牦牛10个STR基因座遗传多态性研究
2015-01-20杨启林徐尚荣王志强
杨启林,张 君,徐尚荣,彭 巍,王志强
(1.青海大学,青海西宁810016;2.青海大学畜牧兽医科学院,青海西宁810016)
我国牦牛的数量约占世界牦牛总数的95%以上[1],多年以来牦牛的育种工作由于各种原因开展不顺利,缺乏可靠稳定的分子遗传标记是重要的原因之一。微卫星标记(microsatellite),又称为短串联重复序列,是以2~6个核苷酸为基本重复单位的串联重复序列[2-3]。微卫星作为第二代分子标记,以其较高的多态性及简便的操作,能够满足一般实验室对遗传研究的要求,而得到较为广泛的应用[4-5],现已被用于种群遗传结构分析、系统发生关系分析及保种效果检测等方面[6-8]。目前将牦牛自身基因组微卫星标记应用于牦牛群体遗传的报道较少[9],研究人员只能利用普通牛的微卫星序列分析牦牛的群体遗传结构特征和系统分类[10-13]。
本研究选用联合国粮农组织(FAO)和国际动物遗传学会(ISAG)联合推荐的用于牛遗传分析的9个Holstein奶牛微卫星基因座和自选的一个微卫星基因座对68个牦牛样本的遗传多样性指征进行分析,旨在初步揭示牦牛核基因组的遗传多样性水平。
1 材料与方法
1.1 试验材料
抽取68 份牦牛血液样本,用EDTA 抗凝后-20 ℃冷冻保存。
1.2 主要试剂与仪器
10×PCR buffer、dNTP、Marker、核酸染料、均购于上海生工生物工程有限公司;高速离心机购自上海HEAL FORCE 公司;PCR 仪(My-Cycle TM Thermal Cycler)购自美国BTO-RAD公司。
1.2 试验方法
1.2.1 基因组DNA 抽提 采用TIANamp Blood DNA Kit试剂盒(天根生化科技(北京)有限公司),按照说明书操作进行,-20 ℃冷冻保存。
1.2.2 PCR 反应体系及反应条件 PCR 扩增反应体系:PCR 体系总体积为25μL,2×PCR Master缓冲液12.5μL,上下引物各0.6μL,DNA 模板1 μL,超纯水10.5μL,采用PCR 仪进行扩增。各STR 基因座的扩增条件为:94 ℃变性10min,退火30s,72 ℃延伸45s,共扩增35~40 个循环,最后72 ℃延伸5 min,4 ℃保存。PCR 产物经2%的琼脂糖凝胶检测后,送往北京优博兰基因技术公司进行基因分型,得到微卫星位点基因型并建立数据库。
1.2.3 微卫星标记与引物 采用了10个微卫星基因座用于牦牛微卫星的基因座筛选,其中9个基因座(ETH10、ETH225、BM1818、BM1824、IL-STS006、CSSM66、HEL9、TGLA53、TGLA126)是根据联合国粮农组织(FAO)和国际动物遗传学会(ISAG)联合推荐的用于牛遗传分析的Holstein奶牛微卫星基因座,另外一个基因座(ILST0013)是根据相关文献提供的微卫星,由上海生工生物公司合成10对引物,加入FAM 荧光标记,引物序列见表1。

表1 所用STR 基因座及引物序列Table 1 Objective STR and their primer sequences
1.2.4 统计分析 用Cervus2.0利用基因型数据库,通过模拟亲子关系及对父母本相似性的分析得到每个位点的等位基因频率、杂合度、多态信息含量(PIC)等遗传学指标[14],同时进行Hardy-Weinberg平衡检验。多态性能够反映物种的进化历史,群体中频率最高的等位基因是该物种中最原始、最保守的基因座。期望杂合度是理论计算得出的杂合度,观望杂合度是随机抽取的两个样本的等位基因不相同的频率。
2 结果与分析
2.1 不同微卫星位点的基因分型
采用荧光标记技术对微卫星PCR 产物进行分析,图1~3显示了部分位点的荧光信号检测等位基因片段大小的自动判别结果。INRA037位点的等位基因大小分别为124.06和132.42bp(图1);ILST0013位点的等位基因大小分别为121.53 和127.29bp(图2);TGLA53位点的等位基因大小分别为153.80和155.86bp(图3)。
2.2 等位基因个数及频率
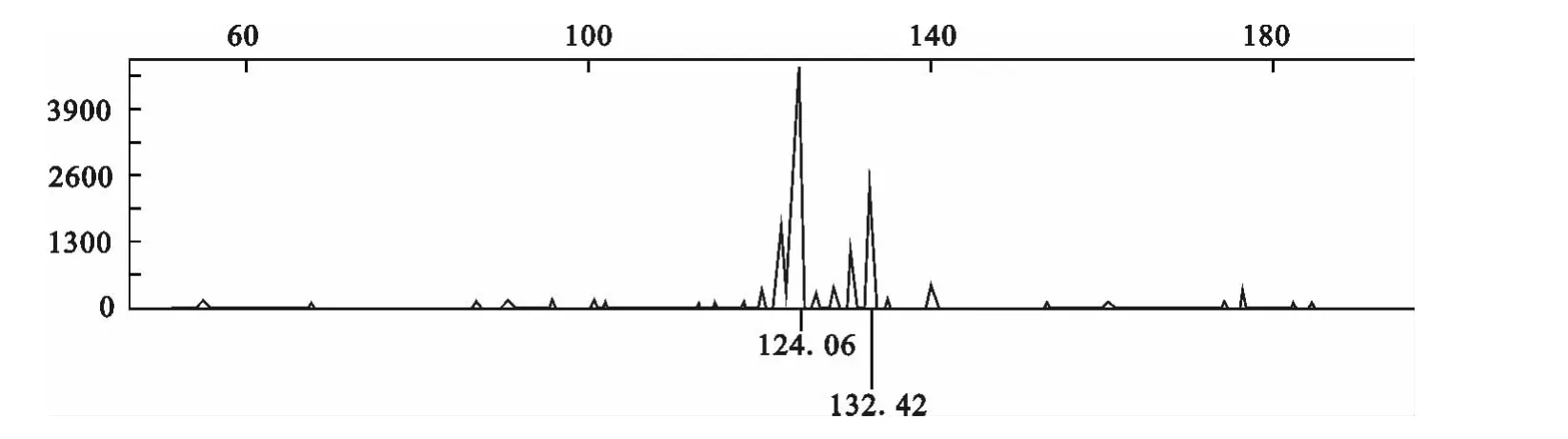
图1 INRA037位点的扩增产物电泳信号Fig.1 Electrophoreses signal about amplification products of INRA037site
分析68头牦牛10个STR 基因座等位基因个数及频率分布规律。由表2可知,TGLA53位点发现有16个等位基因,基因片段大小在151~273bp之间,该等位基因的基因频率范围在0.0074~0.2426。INRA037位点有14 个等位基因,基因片段大小在119~134bp之间,该等位基因的基因频率范围在0.0074~0.3971。ILST0013 位点有11个等位基因,基因片段为117~138bp。其等位基因的基因频率范围为0.0109~0.4485。ETH225、ETH10、BM1824、CSSM66 四个位点有10 个等位基因,基因片段大小分别为133~150bp、209~236 bp、178~263bp、173~190bp。各等位基因的基因频率范围为0.0075~0.2612、0.0074~0.2941、0.0161~0.5597、0.0100~0.3235。BM1818 位点等位基因片段大小为252~260bp,基因频率为0.0278~0.4722;TGLA126 位点等位基因片段大小为119~134 bp,等位基因频率为0.0100~0.3300;LSTS006位点等位基因片段大小为27~291bp,等位基因频率为0.0217~0.4348。
2.3 Hardy-Weinberg平衡检验
利用Cervus 2.0对10个微卫星位点的Hardy-Weinberg 进行平衡检验[15],检测结果显示:BM1824、ILSTS006呈极显著偏离平衡状态(P<0.01),TGLA53、BM1818检测未完成。
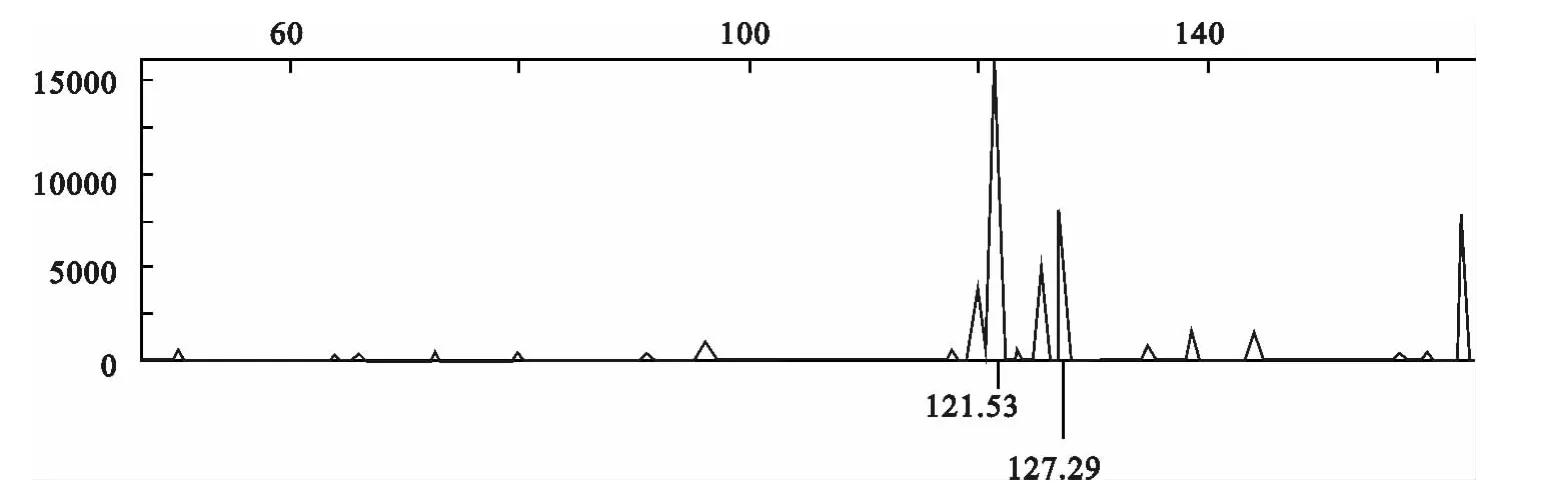
图2 ILST0013位点的扩增产物电泳信号Fig.2 Electrophoreses signal about amplification products of ILST0013site

图3 TGLA53位点的扩增产物电泳信号Fig.3 Electrophoreses signal about amplification products of TGLA53site
2.4 杂合度和多态信息含量
由表3 可见,10 个微卫星位点的PIC 值在0.367~0.849之间变动,平均值为0.685。平均多态信息含量在0.5以上,均为高度多态性标记,表明基因变异较大,遗传多样性丰富;同时也表明本研究所选用的微卫星标记均能提供较好的遗传信息,可以反映牦牛的遗传多样性。牦牛的不同微卫星位点的观察杂合度平均为0.7995;期望杂合度的变动范围为0.333~0.868,平均期望杂合度为0.6154,属于高度杂合标记和高度杂合种群,较高的微卫星标记杂合度表明该标记所能提供的遗传信息量大,较高的种群杂合度表明种群的遗传多样性丰富[16]。
3 讨论
3.1 牦牛种群遗传多样性
微卫星标记是反映DNA 结构多样性的分子遗传标记之一。一个群体或物种的遗传多样性越高或遗传变异越丰富,其对环境变化的适应能力就越强,越容易扩展其分布范围和适应新的环境。遗传杂合度反映了一个群体中某个等位基因被抽取的前提下,其他等位基因被抽取组成杂合子的条件概率[17]。平均杂合度的大小可反映遗传结构变异程度的高低,杂合度越高说明该群体内遗传变异越大,遗传多样性也丰富;反之则群体内的遗传变异就小,遗传多样性匮乏。Jurg[18]将多态位点定义为遗传杂合度至少为0.10 的位点。耿岩等[19]对中国蒙系6个绵羊品种分析表明平均遗传杂合度在0.8208~0.9336之间。此次分析的10 个位点中,平均期望杂合度(Mean He)为0.6154,低于麦洼牦牛的平均期望杂合度(0.800)[20],牦牛种群遗传多样性较低,影响牦牛后期的发展和壮大,加大牦牛种群的保护力度是今后工作的一大重点。
3.2 遗传标记多态性分析
目前微量遗传样性的3个重要指标是平均多态
信息含量、平均杂合度和平均有效等位基因数,各参数在一定范围内数值越高,说明该群体的遗传样性也越高。杨章平等[21]利用7个微卫星标记揭示湖羊的平均PIC 为0.9024;柴文琼等[22]对甘肃地区的小尾寒羊群体进行分析,其PIC 为0.8293。李利等[23]利用5 个微卫星基因座对四川地方品种进行遗传多样性研究,结果表明所选微卫星在地方山羊群体中遗传多样性丰富。本研究中,10个基因座的平均PIC为0.685,ILSTS006和BM1824为中度多态基因座,其余均为高度多态基因座;平均光合度为0.6154,平均等位基因数为10.2,这10个微卫星位点均能反映本研究牦牛群体内丰富的遗传多态性,选育提高的潜力较大。

表2 牦牛10个STR 基因座基因频率分布Table 2 Allele frequency of 10STR Loci in yak
本试验进行Hardy-Weinberg平衡检验结果显示ILSTS006和BM1824处于不平衡状态,其原因可能是由于长期的人工繁殖过程中,人为选择作用所造成的基因型和基因频率的改变。

表3 10个位点的杂合度、多态信息含量和Hardy-Weinberg平衡Table 3 Heterozygosity,polymorphism information content and Hardy-Weinberg of 10STR Loci
3.3 银染法和DNA 自动测序仪检测结果的对比
在微卫星位点等位基因分型过程中利用银染法和DNA 自动测序仪进行数据采集。DNA自动测序仪比银染法得到的数据准确,客观,速度快,操作方便,但是比较贵。DNA 自动测序仪是用专用软件进行基因型的识别,加上专用的Mark使得收集数据准确。而银染需要人工进行染色处理,条带清晰度远没有DNA 测序仪所采集的图像清晰,同时在读取条带时受人为主观影响很大,同样的数据不同人处理会得到不同的结果。在同样的检测中DNA 测序仪进行分析所使用模板是银染法的三分之一,前期可以利用银染法进行多态性位点筛选,后期利用DNA 测序仪进行群体数据分析。
4 结论
本研究所分析的10个微卫星位点中,平均期望杂合度为0.6154,多态信息含量较高,显示出丰富的遗传多样性和较高的选择潜力。其中两个位点处于不平衡状态,可能的原因:母子关系记录不准确,父本局限在6头公牦牛,导致群体内公母牛比例较大而使整体的近交程度较高,造成了某些基因的频繁出现。目前微卫星遗传标记在牦牛上的研究较少,用于牦牛的检测位点较少,因此试验存在一定的误差,有待进一步改进。
[1]钟金城,陈志华,马志杰,等.牦牛分子育种的理论与实践[J].西南民族大学学报:自然科学版,2006,32(1):114-119.
[2]Gilmore S,Peakall R,Robertson J.Short tandem repeat(STR)DNA maekers are hypervariable and informative in Cannabis sative:implications for forensic investigations[J].Forensic Science International,2003,131(1):65-74.
[3]张云武,张亚平.微卫星及其应用[J].动物学研究,2001,2(4):315-320.
[4]陈振文,赵太云,孙贺娟,等.微卫星DNA 与生化标记分析对长爪沙鼠群体遗传分析的比较[J].中国实验动物学报,2006,14(4):276-279.
[5]闻旭阳,戴广海.Hmlh1基因启动子CpG 导甲基化,微卫星不稳定与胃癌[J].中国医药导报,2011,8(7):9-11.
[6]肖天放,柯柳玉,张 力,等.利用微卫星标记研究鸭品种的遗传多样性[J],应用生态学报,2009,20(1):190-196.
[7]张跟喜,丁馥香.利用微卫星标记分析边鸡遗传多样性及保种效果[J],农业生物技术学报,2012,18(5):944-950.
[8]张 扬,陈 阳,荣光辉,等.采用微卫星DNA 分析我国2个地方鹅种异地保种效果[J].中国畜牧杂志,2012,48(13):1-5.
[9]蔡 欣,泽让东科.麦洼牦牛群体4个新微卫星位点遗传多态性分析[J].西北农林科技大学学报:自然科学版,2014,42(5):30-31.
[10]Nguyen T,Genini S.Application of bovine microsatellite markers for Genetic diversity analysis of Swiss yak(Poephagus grunniens)[J].Anim Genet,2005,36(6):484-489.
[11]Qi X B,Han J L,Lkhagya B,et al.Genetic diversity and differentiation of Mongolian and Russian yak populationa[J].Anim Breed Genet,2005,122(2):117-126.
[12]廖信军,张贵香.中国五个地方牦牛品种遗传多样性的微卫星分析[J].生物多样性,2008,16(2):156-165.
[13]钟金城,陈志华,赵素君,等.牦牛生态类型的分类[J].生态学报,2006,26(7):9 601-9 605.
[14]Kalinowski S T,Marshall T C.Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment[J].Mol Ecol,2007,16(5):1 099-1 106.
[15]周智红,郭松长.藏羚羊5个微卫星标记的多态性分析[J].青海师范大学学报,2014,36(2):36-37.
[16]高建伟,贾文影等东北黑熊养殖群体6个微卫星基因座遗传多态性研究[J].畜牧与兽医,2014,46(3):71-72.
[17]Jurg O.Analysis of Human Genetic Linkage[M].Revised edition.Baltimore:Johns Hopkins University Press,2001:112-115.
[18]耿 岩,杨章平,常 洪,等.中国蒙系6个绵羊品种的遗传分化和基因流[J].扬州大学学报,2007,28(3):22-26.
[19]杨章平,常 洪,孙 伟,等.7个绵羊微卫星DNA 标记在绵(山)羊种群中的多态性检测[J].西北农林科技大学学报:自然科学版,2004,32(12):69-74.
[20]柴文琼,成述儒,靳建华,等.5个绵羊群体微卫星多态性分析[J].扬州大学学报,2012,33(1):39-42.
[21]李 利,刘建成,张国俊,等.用5个微卫星标记分析四川山羊品种的遗传关系[J].中国畜牧杂志,2009,45(3):8-11.
