基于降低水稻谷蛋白多成员表达水平的干涉表达载体构建
2015-03-02赵丰兰段永波薛建平
赵丰兰,段永波,盛 玮,薛建平
(淮北师范大学生命科学学院/资源植物生物学安徽省重点实验室,安徽 淮北 235000)
基于降低水稻谷蛋白多成员表达水平的干涉表达载体构建
赵丰兰,段永波,盛玮,薛建平
(淮北师范大学生命科学学院/资源植物生物学安徽省重点实验室,安徽淮北235000)
摘要:通过水稻全基因组比对分析,获得16个谷蛋白成员信息。序列比对获得其保守区域,并选取其中129 bp构建RNAi干涉载体,双酶切验证和测序结果表明干涉载体构建成功。
关键词:水稻;谷蛋白;RNAi干涉载体;保守区域
水稻为全球一半以上人口的主食,对保障全球粮食安全极为重要。同时水稻种子也是目前认为最有潜力作为生物反应器生产重组药用蛋白的“工厂”之一[1]。目前,限制水稻种子特异表达体系在重组药用蛋白大规模应用的主要因子是目标基因表达量过低。外源基因表达受到转录、翻译或翻译后加工修饰等多层面的协同调控。已用于提高外源基因表达水平的策略包括使用强特异表达启动子、改善蛋白运输环境、提高翻译效率、特定细胞器储存等[2 -5],都一定程度上提高了外源蛋白表达量。这些方法都是针对目的基因表达过程的直接改造以提高表达水平,该类体系已较成熟,如果再想大幅提高外源蛋白的表达量难度较大。
外源基因在表达过程中涉及与内源相关蛋白对氨基酸和存储空间的直接竞争[6]。2003年,Tada等将转大豆球蛋白(glycinin)基因的常规水稻品种与日本晴的低谷蛋白水稻突变体(LGC-1)进行杂交,LGC-1后代中glycinin表达量比其亲本日本晴的杂交后代提高120%,达到237 μg/g,暗示突变体中谷蛋白含量的降低提供了更多的氨基酸和空间供glycinin合成[7]。Kuroda等设计了一套包括抑制水稻内源蛋白合成和种子特异表达外源目标基因这2个表达框的植物表达载体,结果发现内源蛋白含量降低的转基因水稻种子中绿色荧光蛋白(GFP)表达水平比抑制前显著增强[8]。Kurokawa等筛选到霍乱毒素B亚基表达量显著提高的转基因株系,表明利用外源基因与相关内源基因的竞争关系可提高外源蛋白表达水平,然而关于降低水稻种子内源蛋白含量如何增强外源基因表达尚不清楚[9-10]。
另一方面,水稻种子具有调节各种内源蛋白组成以保持其总蛋白含量于一定范围的动态平衡机制。Maruta等利用RNAi技术降低水稻谷蛋白表达以改良水稻蒸煮品质,结果检测到种子的其他内源储藏蛋白含量增加[11]。Kim等在抑制水稻13 kDa醇溶蛋白合成改进营养品质的研究中发现,该醇溶蛋白显著降低的同时,其他蛋白含量如10 kDa醇溶蛋白、谷蛋白和伴侣蛋白等增加[12]。综合分析多个研究组的研究结果,无论低内源蛋白突变体、RNAi降低内源蛋白的水稻种子,还是以二者作为受体表达外源蛋白的种子,均与其野生型在总蛋白含量上无显著差异,表现为各种内源蛋白或与外源蛋白之间的此消彼长,说明水稻种子各种蛋白组分间存在一种动态平衡机制,使得其总蛋白含量维持于一恒定范围[11-14]。
因此,以水稻中谷蛋白所有成员的保守区域为靶序列,通过RNAi技术降低谷蛋白的含量,可创制低谷蛋白的突变体,为提高外源重组蛋白表达水平奠定基础。
1 材料与方法
1.1材料
日本晴水稻种子由安徽省农业科学院水稻研究所提供。
植物表达载体pCAMBIA1390RNAi骨架载体、大肠杆菌(Escherichia coli)菌株DH5α、根癌农杆菌(Agrobacterium tumefaciens)菌株EHA105为淮北师范大学生命科学学院资源植物生物学安徽省重点实验室课题组保存。
PCR扩增试剂为生工生物工程(上海)股份有限公司产品,限制性内切酶SacI、StuI、BamHI、MluI、T4 DNA连接酶均购自New England BioLabs (NEB)公司,胶回收试剂盒购自AxyGen Biosciences公司,pMD18-T vector购自TaKaRa生物工程(大连)有限公司,卡那霉素、氨苄青霉素钠盐和利福平等购自生工生物工程(上海)股份有限公司,引物和基因合成以及测序由生工生物工程(上海)股份有限公司完成。
1.2方法
1.2.1水稻谷蛋白基因及其保守区域分析通过NCBI软件分析水稻全基因组范围内谷蛋白分布情况,并采用Blast程序对所获成员的保守区域进行分析。选定其中部分保守序列作为靶位点设计引物,构建RNAi干涉表达载体(表1)。

表1 RNAi干涉表达载体构建所用引物序列
1.2.2靶片段的获得及载体构建以日本晴基因组DNA为模板扩增正义和反义靶片段。PCR扩增体系(50μl)如下:5 μl 10×PCR buffer,4 μl 25 mmol MgCl2,4 μl 2.0 mmol dNTPs,2 μl 10 μmol上下游引物,0.4 μl 5 U/μl Taq DNA聚合酶,100 ng DNA模板,ddH2O至50 μl。PCR扩增条件为94℃预变性5 min;94℃30 s,55℃30 s,72℃30 s进行30循环。PCR产物连接至pMD18-T vector,挑取PCR阳性克隆送生工生物工程(上海)股份有限公司测序。
1.2.3RfTNFR-Fc种子特异表达载体构建以SacI/StuI对pCAMBIA1390RNAi骨架载体质粒进行双酶切和含有正义片段的pMD18-T vector载体,回收片段后进行T4DNA酶重组连接;提取所获阳性菌落质粒进行BamHI/MluI双酶切使载体线性化,同理将反义片段连入表达载体。提取阳性克隆质粒以相应内切酶进行正义片段和反义片段的酶切验证,鉴定正确的克隆送生工生物工程(上海)股份有限公司测序。
1.2.4表达载体导入根癌农杆菌将测序验证正确的重组质粒转入根癌农杆菌EHA105感受态细胞。取2 μg质粒DNA加入从-70℃超低温冰箱取出的农杆菌感受态细胞,混匀后冰浴30 min;转入液氮速冻1 min;加入1 ml YEP培养基(不含抗生素),30℃、120 r/min培养4 h;4 000 r/min离心1 min,弃上清;加150 μl YEP培养基(不含抗生素)重悬,将菌液涂布与含50 μg/ml Kan和10μg/ml Amp的YEP固体平板;28℃培养48 h至单菌落长出,进行菌落PCR鉴定。
2 结果与分析
2.1水稻中谷蛋白同源性分析
以谷蛋白基因Os10g0400200全长序列(1540bp)对水稻全基因组数据库进行搜索比对,获得16个谷蛋白基因信息(Os10g0400200、Os01g0762500、Os02g0249800、Os02g0249000、Os02g0249600、Os02g0268300、Os02g0268100、Os02g0248800、Os02g 0242600、Os02g 0453600、Os02g 0249000、Os03g 0427300、Os12g 0155200、Os08g 0127900、Os11g 0153400、Os05g0170300)。通过NCBI Blast软件对这些基因进行全长比对,结果表明其同源性极高(图1)。选取其中部分保守区域作为靶位点,序列为:tttgataggttgcaagcatttgagccaattcggagtgtgaggtctcaagctggcacaactgagttcttcgatgtctctaatgagttgtttcaatgtaccggagtatctgttgtccgccgagttattgaa。以该区域为靶序列可能同时抑制所有谷蛋白基因的表达。
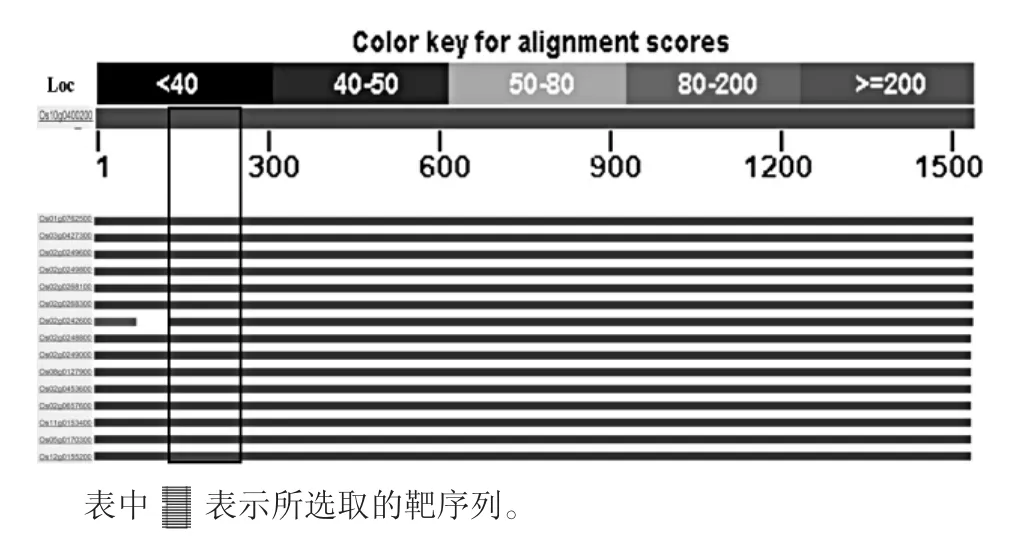
图1 水稻谷蛋白基因的同源性比对
2.2表达载体构建
回收PCR片段后连入T载体,分别酶切后与相应的酶线性化的载体进行连接,对所获重组子进行正义片段和反义片段的酶切验证。酶切结果显示,所获表达载体中含有129 bp的正义和反义片段(图2)。测序结果表明,正义和反义片段已正确连入pCAMBIA1390 RNAi骨架载体。
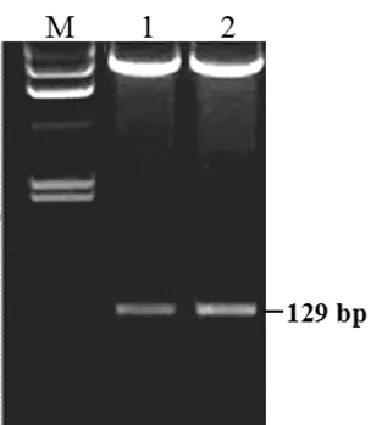
M为genomic DNA marker II;1为正义片段酶切结果;2为反义片段酶切结果。图2重组子正义和反义片段的双酶切验证
2.3重组子导入根癌农杆菌
提取测序正确克隆的质粒DNA转化根癌农杆菌EHA105感受态细胞,液氮速冻和恢复培养后涂于含卡那霉素和利福平的YEP平板。培养2 d后获得大量转化克隆(图3),菌落PCR表明重组质粒已导入根癌农杆菌菌株EHA105。

图3 阳性农杆菌克隆
3 结论
谷蛋白是水稻种子中含量最高的储藏蛋白,其含量占总蛋白的80%左右,包括至少12个编码基因[3]。通过敲除或降低谷蛋白表达,对提高外源基因的表达水平很有意义和潜力。该研究通过水稻全基因组比对分析,获得16个谷蛋白成员信息。序列比对获得其保守区域,并选取其中129 bp构建RNAi干涉载体,双酶切验证和测序结果表明干涉载体构建成功。
参考文献:
[1]OU J Q,GUO Z B,SHI J N,et al. Transgenic rice endosperm as a bioreactor for molecular pharming[J]. Plant Cell Reports,2014,33:585-594.
[2]YANG L,SUZUKI K,HIROSE S,et al. Development of transgenic rice seed accumulating a major Japanese cedar pollen allergen(Cry j1)structurally disrupted for oral immunotherapy[J]. Plant Biotechnol J.,2007,5:815-826.
[3]GREENHAM T,ALTOSAAR I. Molecular strategies to engineer transgenic rice seed compartments for large -scale production of plant -made pharmaceuticals[J]. Methods Mol. Biol.,2013,956:311-326.
[4]BENCHABANE M,GOULET C,RIVARD D,et al. Preventing unintended proteolysis in plant protein biofactories[J]. Plant Biotechnol J.,2008,6(7):633-648.
[5]BOOTHE J,NYKIFORUK C,SHEN Y,et al. Seedbased expression systems for plant molecular farming [J]. Plant Biotechnol J.,2010,(5):588-606.
[6]SHIGEMITSU T,OZAKI S,SAITO Y,et al. Production of human growth hormone in transgenic rice seeds:co -introduction of RNA interference cassette for suppressing the gene expression of endogenous storage pro-teins[J]. Plant Cell Rep.,2012,31:539-549.
[7]TADA Y,UTSUMI S,TAKAIWA F. Foreign gene products can be enhanced by introduction into low storage protein mutants[J]. Plant Biotechnol J.,2003,1:411-422.
[8]KURODA M,KIMIZU M,MIKAMI C. A simple set of plasmids for the production of transgenic plants[J]. Biosci. Biotechnol. Biochem.,2010,74(11):2 348-2 351.
[9]TOKUHARA D,YUKI Y,NOCHI T,et al. Secretory IgA-mediated protection against V. cholerae and heatlabile enterotoxin-producing enterotoxigenic Escherichia coli by rice-based vaccine[J]. PNAS,2010,107:8 794-8 799.
[10]KUROKAWA S,KURODA M,MEJIMA M,et al. RNAi-mediated suppression of endogenous storage proteins leads to a change in localization of overexpressed cholera toxin B-subunit and the allergen protein RAG2 in rice seeds[J]. Plant Cell Rep.,2014,33:75-87.
[11]MARUTA Y,UEKI J,SATO H,et al. Transgenic rice with reduced glutelin content by transformation with glutelin A antisense gene[J]. Mol. Breed,2001,8:273-284.
[12]KIM H J,LEE J Y,YOON U H,et al. Effects of reduced prolamin on seed storage protein composition and the nutritional quality of rice[J]. Int. J. Mol. Sci.,2013,14:17 073-17 084.
[13]KUSABA M,MIYAHARA K,IIDA S,et al. Low glutelin content1:A dominant mutation that suppresses the glutelin multigene family via RNA silencing in rice [J]. Plant Cell,2003,15:1 455-1 467.
[14]KAWAKATSU T,HIROSE S,YASUDA H,et al. Reducing rice seed storage protein accumulation leads to changes in nutrient quality and storage organelle formation[J]. Plant Physiol.,2010,154:1 842-1 854.
(本文责编:金苹)
通讯作者:薛建平(1966—),男,河南温县人,教授,博士,主要从事植物生物技术研究。E-mail:xuejp@163.com
作者简介:赵丰兰(1979—),女,安徽寿县人,讲师,硕士,主要从事植物生物技术研究。E-mail:zhaofenglan1997@163. com
基金项目:安徽省教育厅省级高校自然科学基金重点项目(KJ2014A226)
收稿日期:2015-07-11
doi:10.3969/j.issn.1001-1463.2015.09.011
中图分类号:S511
文献标识码:A
文章编号:1001-1463(2015)09-0028-04
