土壤微生物在植物获得养分中的作用
2015-01-20沈仁芳赵学强
沈仁芳,赵学强
土壤与农业可持续发展国家重点实验室,中国科学院南京土壤研究所,南京 210008
土壤微生物在植物获得养分中的作用
沈仁芳*,赵学强
土壤与农业可持续发展国家重点实验室,中国科学院南京土壤研究所,南京 210008
大量施用化肥是当今农业的一个重要特征。化肥为粮食增产做出了巨大贡献,同时也带来一系列问题,如土壤酸化、水体富营养化、温室气体排放、资源耗竭等,直接威胁着农业可持续发展。土壤微生物是陆地生态系统植物多样性和生产力的重要驱动者,直接参与了植物获得养分和土壤养分循环两个过程。因此,通过调控土壤微生物的功能,有望降低农业对化肥的过分依赖。介绍了共生固氮菌、菌根真菌和根际促生菌对植物获得养分能力的影响及其机制,分析了土壤微生物对土壤氮、磷循环的影响及其与土壤养分生物有效性、养分损失的关系。依据这些知识,提出了改善植物营养、降低化肥施用的土壤微生物途径。虽然大量试验已证明了土壤微生物在改善植物营养中的重要作用,但是大面积应用土壤微生物技术来改善植物营养还存在不少问题。随着以后对这方面研究的加强以及上述问题的不断解决,土壤微生物有望在降低化肥施用量和维持农业可持续发展中做出重要贡献。
生物固氮;菌根真菌;根际促生菌;养分循环;农业可持续发展
土壤是地球的皮肤,是陆地表面能够生长植物的疏松层[1-2]。土壤学是科学研究领域的最后前沿之一[3],土壤生物学是土壤学研究领域的前沿[2]。土壤是地球上生物多样性的最大储库[4],微生物是土壤生物多样性体现的主要表现形式。依据土壤的定义,其本质属性是能为植物提供养分,即植物营养功能,土壤微生物在土壤为植物提供养分过程中起着关键作用。据粗略估计,在养分贫瘠的自然土壤上,至少2万个植物种类的生存离不开土壤微生物[5],约72%植物离不开菌根真菌[6],约25%植物离不开固氮菌[7]。土壤微生物是陆地生态系统植物多样性和生产力的重要驱动者[5],地上部植物生长和地下微生物之间有着密切联系[8]。可以想象,假如没有土壤微生物,很多自然土壤可能会是“不毛之地”。
虽然一般承认土壤微生物对自然生态系统植物营养有重要作用,但是土壤微生物在农田生态系统植物获得养分中的作用经常被低估或忽视,其主要原因是化肥的大量施用满足了农田作物的营养需求。不可否认,化肥对世界粮食增产有重要贡献。据估计,20世纪40%世界人口依赖于化肥增产作用,30%—50%粮食增产是氮肥施用的结果[9]。21世纪中叶世界人口将达到90亿,粮食产量只有翻一番才能养活这个世界[10-11]。在耕地面积减少的现状下,如果没有科学技术上的重大突破,意味着可能需要增施更多化肥才能提高产量。然而,化肥的大量施用使得人们正在忽视土壤微生物的功能,甚至有时会怀疑土壤微生物对植物营养的贡献,因为人们会认为只要向土壤中施入足量化肥,就可以获得高产,土壤微生物存在与否,表面上并没有显著改变作物产量。化肥的大量施用正在引起许多负面环境效应,如温室气体的排放、水体富营养化、土壤酸化、土壤生物多样性下降等,直接威胁到农业的可持续发展[12-15]、甚至人类的生存[16]。化肥的大量施用可能掩盖甚至阻碍了土壤微生物对植物营养的影响和贡献,重新探讨土壤微生物在植物获得养分中的作用途径及其机理,有助于降低农田化肥施用量。
本文通过分析土壤微生物对植物获得养分能力和土壤养分循环的影响,探寻改善植物营养的土壤微生物途径,并评价这些途径的可行性,以期达到“增强作物养分吸收、提高作物产量、降低化肥施用量”的目的。虽然目前还有许多问题等着去解决,但是一些研究已证明了这一思路的可行性[17-20],。由于氮和磷是植物两大必需元素,而且二者具有显著的环境效应,这里主要考虑氮和磷。
1 土壤微生物对植物获得养分能力的影响——直接作用
一些土壤微生物能够与植物根系形成共生关系,或者分布在植物根际,影响植物获得土壤养分的能力,这里称之为土壤微生物对植物营养的直接作用。这类微生物主要有共生固氮菌、丛枝菌根真菌、植物根际促生菌。一方面它们帮助植物获得土壤养分,另一方面植物为它们的生长提供碳源,形成了一种互惠互利的关系。
1.1 共生固氮菌
虽然大气中含有80%左右的氮,但是这种分子态氮是惰性的,不能被植物直接利用。氮气必须被合成为氨后,才能被植物利用。氨合成分为工业合成和生物合成。氨的工业合成方式存在耗能高、环境污染、代价高等缺点。生物固氮是一种自然的氨合成方式,是农业可持续发展的一个高效氮源[21]。据联合国粮农组织1995年粗略估计, 全球每年生物固氮量已近2×106t(相当于4×108t尿素),约占全球植物需氮量的3/4,所以生物固氮是地球上最大规模的天然氮肥工厂[22]。然而,这一巨大的生物固氮量对农田生态系统意义却不大,因为大多数农田(如种植水稻、小麦和玉米)生物固氮量极微[22]。自然界固氮菌种类繁多,现己发现分布在100多个菌属,最早的共生固氮蓝细菌可以追溯到2.5亿年前[23]。根据固氮菌与植物的疏密关系,分为共生固氮、联合固氮和自生固氮三个体系[24]。共生固氮体系中有根瘤菌-豆科植物共生、弗兰克氏菌-非豆科植物共生等, 其中以根瘤菌-豆科植物共生体系固氮能力最强, 年固氮量占生物固氮总量的60%以上[24]。根瘤菌和豆科植物共生固氮量能够达到植物需氮量的90%以上[25]。目前,在根瘤菌资源收集、分离和分类、固氮酶结构和功能、固氮基因和结瘤基因鉴定、根瘤菌和固氮植物基因组测序和根瘤菌-根共生机制等多个方面,已取得多项突破进展。大量的报道也已表明,接种固氮菌能够提高作物产量,增强植物对氮素的吸收[17]。
1.2 菌根真菌
菌根是一种最为古老的高等植物与微生物共生的现象[2]。丛枝菌根是菌根中最常见的一种类型,其形成可以追溯到4亿年前[26]。1885年,德国植物生理学家Frank首次用mycorrhiza来描述植物根与真菌形成的共生体,至今,菌根研究历史已近130年[2]。根据共生体结构特征,菌根包括外生菌根、内生菌根和一些特殊菌根类型,如兰科菌根和杜鹃科菌根[2]。近85%的植物科属和几乎所有农作物能够形成内生菌根,而外生菌根主要分布于树木根系[2,26]。丛枝菌根真菌能为植物贡献高达90%的磷[27]。菌根真菌最主要的植物营养功能是促进植物对土壤磷的摄取能力,其机制主要是通过菌丝扩大养分吸收空间、活化土壤有机和无机磷来得以实现[28]。越来越多的证据表明,丛枝菌根真菌还能增强植物对氮的吸收[29]。据估计,北方森林植物约80%氮来源于外生菌根真菌[5]。菌根真菌在植物吸收氮素中的作用及其机制正成为一个新的研究热点[30-32]。菌根真菌还能增强植物吸收微量元素铁、锌、铜等[28,33]。一般认为丛枝菌根真菌主要在低磷土壤上发挥作用,而在高磷土壤上,丛枝菌根真菌甚至能够抑制植物生长,这可能由于真菌寄生消耗了大量碳源,而高磷土壤植物并不需要菌根来增强植物对磷的吸收[34]。近些年来,我国菜地土壤有效磷含量富集严重,一些田块土壤有效磷能够达到上百mg/kg,远高于土壤磷流失风险的临界值[35-36]。有趣的是,在特定的条件下,丛枝菌根真菌在高磷土壤上仍能提高一些蔬菜对磷和氮的吸收利用[37-38]。
1.3 植物根际促生菌
根际一般指距根轴表面数毫米范围内的微区域,是土壤-根系-微生物相互作用的活跃区域,在物理、化学和生物学性质上显著不同于土体[1]。根际是植物吸收养分的门户,植物根际促生菌是分布在根际中的一类有益微生物。有别于共生固氮菌和菌根真菌,这类微生物一般不与植物形成共生关系。1978年,美国奥本大学Kloepper首次提出Plant Growth-Promoting Rhizobacteria(PGPR)的概念,指一群定值于植物根际,能够促进植物生长的一类细菌代表[2]。目前已发现的植物根际促生菌包括假单胞菌属、芽孢杆菌属、农杆菌属、黄杆菌属、沙雷氏菌等20多个种属,其中荧光假单胞菌被认为最具有应用前景。植物根际促生菌的促生机制已比较清楚,包括:(1)产生激素或信号分子,如生长素、赤霉素、细胞分裂素、挥发性物质、一氧化氮,或者通过ACC脱氨酶,阻止植物乙烯的合成;(2)溶解土壤难溶性磷;(3)产生铁载体,提高土壤铁有效性;(4)非共生固氮;(5)作为“助手细菌”,影响根瘤菌、丛枝菌根真菌与植物的共生;(6)增强植物对病原菌的抗性和忍耐力等[2,25,39]。接种根际促生菌能够降低水稻化肥施用量的50%,同时维持水稻产量[40]。
近年来,人们正在把目光转向植物根际促生菌、固氮菌、丛枝菌根真菌的联合接种效应研究。例如,在田间条件下,联合接种菌根真菌和根际促生菌能够显著减少化肥施用量,提高化肥利用率[17,41-42]。
2 土壤微生物对土壤养分循环的影响——间接作用
一方面,土壤微生物可以通过改变植物根系生理和根际环境,直接影响植物获得养分的能力;另一方面,非根际土壤(即土体)中的土壤养分循环,如养分的生物固定和矿化分解、硝化和反硝化等过程,都离不开微生物的参与。土壤微生物是土壤肥力形成和持续发展的核心动力,土壤有机质主要靠微生物来分解, 养分的释放量很大程度上受控于微生物生物量,养分的矿化和转化推动着养分循环,影响着养分的生物有效性[5,24,43]。从微生物本身考虑,微生物参与土壤养分循环的目的并不是为了给植物提供养分,微生物甚至与植物竞争养分,但是微生物对土壤养分循环的影响改变了土壤养分有效性,从而间接地影响了植物营养。这种间接作用可能带来正负两种效果:正效应,提高了土壤养分有效性,改善植物营养;负效应,提高了养分进入环境的风险,如流失到水体,或挥发到大气。
2.1 土壤微生物对土壤氮循环的影响
与土壤微生物直接有关的氮循环过程主要有生物固氮、氮矿化、硝化、反硝化等。其中生物固氮在前面已有介绍。不过,除了共生固氮外,自然界土壤中还存在大量的自生固氮菌,这部分氮对增加土壤氮储库也具有不可忽视的作用。虽然植物能够吸收一些小分子量有机氮源,但是大部分有机氮源对植物是无效的,这些有机氮必须被矿化分解为小分子氮或氨后,才能被植物利用。表征土壤氮有效性的常用指标也是碱解氮。氮矿化是土壤氮素循环的核心,控制着土壤氮素的生物有效性,这一过程受到多种土壤微生物的调控[44]。生物合成的氨、肥料施入的氨以及矿化产生的氨,都会进入下一个氮素循环环节-硝化过程,将铵态氮转化为硝态氮。氨氧化细菌和氨氧化古菌在土壤硝化过程中发挥主要作用[45]。与硝化作用相偶联,土壤中的硝酸根在土壤微生物的参与下,通过还原反应,硝酸根最终被还原成氮气,并在中间过程释放强致热效应的温室气体氧化亚氮[2]。
上述四个氮素循环过程中,生物固氮会增加土壤有效氮含量,提高植物氮营养。氮矿化对氮的影响可能是双方向的,一方面短期会增加土壤有效氮,增加土壤矿质态氮,能够改善植物生长[46];但是如果矿化过度(超过作物需求),也会降低土壤有效氮库存,长期效应可能是负的。有趣的是,一些报道表明丛枝菌根真菌能够改变土壤微生物,进而影响土壤氮矿化,提高植物对氮的吸收[29,47-48]。关于硝化作用,通过添加硝化抑制剂,抑制土壤中硝化作用的微生物过程,可以提高氮肥利用率,减少氮向水体和空气中的排放[49]。另外,硝化作用还与过量氮肥投入导致的土壤酸化、硝酸盐淋溶和氧化亚氮排放等环境问题直接相关[2]。然而,完全抑制土壤硝化作用会带来一些负作用,对于“喜”硝植物,硝化作用的抑制可能会导致铵毒的产生,抑制植物生长。无论从植物营养还是环境效应考虑,反硝化作用均是有害无益的氮素循环过程,应当阻止该过程。因此,土壤氮素循环的微生物调控应该以“提高生物固氮、适当刺激矿化、调节硝化、控制反硝化”为原则。
2.2 土壤微生物对土壤磷循环的影响
不同于氮,植物带走的磷只能通过施肥或活化土壤本身的磷来补充。虽然土壤总磷含量较高,但是磷在土壤中移动性差及易被土壤固定,仅0.1%土壤总磷对植物有效。从植物营养角度考虑,土壤磷循环主要包括磷的吸附和解吸、微生物固磷和分解、有机磷矿化等三个过程。解吸土壤吸附态磷是提高土壤磷有效性的常用方法。据估计,解磷细菌占实验室可培养细菌的40%左右[50],有些解磷菌属于根际促生菌。根系分泌物能够提高土壤解磷菌的活性,增加土壤磷的供应[51]。在温室和田间条件下,接种解磷菌均提高了植物吸磷能力[18]。微生物能将无机磷转化为微生物磷,即磷的微生物固定。土壤微生物也能够快速分离植物残体和有机质中的磷,合成为微生物磷。土壤微生物磷占土壤总磷约2%—10%,有时候甚至超过了植物体磷含量[18]。土壤微生物磷是土壤最具生物活性的磷,它能根据土壤有效磷的丰缺状况,进行固持和转化,满足植物生长需要[18]。土壤有机磷含量很高,与氮相似,植物一般也不能利用有机磷,需要微生物将土壤有机磷矿化为无机磷后,才能被植物吸收。土壤有机磷矿化主要依赖于土壤微生物过程[25]。土壤微生物通过各种过程(如分泌磷酸酶),将有机磷水解为磷酸根和其他小分子含磷化合物,供植物吸收利用。
上述三个土壤磷循环过程的最终效果均是双方向的,既可以提高土壤磷的生物有效性,又可以降低土壤有效磷库,或者增加磷向环境的迁移。例如,土壤大量吸附态磷被解吸到土壤溶液,大部分微生物磷被分解,大部分有机磷被矿化,这些过程短期内增加了土壤磷有效性,但是土壤供磷有可能超过了作物需求,多余的磷会移入周围水体,形成负面效应。另外,土壤微生物磷是土壤有效磷的补充,但是在缺磷条件下,微生物也会和植物竞争磷。土壤既是磷源,也是磷库,土壤中的磷与土壤溶液中的磷存在一个动态平衡,这一平衡向任何方向过度偏斜,可能都是不利的。根据植物营养需求,利用土壤微生物的手段,调控这个动态平衡,是土壤磷素循环的微生物调控原则。
3 改善植物营养的土壤微生物途径、问题与展望
3.1 改善植物营养的土壤微生物途径
根据2.1和2.2的分析,这里建议将改善植物营养的土壤微生物途径分为三类(图1):(1)增强土壤固氮菌、菌根真菌、根际促生菌的功能,提高植物本身对氮、磷和微量元素的吸收,包括图1中①、②、③、④途径;(2)平衡土壤微生物对氮磷的固定与分解,微生物过度固定或分解氮磷都不可取,指图1途径⑤;(3)根据植物营养需求,适当增强氮磷矿化微生物、解磷菌和硝化微生物的功能,促进养分向植物根系迁移,减少甚至阻止氮、磷向水体和大气的排放,包括图1中⑥、⑦、⑧途径。上述途径综合考虑了微生物对植物营养的正负效应,增强了植物对养分的吸收,降低了养分向环境的排放,实现了“既降低化肥施用量、又减少养分环境效应”的双重目的。
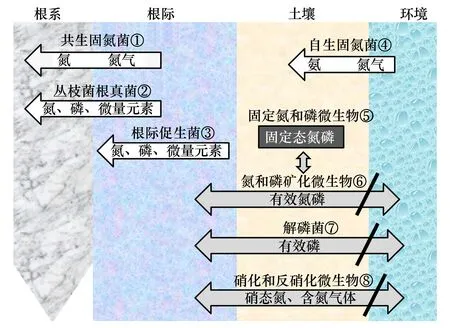
图1 改善植物营养的土壤微生物途径Fig.1 Soil microbial processes to improve plant nutrition
3.2 存在问题
虽然很多研究证明微生物对于改善植物营养具有重要作用,但是在农业上,这些技术和产品目前仍没有实现大面积的应用和推广。存在的主要问题有:
(1)菌种扩繁和接种困难、与土著微生物存在竞争。例如,菌根真菌必须与植物共生才能生存,脱离宿主植物的菌根真菌纯培养很难成功,导致接种菌根真菌不能大面积推广,极大地限制了菌根真菌机制研究及其应用[28]。接种到土壤中的微生物,有时候并不能适应土壤条件,也不能竞争过土著微生物,导致它们不能发挥作用。
(2)大田作物应用效果不佳。在粮食作物水稻、小麦、玉米、多种果树、蔬菜上,大多数固氮微生物不能固氮,即使少数能够固氮,固氮量也很低, 所以这些作物的高产仍依赖于大量氮肥的投入[22]。
(3)微生物效果不稳定。虽然很多微生物接种试验在盆栽条件下获得很好效果,但是在田间试验条件下经常表现不出效果。有时候在一种土壤条件上有效果,在另外一种土壤条件下看不到效果。
(4)研究方法上也存在很大困难。土壤本身具有多种微生物类群,为了证明接种微生物的功能,经常会采用土壤灭菌的方法,但是灭菌的方法又会带来土壤生物、化学和物理等性质的改变,导致最后看到的实验效果不知道是微生物的作用还是土壤性质改变引起的。
(5)微生物肥料目前并没有一个严格可操作的定义和执行标准。由于我国《肥料法》的缺乏[52],这导致市场上微生物肥料种类繁多,质量参差不齐,价格昂贵,施用过程中有时候并不能达到说明书上的效果,这也影响了人们对微生物肥料的认识和阻碍了微生物肥料的推广。
3.3 研究展望
随着化肥环境效应和资源短缺问题的日益加剧,人类必须寻找新的方法去减少化肥施用量,以维持农业的可持续发展和自身的生存。可以想象,在化肥施用之前的年代,微生物在植物获得土壤养分过程中发挥了重要作用。即便当今,在一些原始森林和草原,植物的营养应该仍主要依赖于土壤微生物的作用。这为通过调控土壤微生物来改善植物营养提供了重要信息。因为人们从农田带走了大量农产品,根据“养分归还学说”,就必须向土壤归还带走的养分。因此,微生物方法不能完全代替化肥的施用,但是可以充分利用土壤微生物与植物营养的关系,来减少化肥施用量,在一定程度上实现土壤养分的良性循环。自然生态系统土壤微生物已经而且正在发挥着改善植物营养的功能,所以对于自然生态系统,唯一能做的就是不去破坏土壤微生物的功能。现在面临的主要挑战是如何发挥农田生态系统土壤微生物在植物获得养分中的功能,达到降低化肥用量、提高化肥利用率的目的。
依据上述分析,建议将来研究方向集中在“农田生态系统降低化肥用量的土壤微生物途径研究”。围绕这一方向,建议优先开展以下几方面研究:(1)进一步从自然和农田生态系统筛选、分离和鉴定增强植物营养的高效菌株,并验证这些菌株在农田生态系统中的作用,分析它们起作用的条件;(2)深入剖析固氮菌、丛枝菌根真菌、根际促生菌改善植物营养的关键机制,特别是分子生物学机制,寻找土壤微生物与植物营养之间的“对话信号”,寻求将这些分子机制和对话信号用于改善农田生态系统作物营养的途径;(3)目前大多微生物接种试验是在盆栽条件下开展的,应进一步扩大微生物接种田间试验效果的研究,寻找微生物田间效果不稳定的原因;(4)比较单一微生物效应和微生物群体效应,研究固氮菌、丛枝菌根真菌、根际促生菌之间的相互作用,研究多种微生物在增强植物获得氮、磷及其他养分方面的协同效应;(5)由于目前我国许多农田养分含量偏高,而大多数微生物是在低养分条件下发挥作用,所以需要进一步探讨土壤微生物在高养分土壤农田作物上的施用效果及限制因子,这对于减少化肥施用、降低养分流失具有重要意义。
虽然微生物大范围应用在改善植物营养上还有很长的路要走,但是可喜的是,越来越多的证据表明接种微生物能够降低土壤氮、磷流失和温室气体排放[20,53-55],这极大地增强了人们利用微生物途径改善植物营养的信心。除了改善植物营养外,微生物在土壤重金属和有机污染修复、土传病害防治、抵抗酸性土壤等多个方面具有重要作用。土壤微生物是农业可持续发展的基础,未来的农业有望通过调控土壤微生物,提高作物产量、降低化肥和农药施用,并提高农产品品质[56]。可见,土壤微生物使人类的未来并不可怕。
[1] 周健民, 沈仁芳. 土壤学大辞典. 北京: 科学出版社, 2013.
[2] 贺纪正, 陆雅海, 傅伯杰. 土壤生物学前沿. 北京: 科学出版社, 2015.
[3] Sugden A, Stone R, Ash C. Ecology in the underworld. Science, 2004, 304(5677): 1613-1613.
[4] Crawford J W, Harris J A, Ritz K, Young I M. Towards an evolutionary ecology of life in soil. Trends in Ecology & Evolution, 2005, 20(2): 81-87.
[5] Van Der Heijden M G A, Bardgett R D, Van Straalen N M. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology Letters, 2008, 11(3): 296-310.
[6] Van Der Heijden M G A, Klironomos J N, Ursic M, Moutoglis P, Streitwolf-Engel R, Boller T, Wiemken A, Sanders I R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature, 1998, 396(6706): 69-72.
[7] Van Der Heijden M G A, Bakker R, Verwaal J, Scheublin T R, Rutten M, Van Logtestijn R, Staehelin C. Symbiotic bacteria as a determinant of plant community structure and plant productivity in dune grassland. FEMS Microbiology Ecology, 2006, 56(2): 178-187.
[8] Wardle D A, Bardgett R D, Klironomos J N, Setälä H, Van Der Putten W H, Wall D H. Ecological linkages between aboveground and belowground biota. Science, 2004, 304(5677): 1629-1633.
[9] Erisman J W, Sutton M A, Galloway J, Klimont Z, Winiwarter W. How a century of ammonia synthesis changed the world. Nature Geoscience, 2008, 1(10): 636-639.
[10] Godfray H C J, Beddington J R, Crute I R, Haddad L, Lawrence D, Muir J F, Pretty J, Robinson S, Thomas S M, Toulmin C. Food security: the challenge of feeding 9 billion people. Science, 2010, 327(5967): 812-818.
[11] Tilman D, Balzer C, Hill J, Befort B L. Global food demand and the sustainable intensification of agriculture. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(50): 20260-20264.
[12] Zhu Z L, Chen D L. Nitrogen fertilizer use in China-Contributions to food production, impacts on the environment and best management strategies. Nutrient Cycling in Agroecosystems, 2002, 63(2/3): 117-127.
[13] 沈仁芳. 铝在土壤-植物中的行为及植物的适应机制. 北京: 科学出版社, 2008.
[14] Foley J A, Ramankutty N, Brauman K A, Cassidy E S, Gerber J S, Johnston M, Mueller N D, O′Connell1 C, Ray D K, West P C, Balzer C, Bennett E M, Carpenter S R, Hill J, Monfreda C, Polasky S, Rockström J, Sheehan J, Siebert S, Tilman D, Zaks D P M. Solutions for a cultivated planet. Nature, 2011, 478(7369): 337-342.
[15] Zhao X Q, Chen R F, Shen R F. Coadaptation of plants to multiple stresses in acidic soils. Soil Science, 2015, 179(10-11): 503-513.
[16] Amundson R, Berhe A A, Hopmans J W, Olson C, Sztein A E, Sparks D L. Soil and human security in the 21st century. Science, 2015, 348(6235): 1261071.
[17] Adesemoye A O, Kloepper J W. Plant-microbes interactions in enhanced fertilizer-use efficiency. Applied Microbiology and Biotechnology, 2009, 85(1): 1-12.
[18] Richardson A E, Simpson R J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiology, 2011, 156(3): 989-996.
[19] Bender S F, Conen F, Van Der Heijden M G A. Mycorrhizal effects on nutrient cycling, nutrient leaching and N2O production in experimental grassland. Soil Biology Biochemistry, 2015, 80: 283-292.
[20] Cavagnaro T R, Bender S F, Asghari H R, Van Der Heijden M G A. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends in Plant Science, 2015, 20(5): 283-290.
[21] Peoples M B, Herridge D F, Ladha J K. Biological nitrogen fixation: an efficient source of nitrogen for sustainable agricultural production?. Plant and Soil, 1995, 174(1/2): 3-28.
[22] 沈世华, 荆玉祥. 中国生物固氮研究现状和展望. 科学通报, 2003, 48(6): 535-540.
[23] Raven J A. Evolution of cyanobacterial symbioses // Rai A N, Bergman B, Rasmussen U, eds. Cyanobacteria in Symbiosis. Netherlands: Kluwer Academic Publishers, 2002: 329-346.
[24] 陈文新, 李阜棣, 闫章才. 我国土壤微生物学和生物固氮研究的回顾与展望. 世界科技研究与发展, 2002, 24(4): 6-12.
[25] Richardson A E, Barea J -M, McNeill A M, Prigent-Combaret C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant and Soil, 2009, 321(1/2): 305-339.
[26] Lambers H, Mougel C, Jaillard B, Hinsinger P. Plant-microbe-soil interactions in the rhizosphere: an evolutionary perspective. Plant and Soil, 2009, 321(1/2): 83-115.
[27] Van Der Heijden M G A, Streitwolf-Engel R, Riedl R, Siegrist S, Neudecker A, Ineichen K, Boller T, Wiemken A, Sanders I R. The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland. New Phytologist, 2006, 172(4): 739-752.
[28] 李晓林, 姚青. VA菌根与植物的矿质营养. 自然科学进展, 2000, 10(6): 524-531.
[29] Hodge A, Campbell C D, Fitter A H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature, 2001, 413(6853): 297-299.
[30] Veresoglou S D, Chen B D, Rillig M C. Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biology and Biochemistry, 2012, 46: 53-62.
[31] Jin H R, Liu J, Liu J, Huang X W. Forms of nitrogen uptake, translocation, and transfer via arbuscular mycorrhizal fungi: a review. Science China Life Sciences, 2012, 55(6): 474-482.
[32] Hodge A, Storer K. Arbuscular mycorrhiza and nitrogen: implications for individual plants through to ecosystems. Plant and Soil, 2015, 386(1/2): 1-19.
[33] Pellegrino E, Bedini S. Enhancing ecosystem services in sustainable agriculture: Biofertilization and biofortification of chickpea (CicerarietinumL.) by arbuscular mycorrhizal fungi. Soil Biology and Biochemistry, 2014, 68: 429-439.
[34] Peng S, Eissenstat D M, Graham J H, Williams K, Hodge N C. Growth depression in mycorrhizal citrus at high phosphorus supply (Analysis of carbon costs). Plant Physiology, 1993, 101(3): 1063-1071.
[35] 赵学强, 沈仁芳. 我国菜地土壤氮磷钾养分状况与均衡调控 // 江苏省土壤学会编. 江苏耕地质量建设论文集. 南京: 河海大学出版社, 2008: 97-101.
[36] Liang L Z, Shen R F, Yi X Y, Zhao X Q, Chen Z C, Chen R F, Dong X Y. The phosphorus requirement ofAmaranthusmangostanusL. exceeds the “change point” of P loss. Soil Use and Management, 2009, 25(2): 152-158.
[37] Xu P, Liang L Z, Dong X Y, Shen R F. Effect of arbuscular mycorrhizal fungi on aggregate stability of a clay soil inoculating with two different host plants. Acta Agriculturae Scandinavica, Section B-Soil and Plant Science, 2015, 65(1): 23-29.
[38] Xu P, Liang L Z, Dong X Y, Xu J, Jiang P K, Shen R F. Response of soil phosphorus required for maximum growth ofAsparagusofficinalisL. to inoculation of arbuscular mycorrhizal fungi. Pedosphere, 2014, 24(6): 776-782.
[39] 康贻军, 程洁, 梅丽娟, 胡健, 朴哲, 殷士学. 植物根际促生菌作用机制研究进展. 应用生态学报, 2010, 21(1): 232-238.
[40] Jetiyanon K, Plianbangchang P. Potential ofBacilluscereusstrain RS87 for partial replacement of chemical fertilisers in the production of Thai rice cultivars. Journal of the Science of Food and Agriculture, 2012, 92(5): 1080-1085.
[41] Adesemoye A O, Torbert H A, Kloepper J W. Enhanced plant nutrient use efficiency with PGPR and AMF in an integrated nutrient management system. Canadian Journal of Microbiology, 2008, 54(10): 876-886.
[42] Adesemoye A O, Torbert H A, Kloepper J W. Plant growth-promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microbial Ecology, 2009, 58(4): 921-929.
[43] Kuzyakov Y, Friedel J K, Stahr K. Review of mechanisms and quantification of priming effects. Soil Biology and Biochemistry, 2000, 32(11-12): 1485-1498.
[44] Schimel J P, Bennett J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology, 2004, 85(3): 591-602.
[45] Che J, Zhao X Q, Zhou X, Jia Z J, Shen R F. High pH-enhanced soil nitrification was associated with ammonia-oxidizing bacteria rather than archaea in acidic soils. Applied Soil Ecology, 2015, 85: 21-29.
[46] Unkovich M, Jamieson N, Monaghan R, Barraclough D. Nitrogen mineralisation and plant nitrogen acquisition in a nitrogen-limited calcareous grassland. Environmental and Experimental Botany, 1998, 40(3): 209-219.
[47] Herman D J, Firestone M K, Nuccio E, Hodge A. Interactions between an arbuscular mycorrhizal fungus and a soil microbial community mediating litter decomposition. FEMS Microbiology Ecology, 2012, 80(1): 236-247.
[48] Nuccio E E, Hodge A, Pett-Ridge J, Herman D J, Weber P K, Firestone M K. An arbuscular mycorrhizal fungus significantly modifies the soil bacterial community and nitrogen cycling during litter decomposition. Environmental Microbiology, 2013, 15(6): 1870-1881.
[49] Shoji S, Delgado J, Mosier A, Miura Y. Use of controlled release fertilizers and nitrification inhibitors to increase nitrogen use efficiency and to conserve air and water quality. Communications in Soil Science and Plant Analysis, 2001, 32(7/8): 1051-1070.
[50] Kucey R M N. Phosphate-solubilizing bacteria and fungi in various cultivated and virgin Alberta soils. Canadian Journal of Soil Science, 1983, 63(4): 671-678.
[51] Richardson A E. Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Australian Journal of Plant Physiology, 2001, 28(9): 897-906.
[52] 沈仁芳, 梁林洲, 赵学强. 日本、韩国、台湾地区肥料相关的立法及其对我国肥料管理的启示. 土壤, 2012, 44(4): 529-534.
[53] Govindarajulu M, Pfeffer P E, Jin H R, Abubaker J, Douds D D, Allen J W, Bücking H, Lammers P J, Shachar-Hill Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature, 2005, 435(7043): 819-823.
[54] Bender S F, Plantenga F, Neftel A, Jocher M, Oberholzer H R, Köhl L, Giles M, Daniell T J, Van Der Heijden M G A. Symbiotic relationships between soil fungi and plants reduce N2O emissions from soil. The ISME Journal, 2014, 8(6): 1336-1345.
[55] Bender S F, Van Der Heijden M G A. Soil biota enhance agricultural sustainability by improving crop yield, nutrient uptake and reducing nitrogen leaching losses. Journal of Applied Ecology, 2015, 52(1): 228-239.
[56] Tikhonovich I A, Provorov N A. Microbiology is the basis of sustainable agriculture: an opinion. Annals of Applied Biology, 2011, 159(2): 155-168.
Role of soil microbes in the acquisition of nutrients by plants
SHEN Renfang*, ZHAO Xueqiang
StateKeyLaboratoryofSoilandSustainableAgriculture,InstituteofSoilScience,ChineseAcademyofSciences,Nanjing210008,China
Contemporary agriculture is characterized mainly by extensive fertilizer application. Worldwide, fertilizer application indeed contributes greatly to increase in crop yield, but it also causes several negative effects such as soil acidification, water eutrophication, greenhouse gas emission, and resource depletion, thereby threatening the sustainable development of agriculture. Soil microbes drive plant diversity and productivity in terrestrial ecosystems, and they are directly involved in regulating the ability of plants to acquire nutrients and the nutrient cycles occurring in soils. Therefore, it is feasible to reduce the dependence of agriculture on fertilizers through the development of soil microbial approaches. Here, we described the role of biological nitrogen fixation, mycorrhizal fungi, and plant growth-promoting rhizobacteria in enhancing the ability of plants to acquire nutrients, and the underlying mechanisms. Moreover, we investigated the effect of soil microbes on soil nitrogen and phosphorus cycles, and their relationship with soil nutrient bioavailability and nutrient loss. Based on the obtained information, soil microbial processes to improve plant nutrition have been suggested, with the objective of reducing fertilizer application. Numerous studies have demonstrated the vital role of soil microbes in improving plant nutrition, but many problems regarding the use of soil microbial approaches need to be solved. Further extensive research in this field will help us solve these problems, and accelerate the development and application of soil microbial techniques for reducing fertilizer use, thereby contributing to sustainable agriculture.
biological nitrogen fixation; mycorrhizal fungi; plant growth-promoting rhizobacteria; nutrient cycle; sustainable agriculture
中国科学院战略性先导科技专项B类(XDB15030000);国家973项目(2014CB441000)
2015-06-05;
2015-08-12
10.5846/stxb201506051140
*通讯作者Corresponding author.E-mail: rfshen@issas.ac.cn
沈仁芳,赵学强.土壤微生物在植物获得养分中的作用.生态学报,2015,35(20):6584-6591.
Shen R F, Zhao X Q.Role of soil microbes in the acquisition of nutrients by plants.Acta Ecologica Sinica,2015,35(20):6584-6591.
