June. The spawning populations included wintering and yearling individuals. The body lengths of the ovigerous females ranged from 27.07 to 53.71 mm, and the body length of 50% was estimated to be 37.06 mm. The overall sex ratio was 1.46︰1 and females significantly outnumbered males. The absolute fecundity of E. Modestus ranged from 31 to 294 eggs [(127±53) eggs, mean±SD], and the relative fecundity was (115±22) eggs/g (mean ± SD). The absolute fecundity was related to the body length (FA=0.0031L2.8632, r=0.72, n=106) and the body weight (FA=104.63W+9.9534, r=0.79, n=106). The average diameter of the eggs of E. Modestus was (1215±102) μm (mean±SD, n=4171). In order to properly protect and utilize the freshwater shrimp resources, we recommended to apply more strict management policies on shrimp harvest, for example, to reduce or forbid captures during the spawning season (April to September).
Three Gorges Reservoir; Exopalaemon modestus; Growth; Reproductive biology
Q958.8
A
1000-3207(2015)05-0989-08
10.7541/2015.129
2015-03-26;
2015-05-23
国家科技支撑计划课题(2012BAD25B08); 中国长江三峡集团公司科研项目(CT-12-08-01)资助
陈思宝(1989—), 男, 安徽肥东人; 博士研究生; 研究方向为渔业生态学。E-mail: chensb@ihb.ac.cn
刘家寿, E-mail: jsliu@ihb.ac.cn
