淡水水生生态系统温室气体排放的主要途径及影响因素研究进展
2015-01-20杨平,仝川
杨 平, 仝 川
1 湿润亚热带生态-地理过程教育部重点实验室, 福州 350007 2 福建师范大学地理科学学院, 福州 350007 3 福建师范大学地理研究所, 福州 350007 4 福建师范大学亚热带湿地研究中心, 福州 350007
淡水水生生态系统温室气体排放的主要途径及影响因素研究进展
杨 平1,2, 仝 川1,3,4,*
1 湿润亚热带生态-地理过程教育部重点实验室, 福州 350007 2 福建师范大学地理科学学院, 福州 350007 3 福建师范大学地理研究所, 福州 350007 4 福建师范大学亚热带湿地研究中心, 福州 350007
淡水水生生态系统是全球陆域生态系统的重要组成部分,近年来,关于淡水水生生态系统温室气体排放的研究日益增多。基于国内外目前对湖泊、河流、水库及浅水池塘等淡水生态系统开展的最新研究成果,总结分析了淡水水生生态系统温室气体排放的3个主要途径及相应观测方法。气泡排放的观测方法有倒置漏斗法、开放式动态箱法和超声探测技术;植物传输的观测方法有密闭箱法和植株切割法;扩散途径的观测方法有静态浮箱法、模型估算法/梯度法、微气象学法、TDLAS吸收光谱法等。从物理因素、化学因素、生物因素、水动力因素和人类活动等角度,深入探讨了淡水水生生态系统温室气体排放通量的影响因素。最后根据当前研究中存在的不足,对今后的研究方向提出了建议,以期为我国进一步深入开展相关研究提供借鉴。
淡水水生生态系统; 温室气体; 排放途径; 观测方法; 影响因素
大气温室气体浓度增加及全球气候变暖引发的一系列生态与环境问题已引起人们对温室气体“源/汇”的广泛关注[1]。作为一个重要的生态系统类型,水生生态系统(aquatic ecosystems)排放到大气中的温室气体越来越引起人们的关注。水生生态系统作为温室气体重要的源和汇,对大气环境中温室气体浓度变化具有重要影响[2-4]。水生生态系统是地球表面各类水域生态系统的总称,按水体盐度高低可分为淡水生态系统和海洋生态系统,淡水又分为静水和流水生态系统。目前有关淡水生态系统温室气体排放的研究主要集中在静水生态系统,包括湖泊[5-7]、水库[8-10]和浅水池塘[11-12],而有关流水生态系统的研究鲜见报道[13-14]。由于水生生态系统温室气体的产生、转移及排放是一个复杂的、动态的生物地球化学过程,且这一过程受到诸多因素的影响,进而导致水生生态系统温室气体排放存在很大的不确定性。鉴于水生生态系统的重要性,近年来我国开展了一些相关的研究[15-18],但从广度和深度看,与国际上还有一定的差距。本文重点从陆域淡水水生生态系统(以下简称水生生态系统)温室气体(CO2、CH4和N2O)的排放途径、观测方法及影响因素等方面进行了归纳总结,以期为深入理解水生生态系统温室气体的产生与排放机制,为今后我国进一步开展水生生态系统温室气体产生、排放与调控等方面的研究提供思路与帮助。
1 温室气体的排放途径
水生生态系统温室气体主要通过沉积物-水-大气(或沉积物-植物、浮游植物-大气)体系向大气中排放(图1)[19]。河流、湖泊、池塘等水体环境产生的温室气体通过气泡排放、植物传输和扩散途径进入大气;对于水库而言,气体除通过上述3种途径进入大气外,还会通过水轮机、溢洪道和大坝下游等途径由水体逃逸至大气环境。

图1 水生生态系统温室气体的排放途径[19]Fig.1 Emission approaches of greenhouse gas derived from aquatic ecosystem[19]
1.1 气泡排放
气泡排放是无植物区域沉积物-水界面CH4等微溶气体迁移至水体表面进入大气的主要途径(图1)。该途径排放气体的强度在很大程度上受到不同气候带水体深度分压差的影响[20-21],但两者关系具有复杂性和不确定性。热带区域淡水水体(水库和湖泊)受水体静压力的作用,CH4通过气泡的排放量随深度增加而减少,且最大排放量往往发生在水深小于10 m的浅水域[21]。Rosa等[22]对巴西Samuel水库不同深度CH4排放通量的研究发现,水深为5 m或小于5 m处,CH4通过气泡的排放量为2.501—2.92 mg m-2h-1,而深度大于5 m处,排放量仅有0.08 mg m-2h-1。Keller等[23]对巴拿马Gatun湖泊,Galy-Lacaux等[24]和Rosa等[22]对亚马逊流域Tucuru水库研究也得出类似的结论。然而,Delsontro等[8]和Diem等[25]对北半球温带地区湖泊和水库,以及Bastien和Demarty[3]对南半球亚热带地区水库研究得出不同的结论。Delsontro等[8]和Diem[25]等研究发现,CH4通过气泡方式排放的最大通量源自于水下30 m以外的深水区域。Bastien和Demarty[3]对澳大利亚Koombooloomba水库的研究发现,CH4通过气泡方式排放的最大通量与水体深度及其分压差并不存在直接地关系。以上研究结论的不一致表明,不同深度分压差并不是造成气泡排放CH4强度呈现差异的唯一或主要因素,水生生态系统自身特征和其它外界因素也具有重要作用。
水体环境中气泡发生具有难以预测性、零星分布、持续时间极为短暂等特征,CH4气泡形成需具备3个条件:(1)沉积物具有较高的CH4产生速率;(2)水中气体的浓度较高;(3)水体压力超过沉积物溶液表面张力,其中,沉积物CH4产生速率对于水中气泡的形成及气泡数量的多寡起着十分重要的作用。理论上,当沉积物CH4产生速率较低时,一般不会或只产生极少量气泡;而一旦CH4产生速率超过某一临界值,沉积物液相中的CH4达到饱和,且来不及扩散的CH4气体分子便合并成CH4气体分子团形成富含CH4的气泡[8, 21]。水体中CH4气泡的形成、转移与释放过程是个十分复杂的动力学过程,除受水体环境不同深度分压差影响外,其他因素,如沉积物底质孔隙率、水体溶解氧、近表水层的温度效应、风速、碎浪作用、上升流变动、水流速度与方向、水生生物(如微生物、底栖动物)活动及大气边界层性质等亦显著干扰气泡释放过程,并且这些因素具有显著的时空变化[26-27]。目前国际上关于上述因素对CH4气泡排放的具体作用机制及其影响程度的研究已开展了相关工作,但国内并没有给予充分的关注。
相对于CH4气体,CO2因溶解度和液相阻力相对较大,气泡中含有的CO2量较少,所以气泡并不是水体中CO2进入大气环境的主要途径。已有研究显示,通过气泡排放途径进入大气环境中的CO2通量往往不到扩散通量的1%—2%[27-28]。Keller和Stallard[23]对巴拿马Gatun湖泊研究发现,98% CH4通过气泡排放途径进入大气,而CO2通过气泡排放的比例不到2%。Casper等[29]对英国一个低生产力湖泊(Priest Pot湖泊)CH4和CO2排放的研究也得出相似结论,发现CH4和CO2通过气泡途径排放的比例分别为96%和1%,并指出气泡排放存在显著的时空变化。Yang等[30]对三峡水库水-气界面CO2通量的研究结果显示,仅有5% 的CO2是通过气泡途径进入大气中。但Bastien和Demarty[3]对大洋洲Koombooloomba水库温室气体排放研究得出了不同的结论,发现Koombooloomba水库气泡途径排放的CO2通量高于CH4,与其它水库相比两种气体通过气泡途径的排放量相当,并维持在较低的水平(CH4:0.002 mg m-2h-1,CO2:0.003 mg m-2h-1)。以上不同的研究结果可能是由于研究区域地理位置、水体环境条件、观测方法(观测手段、观测时间及样品采集次数等)的不同而引起。
1.2 植物传输
水生植物为维持根的呼吸功能,往往形成输导组织向根部输送氧气,这些输导组织也成为植物传输水体温室气体(主要是CO2和CH4)的通道(图1)。在植物群落茂密的水域,植物传输是沉积物中CH4迁移到大气环境的重要途径,大约50%—90%的CH4可通过植物通气组织由沉积物直接输送到大气中[27],但不同植物群落之间的传输量存有较大差异。青藏高原花海不同植物群落CO2和CH4排放通量研究表明,与沉水植物(眼子菜Potamogeton)相比,挺水植物(杉叶藻Hippuris、藨草Scirpus、苔草Carex)具有较高的CO2和CH4排放通量,表现为眼子菜<藨草<苔草<杉叶藻[31-32]。段晓男等[33]对乌梁素海不同植物群落CH4排放通量的野外观测也得出类似结论,发现沉水植物(龙须眼子菜群落)CH4排放速率为3.44 mg m-2h-1,仅为挺水植物(芦苇(Phragmitesaustralis)群落)平均排放速率的21.94%。分析原因是不同植物群落对温室气体产生、氧化过程及其传输能力存在较大差异而引起。通常认为挺水植物可以将沉积物中产生的温室气体直接输送到大气中,而沉水植物只能将气体输送到水体中,再由水-气界面进入大气,该过程会增加CO2在植物体内被固定和CH4被氧化的可能[32-33]。此外,可能还会受植物传输温室气体机制的影响[12]。
植物传输温室气体的机制分为分子扩散和对流传输。分子扩散依赖于植物根与沉积物上部以及植物体内和大气之间的气体浓度差,扩散的能力主要受温度影响。该种机制较多地存在于沉水植物(如眼子菜)和一些小型挺水植物,如藨草(Scirpuslacustris)、苔草属植物(Carex)、贝母(Cladium)群落[32, 34],扩散规律可以用流体力学中Darcy扩散定律来描述[35]:
Q=KAJ
(1)
式中,Q为气体通量,K为扩散系数,A为通道面积,J为浓度梯度差。
对流传输是由气体的分压差驱动,气体随气流运动而输送,传输能力主要受到通过植物通气组织的湿度(湿度诱导)、热(热力学诱导)或者风速影响[36]。该种机制通常存在于芦苇、互花米草(Spartinaalterniflora)、香蒲(Typha)、王莲(Victoriaamazonica)等大型挺水植物或浮水植物中[37],传输规律可用气体运动理论描述:
Es=FA-1ΔP-1
(2)
(3)
ΔPt=Pwi-Pwa(湿度诱导)
(4)
式中,Es为气体对流效率(cm3min-1cm-2Pa-1),F为气流速率(cm3/min),A为叶片面积(cm2),ΔP为压强差,ΔPt为热力学压强差,ΔPw为湿度压强差,Pwi为叶片维管内湿度压强,Pwa为大气湿度压强,Pa为大气压强,Ti为叶片内温度,Ta为大气温度。一般来说,对流传输的输送效率高于分子扩散[37]。
1.3 扩散排放
扩散是指CO2、CH4或N2O等气体分子在浓度梯度下从高浓度向低浓度的一种任意运动,其通量F取决于气体溶解系数α、气体传质系数kg,T以及气体在水体与大气间的浓度差ΔP[4]:
F= αkg,TΔP
(5)
在进行水生生态系统温室气体通量扩散途径排放的研究中,气体传质系数的确定是核心,其传质系数受水-气界面紊流混合作用的控制[4],并且受到多种环境因素的影响。已有研究表明,风速或风场[38-39]、降雨[40]、热对流[41]、水流[42]、光照、水体酸碱度[26, 43]、有机质或悬浮物质[18]、水体氧化还原环境[19, 44]等均对气体传质系数产生影响。
1.4 其它途径
除上述3种途径外,水轮机、溢洪道和大坝下游也是水库温室气体排放的重要途径(图2)[4]。大部分水电大坝为了有足够的动力来运行水轮机,会将进出水口设计在距离水库表面数米低的地方,当富含气体的水通过水轮机时,水体静水压力发生急剧变化,大量气体将从水中逃逸出来。已有研究发现,水轮机和溢洪道是水库中温室气体逃逸至大气环境不可忽略的途径,但释放气体能力存在区域差异。Fearnside[45]对巴西Tucurui水库研究结果显示,水库CH4通过水轮机和溢洪道的排放量有时甚至会占到整个水库排放量的70%;而Diem[25]等对阿尔卑斯山区的Grimsel、Luzzone和Sihl水库研究发现,水库在泄水过程中因产生“水进复氧”现象,导致CH4被氧化,造成通过水轮机和溢洪道传输到大气中的CH4量减少,从而致使研究中的3个水库CH4通过水轮机的排放量仅占到水库总排放量的14%—44%。水库在泄水过程中通过水轮机和溢洪道作用除将部分温室气体排放到大气中外,水体中仍然含有部分的溶解有机碳、颗粒有机碳和溶解CO2、CH4排至水库下游,因此也有部分的CO2、CH4通过大坝下游途径释放到大气中。Guérin等[46]通过测量Petit Saut水库表层、下游河流0—40 m和40—80 m范围处的CO2和CH4发现,下游河流与水库深层次水体中的CO2和CH4浓度有着显著的相关性,多年跟踪研究结果显示,该水库约40% CO2和70% CH4的排放发生在大坝下游河流。Kemenes等[28, 47]对巴西Balbina水库CO2的研究也得出相似结果,但大坝下游CH4排放所占比例相对较低。
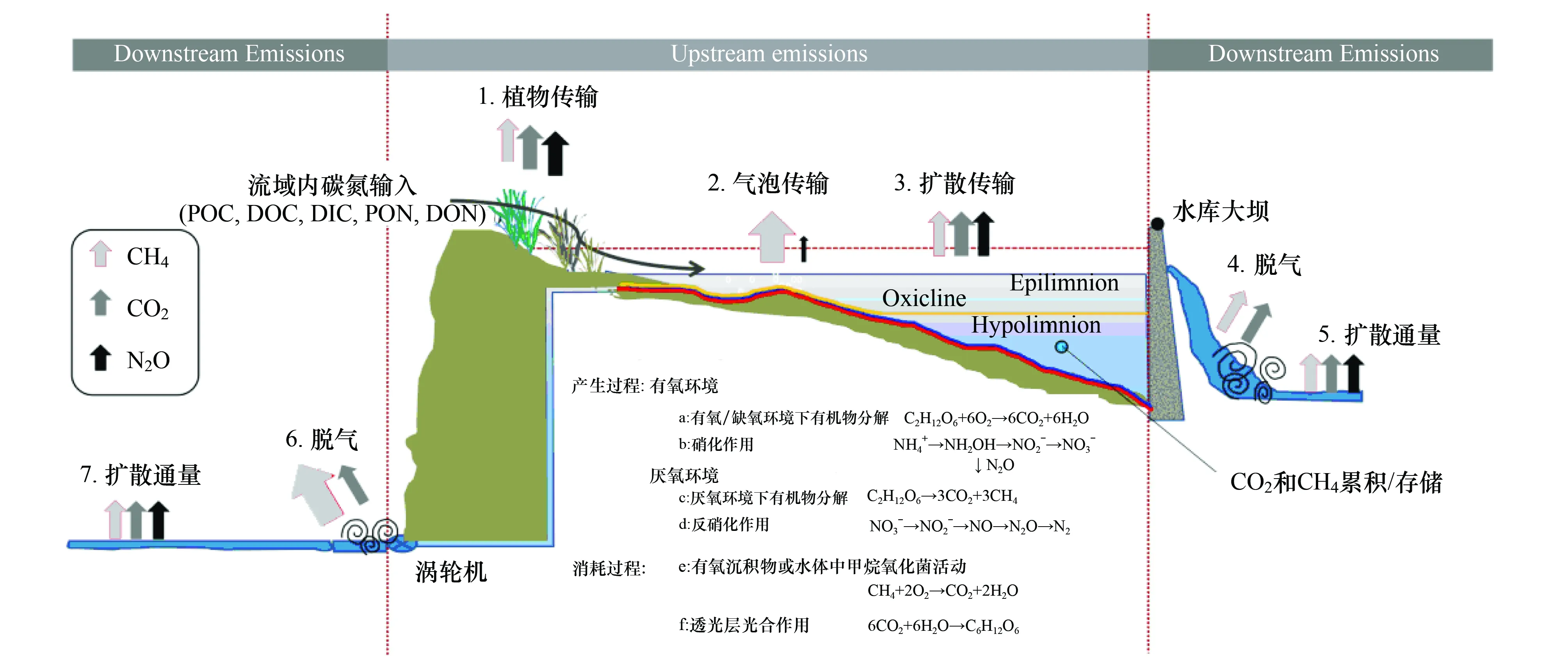
图2 水库生态系统温室气体产生与排放过程[4] Fig.2 Carbon dioxide, Methane and Nitrous oxide dynamics derived from hydroelectric reservoir ecosystem[4]
2 温室气体的观测方法
鉴于水生生态系统温室气体排放中各途径的重要性,目前国际上已有多种方法用于水生生态系统温室气体不同排放途径的研究(表1)。
(1)气泡排放和植物传输 倒置漏斗法是国际上用于气泡排放途径研究的常见方法,主要集中在热带、亚热带及一些浅水域温室气体通量的观测研究[3-4, 23, 48-49],无法实现区域连续监测和全面捕捉气泡排放信息是该种监测技术的不足。为弥补倒置漏斗法上述的缺陷,国际上已发展了开放式动态箱[50]和超声探测技术[51],优点是测量精确度高、连续性好,但技术要求较高。密闭箱法和植株切割法是用于观测水体中温室气体以植物通气组织方式排放的常见方法,但是两种方法亦仅局限于点的观测,无法进行大范围、全区域的连续监测;且样品监测过程中易受人为操作、天气及水面环境的影响,可能导致测定结果与实际情况存在误差。
(2)扩散排放途径 浮箱法[52-54]和模型估算法[9, 55-57]是目前用于水-气界面温室气体扩散通量最常用的两种野外现场监测方法。但在运用浮箱采集气样时,可能会因人为干扰和水面波动而改变箱内气体的浓度,从而导致测量的误差,并且该种方法也会受天气状况影响(通常在无雨期间操作)而无法实现全时段、大区域的连续观测。模型估算法中的气体交换系数k值的确定是决定该方法观测精度的关键,但k值时常受到表层水体紊流混合作用及环境因素影响而变的十分复杂,因此模型估算的结果可能存在较大的不确定性[58]。可调谐二极管激光吸收光谱(Tunable Diode Laser Absorption Spectroscopy, TDLAS)技术和微气象学观测方法中的涡度相关法是用于对水域温室气体扩散通量进行大面积连续测量最有效、精确度较高的两种方法,但其对环境条件和技术要求均较高,国际上目前已采用上述两种方法对面积较大的水库和湖泊进行观测研究[4, 39, 59],国内还比较鲜见。微气象学观测方法中的空气动力学法、能量平衡法和质量平衡法等也均可以用于现场测量水生生态系统温室气体扩散通量的计算[60]。
尽管已经有众多可行技术用于研究水生生态系统温室气体排放,但是,如何快速、有效而准确地评估大尺度范围温室气体排放通量时空变化的特征,对于当前科研人员来说仍是个重大挑战。

表1 水生生态系统温室气体通量的主要研究方法
3 温室气体排放通量的主要影响因素
水生生态系统温室气体排放通量大小由产生和传输过程共同决定,对于CH4气体来说,氧化也是一个重要环节。以上过程受到诸多环境因素的影响,包括气象因素、水体理化性质、生物作用、水动力条件等。
3.1 物理因素
3.1.1 温度
水温是水体重要物理参数,对水体CO2通量影响因生境不同而呈现差异。在相对封闭或静止的水体中(如天然湖泊和浅水池塘),较高水温促使有机质分解,引起水体二氧化碳分压(pCO2)上升,增加水体CO2向大气释放[11, 61]。对河流和水库而言,纵向输移和垂向混合的生境特征迫使该种水生生态系统具有一定开放性水温特点,致使水体生物地球化学过程同湖泊有较大差异。在上述生境下,较高水温会增强水生生物光合作用对水体中溶解性CO2吸收,提高水体pH,降低pCO2,进而增加水体对大气CO2吸收[18, 62]。
水温通过影响微生物活性、氧化还原环境及气体在水中溶解度间接的影响到CH4和N2O排放。水温升高会激发微生物活性,降低气体在水中的溶解,促进水体中CH4和N2O向大气释放[62-63]。Avery等[64]采用14C标记对美国北卡罗莱纳州White Oak河流沉积物的研究发现,沉积物中乙酸发酵途径产生的CH4约占CH4产生总量的69%,CH4产生速率随温度升高呈指数增长,沉积物中CH4产生的微生物活性主要受控于温度。Silvennoinen等[65]对水体N2O排放速率研究发现,随着水温增加,水体沉积物的硝化和反硝化速度因微生物活性增强而促进了水-气界面N2O的排放速率。也有研究表明,较高水温易导致水体缺氧而抑制硝化作用,进而会抑制N2O的产生与释放[11]。
气温通过对水温和沉积物底质温度的调节作用引起水体温室气体排放通量呈现显著地时间变化特征[5, 10, 12]。Garcia-Ruiz等[13]对英国Swale-Ouse河5个不同地段的研究发现,各河段水体最大N2O排放量均出现于气温较高的5—10月,而其它时间明显降低。Xing等[15]对亚热浅水湖泊CH4排放的研究也发现,湖泊向大气排放CH4的峰值出现在气温较高的夏季,其它季节CH4排放量均维持在较低水平。但不同水体环境温室气体排放通量日变化与气温的关系具有一定的复杂性,主要表现为:无相关性[63],正相关[12, 20, 66]或负相关[67]。
3.1.2 风速
风速通过风应力致使水体表面破碎,增加水体与空气接触面积,改变表层水与大气间的CO2浓度梯度来影响气体扩散的强度[38, 48]。在浅水环境中,风力扰动作用还会促使沉积物中碳酸盐溶解于水体,引起水体碱度上升,增加水体CO2向大气中释放[12, 63]。
风速对水-气界面CH4排放的影响具有双重性:一是风力对水体的扰动将引起沉积物层产生大量气泡[22, 68],并且通过剪切水面波浪降低水-气界面的气体扩散阻力来增加水体CH4的排放;二是风力扰动会增加水中溶解氧(DO)含量和增强深水层中的氧化作用,产甲烷菌活动因而受抑制,从而减少了CH4的产生与排放[43]。
风速可通过影响N2O在水-气界面的分压平衡来影响水体N2O释放,主要表现为:一方面,风的切应力导致水体表面破碎,增加水-气交换面积,促进水体N2O释放;另一方面,风速变化通过影响N2O在水体中的饱和度引起水体中N2O源/汇角色转换。在高风速条件下,水体N2O极有可能出现欠饱和,在该种情形下水体会形成向下的泵吸作用[69],促使水体对大气中N2O的吸收[70]。
3.1.3 光合有效辐射
光合有效辐射(PAR)通过影响温度、植物及微生物活性间接影响水体温室气体的地球化学过程,并且这种影响因水体植被覆盖状况而呈现出差异性。在水生植物覆盖率较高或浮游植物较丰富的水环境中,水体与大气间CO2通量的昼夜变化受太阳辐射影响[12, 71]。日间太阳辐射较强,植物代谢过程旺盛,强烈的光合作用大量吸收水中溶解性CO2,引起水体pH上升,致使水体pCO2低于大气水平,从而削减水体系统总呼吸的CO2量[18, 71];而夜间的情况正好与之相反。
PAR可通过影响植物光合作用强度以及微生物活性间接的影响到CH4产生与排放。昼间强烈的PAR促使植物光合作用释放O2,激发CH4氧化菌活性,致使沉积物产生的CH4被较快氧化[11-12]。此外,昼间PAR可提高水生植物体内的水汽蒸发和温度,加大植物体内湿度诱导的压强梯度和热力学诱导渗透,进而加强气体对流;而一旦光照条件消失,气流则停止[35];因此,PAR也会影响植物传输CH4机制的昼夜转换。很多水生植物,如芦苇、宽叶香蒲(Typhalatifolia)、长苞香蒲(Typhadomingensis)等在低光照或夜间的时候,采用分子扩散机制来排放CH4;而在日间光照充足的条件下,以对流传输为主[33-34]。
PAR可通过引起水体溶解氧变化来对水体中N2O的产生与排放过程产生影响。强光照条件下,沉积物反硝化速率因水体氧气渗透能力增强而降低[72],从而造成通过反硝化途径产生的N2O减少[70]。此外,该过程中的硝化速率也随水体氧的渗透能力增强而提高,但在水体环境下N2O主要来源于沉积物反硝化过程,硝化作用增加的N2O量不足以抵偿反硝化过程削弱而减少的量。因此,强光照条件下水体向大气中释放的N2O可能会减少[66, 73]。
3.2 化学因素
3.2.1 pH值和氧化还原电位
氧化还原电位对水体温室气体排放通量影响具有不可忽视的作用。但目前关于水生生态系统CO2和CH4排放通量对氧化还原电位的响应存有争议。一方面认为,氧化还原电位是反映沉积物或水体环境氧化还原能力强弱的一个重要指标,是有机质分解的制约因素,直接关系到温室气体的产生及在迁移过程中的转化,对水体CH4和CO2排放有着显著影响[76]。另一方面认为,野外风速和风向对水体扰动存在随机性,进而导致水体氧含量不断发生变化,从而使得氧化还原电位对水体CH4和CO2排放通量的影响具有不确定性[67]。
3.2.2 溶解氧和溶解性有机碳
水体溶解氧(DO)含量及分布情况决定着水体中有机物降解途径及其产物,对水体生源要素循环有重要影响[68]。对于碳循环而言,有机物在有氧情况下产生CO2和CH4,而在缺氧状态下主要产生CH4;对于氮循环而言,在有氧环境中主要发生硝化反应,厌氧环境中主要发生反硝化反应。Rosa等[22]对热带地区水库研究发现,沉积物中产生的CH4,一部分通过扩散方式上升到水面,在上升过程中,由于水中氧含量的逐渐上升,产生的大部分CH4被有氧-缺氧临界面的甲烷氧化菌消耗掉,从而使得水体扩散到大气的CH4量明显减少。Xing等[15]对亚热带浅水湖泊研究发现,湖泊水体夏季向大气排放的CH4大于春季、秋季及冬季,除受湖泊初级生产力影响外,夏季水体氧含量较低、厌氧程度高也是一个重要影响因素。Schrier-Uijl等[63]研究也发现,不同水体环境中CH4排放量与水体DO变化有着显著的负相关关系,通常情况下DO低的水体排放的CH4要大于DO高的水体。韩洋等[70]研究发现,南京典型水体春季N2O排放量昼间低于夜间,与昼间水体DO高致使硝化作用增强有关。
溶解性有机碳(DOC)丰富程度对水体中CO2和CH4的产生与排放具有重要影响[9, 77]。通常情况下,单位面积水体的CO2和CH4通量与流域或水体中DOC浓度呈正相关关系[20, 78]。对水库的研究表明[18, 21, 23],在水库运行初期因水库淹没大量植被和土壤,水体中易分解的DOC浓度高,CO2和CH4排放量亦高;随着水库年龄增加,DOC浓度降低,CO2和CH4排放通量也随之减少,最终趋于天然河道水平。DOC影响水体CO2和CH4排放通量的主要作用机制是DOC为水体中CO2和CH4产生的直接碳源,其含量高低直接关系到相关微生物活性。此外,DOC对水生生态系统p(CO2)也具有一定影响,并且这种影响因湖泊或水库所处流域中DOC丰富程度、来源途径、水体规模及地理位置不同而呈现出差异[43]。
3.2.3 营养盐
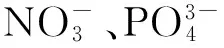

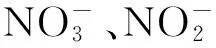
3.3 生物因素
水生生物对水-气界面CO2交换通量的影响主要体现在2个方面,一方面在自养状态下,水生浮游植物初级生产力大于水生生物呼吸作用,碳被固定,引起水体p(CO2)下降,促使水体吸收大气中的CO2;另一方面,在异养状态下,水生生物呼吸作用强于光合作用,水体p(CO2)增加,促使水体向大气中释放CO2[18, 21, 67]。在对CH4的影响方面,水生生物活动可通过影响水中DO含量而间接影响水-气界面CH4通量。一方面,水生浮游植物的光合作用可增加水中DO含量,造成水体中CH4氧化量增加;另一方面,水生生物通过呼吸作用、有机物的分解等途径消耗水中DO含量,CH4产生与排放量因厌氧环境增强而增大[68]。
植被对水生生态系统温室气体排放的影响不可忽视,其主要通过以下机制或途径产生作用:(1)植物通过光合作用固定碳或降低水流流速富集大量的有机物[16, 86],为相应气体产生提供充足的碳氮源;(2)植物通过根系周转及其对碳物质的分泌[87],为微生物活动提供原料;(3)为沉积物厌氧区生成的CH4、N2O等气体直接进入大气提供通道[86, 88-89];(4)植物自身产生或吸收CH4气体[90]。
3.4 水动力因素
温室气体从水体释放到大气的过程是一个动力学过程,与水流动力及水-气界面动力息息相关[82]。水流速度通过影响藻类生长环境及水体富营养化来影响光在水体中的通透性[68],进而影响到水生生物在水体碳氮循环过程中的活性。Silvennoinen等[65]波罗的海Temmesjoki河的研究发现,较快的水流速度及较大的风速有利于水-气界面温室气体交换。此外,水位季节性波动影响到深层水体温度的变化,进而影响到有机物分解速率及气泡的比率[72],从而导致温室气体排放量也会发生季节性变化[3, 68, 91]。
3.5 人类活动的影响
土地利用方式变化、大坝与水库建设改变了水生生态系统原有的自然演进和变化过程,直接影响到C、N等生源要素在水体中的生物地球化学行为。陆域湖泊多属于浅水富营养性湖泊,浮游植物生产力高,水生高等植物丰富,绝大多数情况下是吸收大气中的CO2,可能是未被发现的重要碳汇[92]。已有研究初步估计,湖泊对大气CO2的总汇(以C计算)可达5.32×107t/a[92]。但长期盲目的土地利用方式变化,如天然湖泊围垦转变为农田,导致湖泊生态系统碳的大量流失,湖泊在水生生态系统中的碳汇能力正在逐步地被弱化。大坝与水库建设是当前人类活动对河流碳氮循环过程影响最为主要的干扰方式。河流上大坝和水库的广泛存在,一方面加大了来自河流上游输移的陆源侵蚀物质的沉积量[93],加之大坝的拦截使水体停留时间延长,有利于自源含碳物质的生产[94]和有机物的降解,进而会改变水体的碳汇/源功能,其中寒带及温带地区水体主要表现为CO2的“源”,热带地区主要表现为CH4的释放“源”[95]。另一方面大坝下游水流速度趋缓,加上沿途大量工农业废水及居民生活污水的排入,水体营养盐浓度升高,易导致水体富营养化[95],直接引起水体理化状况的变化[96],势必也会影响到大坝下游河流水体温室气体的产生与排放过程[91]。但目前关于建坝前后的大坝下游河流水体温室气体通量差异性对比的研究鲜见报道。
内陆水域水产养殖是影响人工池塘和天然湖泊温室气体通量变化较为常见的一种人为干扰方式。据Boyd等人的研究估算[97],世界范围内1.11×105km2的水产养殖塘的沉积物每年可封存1.66×107t的CO2(以C计算),占全球每年碳排放总量的0.21%,水产养殖业在固定(形成碳汇)和移除(碳提取)水生生态系统碳方面发挥着重要作用。但是目前国际上在该方面的研究仅仅是从渔业统计数据角度初步分析水产养殖塘的能耗及构成,据此来估算水产养殖活动中CO2的排放强度[92],对于直接开展水产养殖活动对水体碳氮循环过程及其温室气体排放影响的研究鲜见报道[80]。
4 问题与展望
作为全球生态系统的重要组成部分,淡水生态系统温室气体的产生和排放过程具有自身的特点,近年来引起较多的关注,并取得一定的研究成果,但仍存在着局限性。鉴于当前研究中存在的问题,建议今后的研究方向可侧重于以下几个方面:
(1)加强对水生生态系统温室气体通量测定方法的标准化研究。尽管已有许多方法用于淡水生态系统温室气体通量的监测,但目前各种方法的测定结果之间的可比性较差。因此,探索有效而准确的测定方法,并统一标准对今后开展水生生态系统温室气体通量的连续观测具有重要意义。
(2)加强对水生生态系统温室气体排放通量的大范围长期连续观测研究。当前对水生生态系统温室气体排放研究主要聚集在定点观测上,并据此来表征整个水域温室气体排放通量的动态特征及其总排放量。水生生态系统自身特性及其环境因素的时空异质性势必会对整个观测结果产生影响,导致研究结果存在不确定性和较大误差,有必要采用有效监测技术进行大范围长期连续观测研究。
(3)继续深化水生生态系统温室气体各种排放途径的研究。综合考虑多种因素(包括水生生态系统自身特性、气象因素、水体理化性质、生物因素和水动力因素)、观测时间(昼夜变化和季节变化观测)、加大野外采样频率等方面对水生生态系统温室气体不同排放途径进行观测研究,以期明晰不同排放途径的具体作用机制及其相应时空变化特征。
(4)加强水生生态系统不同界面的温室气体排放研究。水生生态系统温室气体的排放,不仅涉及到水体内部变化,而且与沉积物、陆源输入等都有关联。目前主要聚集在水-气界面温室气体排放的研究,忽视了对水-沉积物界面、水-陆界面温室气体排放通量的研究。
(5)积极开展人类活动影响下水生生态系统温室气体排放特征的研究。土地利用方式变化、大坝与水库建设等人类活动改变了水生生态系统原有的自然演进过程,工农业废水和居民生活污水大量排放引起的水体富营养化,均直接影响到C、N等生源要素在水体中的生物地球化学行为,有必要深入探讨人类活动对水生生态系统温室气体排放的影响。
[1] Intergovernmental Panel on Climate Change. Climate Change 2007:The Physical Science Basis. New York:Cambridge University Press, 2007.
[2] Duc N T, Silverstein S, Lundmark L, Reyier H, Crill P, Bastviken D. Automated flux chamber for investigating gas flux at water-air interfaces. Environmental Science & Technology, 2012, 47(2):968-975.
[3] Bastien J, Demarty M. Spatio-temporal variation of gross CO2and CH4diffusive emissions from Australian reservoirs and natural aquatic ecosystems, and estimation of net reservoir emissions. Lakes & Reservoirs:Research & Management, 2013, 18(2):115-127.
[4] Deshmukh C. Greenhouse gas emissions (CH4, CO2and N2O) from a newly flooded hydroelectric reservoir in subtropical South Asia:The case of Nam Theun 2 Reservoir, Lao PDR. New Delhi, India:Université Paul Sabatier -Toulouse III; Teri University, 2013.
[5] Repo M E, Huttunen J T, Naumov A V, Chichulin A V, Lapshina E D, Bleuten W, Martikainen P J. Release of CO2and CH4from small wetland lakes in western Siberia. Tellus, 2007, 59(5):788-796.
[6] Juutinen S, Rantakari M, Kortelainen P, Huttunen J T, Larmola T, Alm J, Silvola J, Martikainen P J. Methane dynamics in different boreal lake types. Biogeosciences, 2009, 6:209-223.
[7] Palma-Silva C, Marinho C C, Albertoni E F, Giacomini I B, Barros M P F, Furlanetto L M, Trindade C R T, Esteves F D A. Methane emissions in two small shallow neotropical lakes:The role of temperature and trophic level. Atmospheric Environment, 2013, 81:373-379.
[8] DelSontro T, McGinnis D F, Sobek S, Ostrovsky I, Wehrli B. Extreme methane emissions from a Swiss hydropower reservoir:Contribution from bubbling sediments. Environmental Science & Technology, 2010, 44(7):2419-2425.
[9] Jacinthe P A, Filippelli G M, Tedesco L P, Raftis R. Carbon storage and greenhouse gases emission from a fluvial reservoir in an agricultural landscape. CATENA, 2012, 94:53-63.
[10] Yang L, Lu F, Wang X K, Duan X N, Song W Z, Sun B F, Zhang Q Q, Zhou Y J. Spatial and seasonal variability of diffusive methane emissions from the Three Gorges Reservoir. Journal of Geophysical Research:Biogeosciences, 2013, 118(2):471-481.
[11] 林海, 周刚, 李旭光, 周军, 张彤晴, 王桂民. 夏季池塘养殖中华绒螯蟹生态系统温室气体排放及综合增温潜势. 水产学报, 2013, 37(3):417-424.
[12] Natchimuthu S, Selvam B P, Bastviken D. Influence of weather variables on methane and carbon dioxide flux from a shallow pond. Biogeochemistry, 2014, 119(1/3):403-413.
[13] García-Ruiz R, Pattinson S N, Whitton B A. Nitrous oxide production in the river Swale-Ouse, North-East England. Water Research, 1999, 33(5):1231-1237.
[14] Koné Y J M, Abril G, Delille B, Borges A V. Seasonal variability of methane in the rivers and lagoons of Ivory Coast (West Africa). Biogeochemistry, 2010, 100(1/3):21-37.
[15] Xing Y P, Xie P, Yang H, Ni L Y, Wang Y S, Rong K W. Methane and carbon dioxide fluxes from a shallow hypereutrophic subtropical lake in China. Atmospheric Environment, 2005, 39(30):5532-5540.
[16] Duan X N, Wang X K, Mu Y J, Ouyang Z Y. Seasonal and diurnal variations in methane emissions from Wuliangsu Lake in arid regions of China. Atmospheric Environment, 2005, 39(25):4479-4487.
[17] Liu X L, Liu C Q, Li S L, Wang F S, Wang B L, Wang Z L. Spatiotemporal variations of nitrous oxide (N2O) emissions from two reservoirs in SW China. Atmospheric Environment, 2011, 45(31):5458-5468.
[18] 蒋滔, 郭劲松, 李哲, 方芳, 白镭, 刘静. 三峡水库不同运行状态下支流澎溪河水-气界面温室气体通量特征初探. 环境科学, 2012, 33(5):1463-1470.
[19] Diem T. Methane dynamics in oxic and anoxic aquatic systems. Discussions, Eidgenössische Technische Hochschule ETH Zürich, Nr. 18113, 2008.
[20] Huttunen J T, Alm J, Liikanen A, Juutinen S, Larmola T, Hammar T, Silvola J, Martikainen P J. Fluxes of methane, carbon dioxide and nitrous oxide in boreal lakes and potential anthropogenic effects on the aquatic greenhouse gas emissions. Chemosphere, 2003, 52(3):609-621.
[21] Abril G, Guérin F, Richard S, Delmas R, Galy-Lacaux C, Gosse P, Tremblay A, Varfalvy L, Dos Santos M A, Matvienko B. Carbon dioxide and methane emissions and the carbon budget of a 10-year old tropical reservoir (Petit Saut, French Guiana). Global Biogeochemical Cycles, 2005, 19(4), doi:10.1029/2005GB002457.
[22] Rosa L P, Santos M A D, Matvienko B, Sikar E, Lourenço R S M, Menezes C F. Biogenic gas production from major Amazon reservoirs, Brazil. Hydrological Processes, 2003, 17(7):1443-1450.
[23] Keller M, Stallard R F. Methane emission by bubbling from Gatun Lake, Panama. Journal of Geophysical Research, 1994, 99(D4):8307-8319.
[24] Galy-Lacaux C, Delmas R, Kouadio G, Richard S, Gosse P. Long-term greenhouse gas emissions from hydroelectric reservoirs in tropical forest regions. Global Biogeochemical Cycles, 1999, 13(2):503-517.
[25] Diem T, Koch S, Schwarzenbach S, Wehrli B, Schubert C J. Greenhouse gas emissions (CO2, CH4and N2O) from several perialpine and alpine hydropower reservoirs by diffusion and loss in turbines. Aquatic Sciences, 2012, 74(3):619-635.
[26] 鲁中明, 戴民汉. 海气CO2通量与涡动相关法应用研究进展. 地球科学进展, 2006, 21(10):1046-1057.
[27] Kelly C A, Rudd J W M, Bodaly R A, Roulet N P, Louis V L St, Heyes A, Moore T R, Schiff S, Aravena R, Scott K J, Dyck B, Harris R, Warner B, Edwards G. Increases in fluxes of greenhouse gases and methyl mercury following flooding of an experimental reservoir. Environmental Science & Technology, 1997, 31(5):1334-1344.
[28] Kemenes A, Forsberg B R, Melack J M. CO2emissions from a tropical hydroelectric reservoir (Balbina, Brazil). Journal of Geophysical Research, 2011, 116(G3), doi:10.1029/2010JG001465.
[29] Casper P, Maberly S C, Hall G H, Hall G H, Finlay B J. Fluxes of methane and carbon dioxide from a small productive lake to the atmosphere. Biogeochemistry, 2000, 49(1):1-19.
[30] Yang L, Lu F, Wang X K, Duan X N, Tong L, Ouyang Z Y, Li H P. Spatial and seasonal variability of CO2flux at the air-water interface of the Three Gorges Reservoir. Journal of Environmental Sciences, 2013, 25(11):2229-2238.
[31] Hirota M, Tang Y, Hu Q W, Hirata S, Kato T, Mo W H, Cao G M, Mariko S. Methane emissions from different vegetation zones in a Qinghai-Tibetan Plateau wetland. Soil Biology & Biochemistry, 2004, 36(5):737-748.
[32] Hirota M, Tang Y, Hu Q W, Hirata S, Kato T, Mo W H, Cao G M, Mariko S. Carbon dioxide dynamics and controls in a deep-water wetland on the Qinghai-Tibetan Plateau. Ecosystems, 2006, 9(4):673-688.
[33] 段晓男, 王效科, 陈琳, 牟玉静, 欧阳志云. 乌梁素海湖泊湿地植物区甲烷排放规律. 环境科学, 2007, 28(3):455-459.
[34] Van Der Nat F J W A, Middelburg J J, Van Meteren D, Wielemakers A. Diel methane emission patterns fromScirpuslacustrisandPhragmitesaustralis. Biogeochemistry, 1998, 41(1):1-22.
[35] 成水平, 吴振斌, 夏宜琤. 水生植物的气体交换与输导代谢. 水生生物学报, 2003, 27(4):413-417.
[36] Arkebauer T J, Chanton J P, Verma S B, Kim J. Field measurements of internal pressurization inPhragmitesaustralis(Poaceae) and implications for regulation of methane emissions in a midlatitude prairie wetland. American Journal of Botany, 2001, 88(4):653-658.
[37] 段晓男, 王效科, 欧阳志云. 维管植物对自然湿地甲烷排放的影响. 生态学报, 2005, 25(12):3375-3382.
[38] Wanninkhof R. Relationship between wind speed and gas exchange over the ocean. Journal of Geophysical Research, 1992, 97(C5):7373-7382.
[39] Guérin F, Abril G, Serça D, Delon C, Richard S, Delmas R, Tremblay A, Varfalvy L. Gas transfer velocities of CO2and CH4in a tropical reservoir and its river downstream. Journal of Marine Systems, 2007, 66(1/4):161-172.
[40] Ho D T, Veron F, Harrison E, Bliven L F, Scott N, Mcgillis W R. The combined effect of rain and wind on air-water gas exchange:A feasibility study. Journal of Marine Systems, 2007, 66(1/4):150-160.
[41] Eugster W, Kling G T, Jonas J P, McFadden A, MacIntyre S, Chapin F S III. CO2exchange between air and water in an arctic Alaskan and midlatitude Swiss lake:importance of convective mixing. Journal of Geophysical Research, 2003, 108(D12), doi:10.1029/2002JD002653.
[42] Zappa C J, McGillis W R, Raymond P A, Edson J B, Hintsa E J, Zemmelink H J, Dacey J W H, Ho T. Environmental turbulent mixing controls on air-water gas exchange in marine and aquatic systems. Geophysical Research Letters, 2007, 34(10):L10601, doi:10.1029/2006GL028790.
[43] 赵小杰, 赵同谦, 郑华, 段晓男, 陈法霖, 欧阳志云, 王效科. 水库温室气体排放及其影响因素. 环境科学, 2008, 29(8):2377-2384.
[44] 张晓艳, 马静, 李小平, 徐华, 蔡祖聪. 稻田甲烷传输的研究进展. 土壤, 2012, 44(2):181-187.
[45] Fearnside P M. Greenhouse gas emissions from hydroelectric dams:Controversies provide a springboard for rethinking a supposedly “clean” energy source.AnEditorialComment. Climatic Change, 2004, 66(1/2):1-8.
[46] Guérin F, Abri G, Richard S, Burban B, Reynouard C, Seyler P, Delmas R. Methane and carbon dioxide emissions from tropical reservoirs:Significance of downstream rivers. Geophysical Research Letters, 2006, 33(21):L21407. doi:10.1029/2006GL027929.
[47] Kemenes A, Forsberg B R, Melack J M. Methane release below a tropical hydroelectric dam. Geophysical Research Letters, 2007, 34(12):L12809. doi:10.1029/2007GL029479.
[48] Tremblay A, Varfalvy L, Roehm C, Garneau M. Greenhouse Gas Emissions-Fluxes and Processes:Hydroelectric reservoirs and natural environments. New York:Springer, 2005.
[49] Flury S, McGinnis D F, Gessner M O. Methane emissions from a freshwater marsh in response to experimentally simulated global warming and nitrogen enrichment. Journal of Geophysical Research, 2010, 115(G1):G01007, doi:10.1029/2009JG001079.
[50] Ramos F M, Lima I B T, Rosa R R, Mazzi E A, Carvalho J C, Rasera M F F L, Ometto J P H B, Assireu A T, Stech J L. Extreme event dynamics in methane ebullition fluxes from tropical reservoirs. Geophysical Research Letters, 2006, 33(21):L21404, doi:10.1029/2006GL027943.
[51] Ostrovsky I, McGinnis D F, Lapidus L, Eckert W. Quantifying gas ebullition with echosounder:the role of methane transport by bubbles in a medium-sized lake. Limnology and Oceanography:Methods, 2008, 6(2):105-118.
[52] Zhu R B, Liu Y S, Xu H, Huang T, Sun J J, Ma E D, Sun L G. Carbon dioxide and methane fluxes in the littoral zones of two lakes, east Antarctica. Atmospheric Environment, 2010, 44(3):304-311.
[53] Bastien J, Demarty M, Tremblay A. CO2and CH4diffusive and degassing fluxes from 2003 to 2009 at Eastmain 1 hydroelectric reservoir, Québec, Canada. Inland Waters, 2011, 1(2):113-123.
[54] Ding W, Zhu R B, Ma D W, Xu H. Summertime fluxes of N2O, CH4and CO2from the littoral zone of Lake Daming, East Antarctica:effects of environmental conditions. Antarctic Science, 2013, 25(6):752-762.
[55] Demarty M, Bastien J, Tremblay A. Annual follow-up of gross diffusive carbon dioxide and methane emissions from a boreal reservoir and two nearby lakes in Québec, Canada. Biogeosciences, 2011, 8:41-53.
[56] Schilder J, Bastviken D, van Hardenbroek M, Kankaala P, Rinta P, Stöetter T, Heiri O. Spatial heterogeneity and lake morphology affect diffusive greenhouse gas emission estimates of lakes. Geophysical Research Letters, 2013, 40(21):5752-5756.
[57] 高洁, 郑循华, 王睿, 廖婷婷, 邹建文. 漂浮通量箱法和扩散模型法测定内陆水体CH4和N2O排放通量的初步比较研究. 气候与环境研究, 2014, 19(3):290-302.
[58] 赵炎, 曾源, 吴炳方, 陈永柏, 王强, 袁超. 水库水气界面温室气体通量监测方法综述. 水科学进展, 2011, 22(1):135-146.
[59] Huotari J, Ojala A, Peltomaa E, Nordbo A, Launiainen S, Pumpanen J, Rasilo T, Hari P, Vesala T. Long-term direct CO2flux measurements over a boreal lake:Five years of eddy covariance data. Geophysical Research Letters, 2011, 38(18):L18401, doi:10.1029/2011GL048753.
[60] 王庚辰. 陆地生态系统温室气体排放(吸收)测量方法简评. 气候与环境研究, 1997, 2(3):251-263.
[61] Rantakari M, Kortelainen P. Interannual variation and climatic regulation of the CO2emission from large boreal lakes. Global Change Biology, 2005, 11(8):1368-1380.
[62] Soumis N, Duchemin É, Canuel R, Lucotte M. Greenhouse gas emissions from reservoirs of the western United States. Global Biogeochemical Cycles, 2004, 18(3):GB3022, doi:10.1029/2003GB002917.
[63] Schrier-Uijl A P, Veraart A J, Leffeaar P A, Berendse F, Veenendaal E M. Release of CO2and CH4from lakes and drainage ditches in temperate wetlands. Biogeochemistry, 2011, 102(1/3):265-279.
[64] Avery G B, Shannon R D, White J R, Martens C S, Alperin M J. Controls on methane production in a tidal freshwater estuary and a peatland:methane production via acetate fermentation and CO2reduction. Biogeochemistry, 2003, 62(1):19-37.
[65] Silvennoinen H, Liikanen A, Torssonen J, Stange C F, Martikainen P J. Denitrification and N2O effluxes in the Bothnian Bay(northern Baltic Sea)river sediments as affected by temperature under different oxygen concentrations. Biogeochemistry, 2008, 88(1):63-72.
[66] Clough T J, Buckthought L, Kelliher F, Sherlock R. Diurnal fluctuations of dissolved nitrous oxide (N2O) concentrations and estimates of N2O emissions from a spring-fed river:implications for IPCC methodology. Global Change Biology, 2007, 13(15):1016-1027.
[67] 王亮, 肖尚斌, 刘德富, 陈文重, 王雨春, 陈小燕, 段玉杰. 香溪河库湾夏季温室气体通量及影响因素分析. 环境科学, 2012, 33(5):1471-1475.
[68] 程炳红, 郝庆菊, 江长胜. 水库温室气体排放及其影响因素研究进展. 湿地科学, 2012, 10(1):121-128.
[69] Crosswell J R, Wetz M S, Hales B, Paerl H W. Air-water CO2fluxes in the microtidal Neuse River Estuary, North Carolina. Journal of Geophysical Research:Oceans, 2012, 117(C8), doi:10.1029/2012JC007925.
[70] 韩洋, 郑有飞, 吴荣军, 尹继福, 孙霞. 南京河流夏季水-气界面N2O排放通量. 环境科学, 2014, 35(1):348-355.
[71] Davies J M, Hesslein R H, Kelly C A, Carol A, Hecky R E.PCO2method for measuring photosynthesis and respiration in freshwater lakes. Journal of Plankton Research, 2003, 25(4):385-395.
[72] Laursen A E, Carlton R G. Responses to atrazine of respiration, nitrification, and denitrification in stream sediments measured with oxygen and nitrate microelectrodes. FEMS Microbiology Ecology, 1999, 29(3):229-240.
[73] 韩洋, 郑有飞, 吴荣军, 尹继福, 徐静馨, 徐盼. 南京典型水体春季温室气体排放特征研究. 中国环境科学, 2013, 33(8):1360-1371.
[74] Dumestre J F, Vaquer A, Gosse P, Richard S, Labroue L. Bacterial ecology of a young equatorial hydroelectric reservoir (Petit Saut, French Guiana). Hydrobiologia, 1999, 400:75-83.
[75] Garcia J L, Patel B K, Ollivier B. Taxonomic, phylogenetic, and ecological diversity of methanogenicArchaea. Anaerobe, 2000, 6(4):205-226.
[76] Singh S N, Kulshreshtha K, Agnihotri S. Seasonal dynamics of methane emission from wetland. Chemosphere-Global Change Science, 2000, 2(1):39-46.
[77] Wang F S, Wang B L, Liu C Q, Wang Y C, Guan J, Liu X L, Yu Y X. Carbon dioxide emission from surface water in cascade reservoirs-river system on the Maotiao River, southwest of China. Atmospheric Environment, 2011, 45(23):3827-3843.
[78] Hope D, Kratz T K, Riera J L. Relationship betweenpCO2and dissolved organic carbon in northern Wisconsin lakes. Journal of Environmental Quality, 1996, 25(6):1442-1445.
[79] Huttunen J T, Lappalainen K M, Saarijǎrvi E, Väisänen T, Martikainen P J. A novel sediment gas sampler and a subsurface gas collector used for measurement of the ebullition of methane and carbon dioxide from a eutrophied lake. The Science of the Total Environment, 2001, 266(1/3):153-158.
[80] 杨平, 仝川, 何清华, 黄佳芳. 闽江口鱼虾混养塘水-气界面温室气体通量及主要影响因子. 环境科学学报, 2013, 33(5):1493-1503.
[81] Conrad R, Rothfuss F. Methane oxidation in the soil surface layer of a flooded rice field and the effect of ammonium. Biology and Fertility of Soils, 1991, 12(1):28-32.
[82] 袁淑方, 王为东. 太湖流域源头溪流氧化亚氮(N2O)释放特征. 生态学报, 2012, 32(20):6279-6288.
[83] Usui T, Koike I, Ogura N. N2O production, nitrification and denitrification in an estuarine sediment. Estuarine, Coastal and Shelf Science, 2001, 52(6):769-781.
[84] Stow C A, Walker J T, Cardoch L, Spence P, Geron C. N2O emissions from streams in the Neuse river watershed, North Carolina. Environmental Science & Technology, 2005, 39(18):6999-7004.
[85] Clough T J, Buckthought L E, Casciotti K L, Kelliher F M, Jones P K. Nitrous oxide dynamics in a braided river system, New Zealand. Journal of Environmental Quality, 2011, 40(5):1532-1541.
[86] 王洪君, 王为东, 卢金伟, 杨龙元, 尹澄清. 太湖湖滨带秋、冬季CH4排放特征及其影响因素初步研究. 湿地科学, 2006, 4(1):21-28.
[87] Purvaja R, Ramesh R, Frenzel P. Plant-mediated methane emission from an Indian mangrove. Global Change Biology, 2004, 10(11):1825-1834.
[88] Hyvönen T, Ojala A, Kankaala P, Martikainen P J. Methane release from stands of water horsetail (Equisetumfluviatile) in a boreal lake. Freshwater Biology, 1998, 40(2):275-284.
[89] Roura-Carol M, Freeman C. Methane release from peat soils:effects ofSphagnumandJuncus. Soil Biology & Biochemistry, 1999, 31(2):323-325.
[90] Keppler F, Hamilton J T G, Brass M, Röckmann T. Methane emissions from terrestrial plants under aerobic conditions. Nature, 2006, 439(7073):187-191.
[91] Zhao Y, Wu B F, Zeng Y. Spatial and temporal patterns of greenhouse gas emissions from Three Gorges Reservoir of China. Biogeosciences, 2013, 10:1219-1230.
[92] 杨健, 苏彦平, 刘洪波, 戈贤平. 内陆渔业生态系统的碳循环特征及碳汇机制. 水产学报, 2012, 36(5):794-800.
[93] Stallard R F. Terrestrial sedimentation and the carbon cycle:Coupling weathering and erosion to carbon burial. Global Biogeochemical Cycles, 1998, 12(2):231-257.
[94] 姚冠荣, 高全洲. 河流碳输移与陆地侵蚀-沉积过程关系的研究进展. 水科学进展, 2007, 18(1):133-139.
[95] 刘丛强, 汪福顺, 王雨春, 王宝利. 河流筑坝拦截的水环境响应-来自地球化学的视角. 长江流域资源与环境, 2009, 18(4):384-396.
[96] 陈庆伟, 刘兰芬, 刘昌明. 筑坝对河流生态系统的影响及水库生态调度研究. 北京师范大学学报(自然科学版), 2007, 43(5):578-582.
[97] Boyd C E, Wood C W, Chaney P L, Queiroz J F. Role of aquaculture pond sediments in sequestration of annual global carbon emissions. Environmental Pollution, 2010, 158(8):2537-2540
Emission paths and measurement methods for greenhouse gas fluxes from freshwater ecosystems:a review
YANG Ping1,2, TONG Chuan1,3,4,*
1KeyLaboratoryofHumidSub-tropicalEco-geographicalProcessofMinistryofEducation,Fuzhou350007,China2SchoolofGeographicalSciences,FujianNormalUniversity,Fuzhou350007,China3InstituteofGeography,FujianNormalUniversity,Fuzhou350007,China4ResearchCentreofWetlandsinSubtropicalRegion,FujianNormalUniversity,Fuzhou350007,China
Greenhouse gas (GHGs) emissions from freshwater ecosystems are a major component of global terrestrial landscape budgets. Currently, global warming is affecting these ecosystems and may trigger an increase in GHGs emissions, which may further enhance global warming. The identification and accurate quantification of aquatic ecosystems as sinks/sources of GHGs are vital for evaluating GHGs budgets and assessing possible climate feedback effects in order to improve climate models. In recent years, fluxes of carbon dioxide (CO2), methane (CH4), and nitrous oxide (N2O) have been observed from freshwater aquatic environments such as natural lakes, hydropower reservoirs, rivers, ponds, and drainage ditches. This review analyzes and summarizes research developments in GHGs emission paths, observation methods, and key factors affecting emissions from freshwater ecosystems. The mechanism of greenhouse gas production is a complex and interactive process that includes biochemical processes. The main emission paths from aquatic environments are diffusive fluxes across the air-water interface, bubble (ebullition) fluxes resulting from supersaturation of sediment, and plant-mediated fluxes. Attention has been recently been drawn to other emission pathways that contribute to total gas emissions at reservoir surfaces (e.g., gas release immediately below turbines and emissions further downstream in rivers). The monitoring methods vary for aquatic ecosystem emission pathways. Bubble fluxes are measured by funnel techniques, open dynamic floating methods, and ultrasonic detection technologies. Diffusive fluxes are measured by static chamber techniques, model estimations, micrometeorology, and tunable diode laser absorption spectroscopy (TDLAS). GHGs emission is conventionally measured using closed chamber to trap plant-mediated flux components. In addition, we discuss the impacts of physical, biological, hydrodynamic, and anthropogenic factors on GHGs emissions from aquatic ecosystems. We point out that an urgent and key direction for the future is to standardize the observation methods for GHGs fluxes from freshwater aquatic ecosystems and to consider temporal and spatial variability, which rely on long-term field observation.
fresh-water aquatic ecosystems; greenhouse gases; emission paths; measure method; influencing factors
国家自然科学基金资助项目(41371127); 福建师范大学校级创新团队项目(IRTL1205)
2014-06-23; < class="emphasis_bold">网络出版日期:
日期:2014-12-18
10.5846/stxb201406231298
*通讯作者Corresponding author.E-mail: tongch@fjnu.edu.cn
杨平, 仝川.淡水水生生态系统温室气体排放的主要途径及影响因素研究进展.生态学报,2015,35(20):6868-6880.
Yang P, Tong C.Emission paths and measurement methods for greenhouse gas fluxes from freshwater ecosystems:a review.Acta Ecologica Sinica,2015,35(20):6868-6880.
