温度驯化对白腰朱顶雀脱支酶活性的影响1)
2015-02-08王有祥徐兴军邵淑丽李怀永
王有祥 徐兴军 邵淑丽 李怀永
(齐齐哈尔大学,齐齐哈尔,161006) (齐齐哈尔市第一医院放疗科)
吕建伟 王维禹 张伟伟 谢志刚 李旭艳
(齐齐哈尔大学)
温度驯化对白腰朱顶雀脱支酶活性的影响1)
王有祥 徐兴军 邵淑丽 李怀永
(齐齐哈尔大学,齐齐哈尔,161006) (齐齐哈尔市第一医院放疗科)
吕建伟 王维禹 张伟伟 谢志刚 李旭艳
(齐齐哈尔大学)
为探讨温度对白腰朱顶雀(Carduelisflammea)生理生化指标的影响,以白腰朱顶雀为研究对象,将其分为低温组(5 ℃)、高温组(30 ℃)和对照组(23 ℃)进行驯化,驯化后测定其体质量、体脂质量分数(索氏抽提法)、器官的鲜质量和干质量(烘干恒质量法)、基础代谢率和脱支酶活性。结果表明:低温组体质量降低,在驯化0~2周内下降最为明显,第2周结束时与对照组差异极显著(p<0.01);高温组体质量增高,在驯化1~3周内涨幅较大,第1周结束时与对照组差异显著(p<0.05)。低温组基础代谢率显著高于高温组(p<0.05)。3组动物肌肉中脱支酶活性差异不显著(p>0.05);低温组肝脏中脱支酶活性与对照组差异极显著(p<0.01),高温组肝脏中脱支酶活性与对照组差异不显著(p>0.05),低温组肝脏中脱支酶活性与高温组差异极显著(p<0.01)。可见,温度是影响白腰朱顶雀产热特征改变的主要环境因子之一。低温条件下,白腰朱顶雀通过降低体质量、提高基础代谢率、增加摄入能来补偿能量消耗等一系列的生理生化反应,以适应外界环境温度的变化。
白腰朱顶雀;脱支酶;温度驯化;基础代谢率
In order to study the effect of the temperature on physiological and biochemical indexes ofCarduelisflammea, the experimental animals were randomly divided into low-temperature group (5 ℃), high-temperature group (35 ℃) and the control group (23 ℃), and after the domestication, the body weight, the body fat (by soxhlet extraction method), the fresh weight, dry weight (drying constant weight method) of organs, basal metabolic rate and debranching enzyme activity were determined. The low-temperature group weight reduced, and during the domestication within 0-2 weeks, it was declined significantly with extremely significant difference at the end of second week compared with control group (p<0.01). The high-temperature group weight increased,during the domestication within 1-3 weeks, it was increased significantly with extremely significant difference at the end of first week compared with control group (p<0.05). Low-temperature group basal metabolic rate was significantly higher than that in high-temperature group (p<0.05). Three groups of animal muscle debranching enzyme activity was not significantly different (p>0.05). Low-temperature group debranching enzyme activity in the liver was significantly different compared with the control group (p<0.01), but for the difference between high-temperature group debranching enzyme activity in the liver compared with the control group, there was no significant difference (p>0.05). The difference between the low-temperature group debranching enzyme activity in the liver that in high-temperature group was significant (p<0.01). Therefore, temperature is one of the major environmental factors affectingC.flammeathermogenesis. Under low temperature,C.flammeacan reduce body weight, improve the metabolic rate, and increase energy intake to compensate the energy consumption to adapt the temperature changes of external environment.
动物通过采取适当的体温、体质量、产热能力和能量代谢等方面的调节来应对恶劣环境。在季节环境变化过程中,动物的表型可塑性变化对其生存至关重要[1-2]。温度变化被认为是影响鸟类能量代谢及产热能力的重要因素。目前,国内外许多学者对温度影响鸟类能量代谢及产热方面进行了研究。在国外,Wilson et al.[3]在研究电子温度对美国红隼(Falcotinnunculus)能量代谢的影响中取得一定进展。Prinzinger et al.[4]在研究温度波动对日本鹌鹑(Coturnixjaponica)气体代谢的影响中发现短期内的温度变化,不会对基础代谢率产生影响。在国内,徐兴军等[5]在研究pH值和温度对春季麻雀(Passermontanus)体内消化酶活力的影响中发现腺胃、肌胃和胰腺蛋白酶活力最适温度较低,在肠道和肝脏中温度较高。糖原作为重要的储能形式,对动物获得生存所需要的能量具有重要意义[6]。糖原在分解过程中产生许多种酶,脱支酶是其中一种关键酶,它催化糖原脱支的2个反应[7-8]。
白腰朱顶雀(Carduelisflammea)属雀形目(Passeriformes)雀科(Fringillidae),生活于溪边丛生柳林、沼泽化的多草疏林,食性比较杂,随着季节的变化而不同,春夏两季主要以食虫为主,秋冬两季主要以草籽为主。常见于我国东北地区,是中国北方地区最常见的雀形目之一。国内关于白腰朱顶雀的研究主要集中在基础产热和基础代谢率的测定方面,对脱支酶活性的研究相对较少。本研究测定了白腰朱顶雀在高温和低温条件驯化下,其体质量、器官质量、基础代谢率、体脂质量分数和脱支酶活性的可塑性变化,旨在深入了解动物适应环境温度变化条件下的能量学策略,阐明产热和体质量调节在动物适应冷驯化和热驯化环境中的作用和意义,为进一步了解北方小型鸟类的生存机制提供理论依据。
1 材料与方法
1.1 试验动物
试验用白腰朱顶雀捕自黑龙江省齐齐哈尔市扎龙国家自然保护区(123°47′~124°37′E,46°52′~47°32′N),于齐齐哈尔大学生命科学与农林学院动物学实验室饲养,自由取食及饮水,室温(22±1)℃,自然光照。适应饲养1周后,按体质量随机分为低温组8只(5 ℃光照培养箱)、高温组8只(30 ℃光照培养箱)、对照组8只(23 ℃)进行驯化,组间体质量无明显差异。
1.2 试验仪器
电子天平(BS210型)Sartorius公司;台式高速冷冻离心机(TGL-16M)HERMLE Z323K Germany;高压灭菌锅LS-C50L浙江;超低温冰箱Μ-6382E Japan(-86 ℃);IKA氧弹量热仪C2000 Germany;封闭式流体压力呼吸仪,北京师范大学司南仪器厂;可见分光光度计(723N),上海精密科学仪器有限公司;智能光照培养箱GZP-250A,南京红龙仪器设备厂;数显光照培养箱250D,江苏省金坛市荣华仪器制造有限公司。
1.3 体质量及器官质量的测定
体质量和各器官组织鲜质量、干质量用电子天平(BS210型)测量,精确到0.01 g。驯化试验开始后,分别测定对照组及各试验组白腰朱顶雀初体质量,驯化试验结束时,测定各试验组白腰朱顶雀终体质量及主要器官质量。
1.4 基础代谢率的测定
基础代谢率是维持动物正常生理反应和机体机制的最小产生热量的速率,是动物维持身体各部分基本功能所需要的最小能量值[9-12]。试验采取封闭式流体压力呼吸仪测定白腰朱顶雀的基础代谢率,试验动物称量体质量精确至0.01 g,水浴温度控制在(25±0.5)℃,呼吸室体积为3.6 L,用氢氧化钠溶液和二氧化硅来吸收呼吸室内的二氧化碳和水,每隔3 d测1次基础代谢率。测量前先将白腰朱顶雀禁食4 h,进行称质量,然后将其放入呼吸室内适应30 min,待白腰朱顶雀呼吸稳定后开始进行测量,每隔5 min记录1次,连续测定1 h,选取比较稳定的2个连续的最低值,计算BMR。BMR计算公式为,μ0=μt×H×12/(1+apt)×760×W。式中:μ0为0 ℃和760 mmHg(≈101.3 kPa)大气压时的气体体积;μt为动物在1 h内所消耗的氧量;apt为气体在气压不变时某一温度下的扩张系数;H为试验时的气压;W为动物体质量。试验中标准大气压为101.325 kPa,H/760=0.977 6,1+apt=1.091 7。
1.5 脱支酶活性的测定
测定脱支酶的最适温度为50 ℃、最适pH为6.0、最适底物浓度为1%。将300 μL 1%支链淀粉溶液和100 μL上清酶液,加入100 μL乙酸钠缓冲溶液(pH=6.0、0.5 mol·L-1)。在50 ℃水浴中反应1 h,取出100 μL反应液,加入2.5 mL 0.01 mol·L-1的稀碘液后迅速放入分光光度计(620 nm)测定OD值,以未反应的酶液和底物混合物作为空白。在最适条件下作用1 h,每分钟生成支链淀粉造成620 nm下吸光值增加0.01的酶活定义为一个酶活单位。
酶活力(U)=ΔOD×V×N×100/t×直径长度×M。式中:ΔOD表示待测管与对照管OD的差值;V为液体的体积(L);N为稀释倍数;t为反应时间(1 h);M为样品质量(g);直径长度为1 cm。
1.6 摄入热量值的测定
采取全收粪法测定[13-14]。测出动物在一定期间内食入饲料物质的质量与粪便中排出物质的质量。正确记录动物在某阶段的饲料采食量和全部排粪量,对饲料和粪便进行热量测定。
摄入能=摄入干物质质量×食物热值;
粪便能=粪便干质量×粪便热值;
消化能=摄入能-粪便能。
1.7 体脂质量分数的测定
采取索氏抽提法测定[15]。体脂质量分数=((m1-m2)/m)×100%。式中:m1为胴体干质量;m2为抽提后不含体脂的胴体干质量;m为个体体质量。
1.8 数据统计
利用SPSS软件进行数据统计分析。体质量、器官的鲜质量和干质量、基础代谢率、体脂质量分数、脱支酶活性的参数采用一元方差检测组间差异;驯化因素对体质量、热量值、基础代谢率等参数的影响,采用多重比较LSD进行检验。
2 结果与分析
2.1 体质量
驯化开始时3组白腰朱顶雀体质量组间无明显差异,随着驯化时间的延长,1周后3组白腰朱顶雀体质量差异显著(p<0.05)。高温组白腰朱顶雀体质量在驯化1~3周内大幅度增加,在第4周驯化结束后达到最大值,与初始体质量相比增加0.83 g;低温组白腰朱顶雀体质量在驯化0~2周内急剧下降,在第4周驯化结束后达到最小值,与初始体质量相比减少0.77 g;而对照组白腰朱顶雀体质量增加幅度较小,在第4周驯化结束后达到最高值,与初始体质量相比增加0.78 g(表1)。
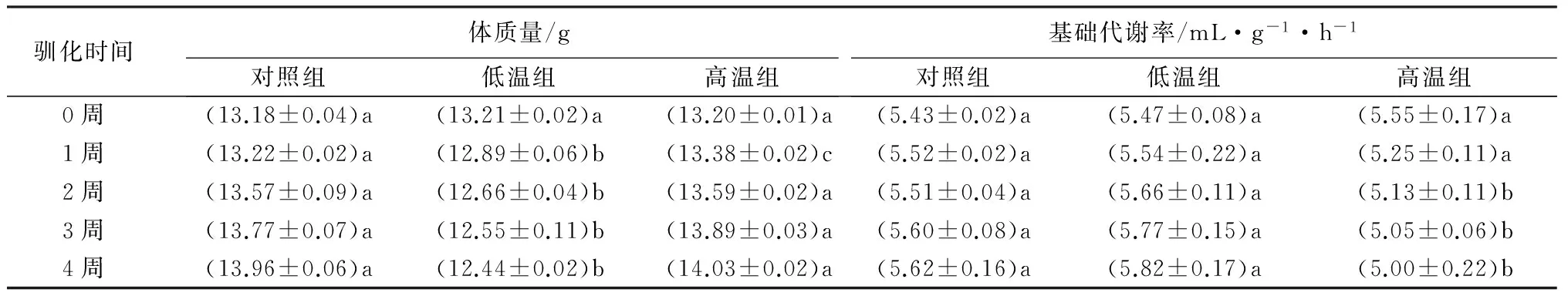
表1 不同温度驯化过程中白腰朱顶雀的体质量与基础代谢率
2.2 基础代谢率(BMR)
在高温驯化过程中,驯化时间对BMR有显著地影响,随着驯化时间的延长,高温组白腰朱顶雀的BMR逐渐降低,在第4周驯化结束后达到最小值,与第1周相比减小0.25 mL·g-1·h-1;低温组白腰朱顶雀的BMR逐渐升高,在1~3周内BMR大幅增加,在第4周驯化结束后达到最大值,与第1周相比增加0.28 mL·g-1·h-1(表1)。
2.3 体脂质量分数
低温组白腰朱顶雀体脂质量分数为10.07%,对照组为11.64%,低温组与对照组间差异显著(p<0.05),高温组白腰朱顶雀体脂质量分数为12.39%,与对照组间差异不显著(p>0.05),低温组体脂质量分数与高温组间差异极显著(p<0.01)。高温组体脂质量分数分别比低温组、对照组提高了2.32%和0.75%。
2.4 消化能
高温组白腰朱顶雀的消化能随着驯化时间的延长而降低,而低温组的消化能随着驯化时间的延长反而升高。低温组、高温组、对照组的消化能分别为57 162.46、31 497.22、51 496.45 J·g-1。低温组、高温组的消化能与对照组间差异极显著(p<0.01)。低温组的消化能分别比高温组、对照组高25 665.24、5 666.01 J·g-1。
2.5 器官鲜质量
低温组、高温组白腰朱顶雀胰脏鲜质量、肾脏鲜质量与对照组间差异极显著(p<0.01),高温组脾脏鲜质量、心脏鲜质量与对照组间差异显著(p<0.05),低温组肝脏鲜质量、胃鲜质量和肠道鲜质量与对照组间差异极显著(p<0.01),高温组肝脏鲜质量、胃鲜质量、肠道鲜质量与对照组相比组间差异不显著(p>0.05)(表2)。
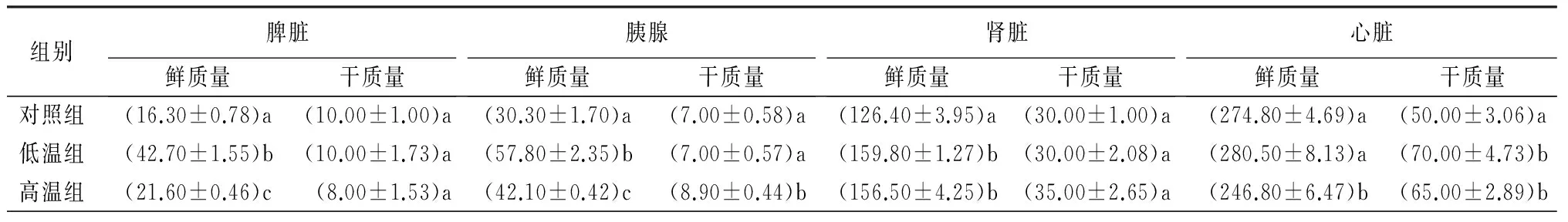
表2 不同温度驯化过程中白腰朱顶雀的器官质量 mg

注:表中数据为平均值±标准误。同列数据后不同字母表示组间差异显著(p<0.05)。
2.6 器官干质量
高温组白腰朱顶雀胰腺干质量、心脏干质量与对照组间差异显著(p<0.05),高温组、低温组脾脏干质量、肾脏干质量、胃干质量、肠道干质量与对照组间差异不显著(p>0.05),低温组心脏干质量、肝脏干质量与对照组间差异极显著(p<0.01),低温组肝脏干质量与高温组差异极显著(p<0.01)(表2)。
2.7 脱支酶活力
低温组、高温组白腰朱顶雀肌肉中脱支酶活性与对照组间无显著差异(p>0.05);低温组肝脏中脱支酶活性与对照组间差异极显著(p<0.01),高温组肝脏中脱支酶活性与对照组间差异不显著(p>0.05),低温组肝脏中脱支酶活性与高温组间差异极显著(p<0.01)(表3)。
表3 不同温度驯化过程中白腰朱顶雀的脱支酶活性
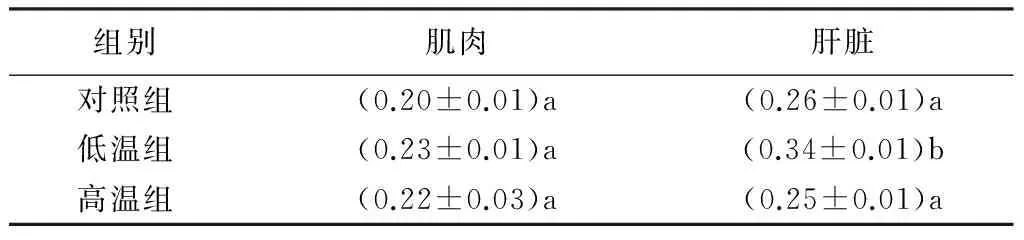
U·g-1
3 结论与讨论
温度是影响鸟类能量代谢及产热能力的重要因素,对鸟类许多生理指标产生重要影响。动物的体质量主要取决于能量收支平衡,体质量的变化反映其营养状态和受环境变化影响进行的一些适应性调节[16-18]。低温组白腰朱顶雀驯化后体质量与驯化前相比降低0.77 g,第4周驯化结束时达到最低值,伴随着体质量的降低呼吸代谢率不断增加,表明白腰朱顶雀通过降低体质量减少总的能量需求来应对低温环境,这可能是由于冷刺激引起机体不自主颤栗,使皮肤血管收缩,血流量减少,机体耗氧量增加进而导致产热量增加。有研究表明,鸟类体脂质量分数的变化与体质量的变化呈正相关[19-20]。由于低温时摄入的能量不足以维持能量需求而通过体内能量储备的转化来补充能量,所以体质量降低的同时体脂质量分数降低。动物在低温环境或生理压力影响下,需要大量的能量来为机体供能。本研究发现白腰朱顶雀低温组的消化能较对照组与高温组高,表明低温是促使白腰朱顶雀能量摄入表现出较高水平的重要原因。这在林琳等[21]的研究中得到了证实。动物的基础代谢率是肝脏、心脏及肾脏等代谢活性器官及肌肉组织等代谢率的总和[22-23]。本试验结果表明,低温组白腰朱顶雀肝脏质量、心脏质量、肾脏质量、胰脏质量、肠道质量较对照组与高温组要高。代谢器官质量发生较小的改变会对基础代谢率产生较大的影响[24]。低温组白腰朱顶雀在第4周驯化结束时基础代谢率达到最高,这与Klaassen et al.[22]研究不同温度对莺基础代谢率及体质量影响的变化结果相一致,提示鸟类通过增加器官质量以适应寒冷的环境条件。高温组白腰朱顶雀驯化后体质量比驯化前增加0.83 g,在第4周驯化结束后达到最大值,这表明白腰朱顶雀体质量具有较强的可塑性,会随着外界环境的变化作出相适应的调节[25]。体质量增加的同时伴随体脂质量分数的升高。脂肪是身体中颤抖性产热的主要的能源物质,高温时动物保留了更多能量用于身体组织的构建和身体机能的恢复。高温环境中,白腰朱顶雀呼吸减慢,基础代谢率降低,同时消化能降低,表明机体通过减少能量的摄入来应对高温环境。
脱支酶是肝糖原、肌糖原分解过程中的一种关键酶。脱支酶活性的高低与糖原含量和分解速度有关[26]。肝糖原分解主要是维持血糖浓度而肌糖原分解主要是为肌肉自身收缩提供能量。当机体受到低温环境影响时,血糖浓度下降促使肾上腺素和胰高血糖素分泌增加,这2种激素与肝脏、肌肉等组织的细胞膜受体结合,经过一系列反应使无活性的磷酸化酶激酶活化为有活性的糖原磷酸化酶抑制糖原生成,促进糖原加速分解,使肝糖原分解为葡萄糖进入血液使血糖浓度升高,为机体提供能量。白腰朱顶雀肝脏中脱支酶活性比肌肉中脱支酶活性高,原因可能是肝糖原可直接分解供能,而肌糖原需要转化为肝糖原后才能分解供能。低温组白腰朱顶雀肝脏中脱支酶活性极显著高于对照组和高温组(p<0.01),肌肉中脱支酶活性差异不显著(p>0.05),而肌肉中脱支酶活性明显低于肝脏中脱支酶活性。原因可能是机体依靠肝糖原直接分解成葡萄糖提供能量,但随着驯化时间的延长,肝糖原不足以维持机体所需的能量,肌糖原经过机体内一系列反应再参与供能,这与徐兴军等[27]在研究温度对麻雀肝糖原和肌糖原含量的影响中得出的结论相一致。有研究表明,冬季麻雀每克肝组织酶活力是夏季的1.6倍[27],也证实了本试验研究结果。
综上所述,在季节性环境变化中,温度是影响白腰朱顶雀产热特征改变的主要环境因子之一,白腰朱顶雀在温度交替变化过程中,通过调整体质量、基础代谢率、消化能等一系列的生理生化反应来适应外界环境温度的变化。低温条件下,通过降低体质量、增加摄入能来补偿能量的消耗,这些适应机制是北方小型鸟类应对低温寒冷环境的生存策略之一。
[1] Corp N, Gorman M L, Speakman J R. Seasonal variation in the resting metabolic rate of male wood miceApodemussylvaticusfrom two contrasting habitats 15 km apart[J]. Journal of Comparative Physiology B,1997,167(3):229-239.
[2] Ashton K G. Patterns of within-species body size variation of birds: strong evidence for Bergmann’s rule[J]. Global Ecology and Biogeography,2002,11(6):505-523.
[3] Wilson G R, Cooper S J, Gessaman J A. The effects of temperature and artificial rain on the metabolism of American kestrels (Falcosparverius)[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology,2004,139(3):389-394.
[4] Prinzinger R. The energy costs of temperature regulation in birds: the influence of quick sinusoidal temperature fluctuations on the gaseous metabolism of the Japanese quail (Coturnixcoturnixjaponica)[J]. Comparative Biochemistry and Physiology Part A: Physiology,1982,71(3):469-472.
[5] 徐兴军,邵淑丽,张伟伟,等.pH值和温度对春季麻雀体内消化酶活力的影响[J].黑龙江畜牧兽医,2012(21):143-145.
[6] 徐兴军,邵淑丽,张伟伟,等.饥饿条件下麻雀体内糖原含量和基础代谢率的变化[J].江苏农业科学,2012,40(11):348-350.
[7] Narahara E, Makino Y, Omichi K. Glycogen debranching enzyme in bovine brain[J]. Journal of Biochemistry,2001,130(3):465-470.
[8] Duan X, Wu J. Advances in studying microbial GH13 starch debranching enzyme[J]. Acta Microbiologica Sinica,2013,53(7):648-656.
[9] Liknes E T, Scott S M, Swanson D L. Seasonal acclimatization in the American Goldfinch revisited: to what extent do metabolic rates vary seasonally?[J]. The Condor,2002,548(104):548-557.
[10] 张国凯,方媛媛,姜雪华,等.白头鹎的代谢率与器官重量在季节驯化中的可塑性变化[J].动物学杂志,2008,43(4):13-19.
[11] 郑蔚虹,方媛媛,姜雪华,等.白头鹎肝脏和肌肉冬夏两季的代谢产热特征比较[J].动物学研究,2010,31(3):319-327.
[12] Montgomery M K, Hulbert A J, Buttemer W A. Metabolic rate and membrane fatty acid composition in birds: a comparison between long-living parrots and short-living fowl[J]. Journal of Comparative Physiology B,2012,182(1):127-137.
[13] 汪诗平,李永宏.放牧率和放牧时期对绵羊排粪量、采食量和干物质消化率关系的影响[J].动物营养学报,1997,9(1):47-54.
[14] 倪小英,林琳,周菲菲,等.光周期对白头鹎体重、器官重量和能量代谢的影响[J].生态学报,2011,31(6):1703-1713.
[15] AL-Mansour M I. Seasonal variation in basal metabolic rate and body composition within individual sanderling bird Calidris alba[J]. J Biol Sci,2004,4:564-567.
[16] Haim A, Zisapel N. Daily rhythms of nonshivering thermogenesis in common spiny miceAcomyscahirinusunder short and long photoperiods[J]. Journal of Thermal Biology,1999,24(5):455-459.
[17] Génin F, Perret M. Photoperiod-induced changes in energy balance in gray mouse lemurs[J]. Physiology & Behavior,2000,71(3/4):315-321.
[18] 赵志军,陈竞峰,王德华.光周期和高脂食物对布氏田鼠能量代谢和产热的影响[J].动物学报,2008,54(4):576-589.
[19] Cooper S J, Swanson D L. Seasonal acclimatization of thermoregulation in the black-capped chickadee[J]. Condor,1994,96(3):638-646.
[20] Pekka T, Rintamäkt P T, Stone J R, et al. Seasonal and diurnal body-mass fluctuations for two nonhoarding species ofParusin Sweden modeled using path analysis[J]. The Auk,2003,120(3):658-668.
[21] 林琳,曹梦婷,胡益林,等.环境温度对白头鹎代谢产热和蒸发失水的影响[J].生态学报,2014,34(3):564-571.
[22] Klaassen M, Oltrogge M, Trost L. Basal metabolic rate, food intake, and body mass in cold-and warm-acclimated Garden Warblers[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology,2004,137(4):639-647.
[23] 王德华,杨明,刘全生,等.小型哺乳动物生理生态学研究与进化思想[J].兽类学报,2009,29(4):343-351.
[24] Williams J B, Tieleman B I. Flexibility in basal metabolic rate and evaporative water loss among hoopoe larks exposed to different environmental temperatures[J]. Journal of Experimental Biology,2000,203(20):3153-3159.
[25] 章迪,周庆宏,何丽娟,等.大绒鼠在不同驯化温度下产热和能量代谢的变化[J].兽类学报,2013,33(4):344-351.
[26] Shen J J, Chen Y T. Molecular characterization of glycogen storage disease type III[J]. Current Molecular Medicine,2002,2(2):167-175.
[27] 徐兴军,王昌河,张伟伟,等.温度对麻雀肝糖原和肌糖原含量的影响[J].黑龙江畜牧兽医,2013(6):164-166.
Effects of Temperature Acclimatization on Enzyme Activity inCarduelisflammeaDebranching//
Wang Youxiang, Xu Xingjun, Shao Shuli
(Qiqihar University, Qiqihar 161006, P. R. China); Li Huaiyong(Department of Radiation Oncology, First Hospital of Qiqihar); Lü Jianwei, Wang Weiyu, Zhang Weiwei, Xie Zhigang, Li Xuyan(Qiqihar University)//Journal of Northeast Forestry University,2015,43(9):107-110,120.
Carduelisflammea; Debranching enzyme; Temperature domestication; Basal metabolic rate
王有祥,男,1990年5月生,齐齐哈尔大学生命科学与农林学院,硕士研究生。E-mail:422645033@qq.com。
徐兴军,齐齐哈尔大学生命科学与农林学院,教授。E-mail:xxj0605@163.com。
2015年5月21日。
Q959.7+39
1)黑龙江省教育厅科学技术研究项目(12541885);齐齐哈尔大学研究生创新科研项目(YJSCX2014-020X)。
责任编辑:程 红。
