固沙植被区土壤呼吸对反复干湿交替的响应
2015-01-20李小军杨昊天
赵 蓉, 李小军, 赵 洋, 杨昊天
中国科学院寒区旱区环境与工程研究所沙坡头沙漠研究试验站, 兰州 730000
固沙植被区土壤呼吸对反复干湿交替的响应
赵 蓉, 李小军*, 赵 洋, 杨昊天
中国科学院寒区旱区环境与工程研究所沙坡头沙漠研究试验站, 兰州 730000
由降水的不连续性引起的土壤干湿交替是荒漠生态系统土壤呼吸的重要影响因子。在恒温培养条件下,研究了固沙植被区土壤呼吸对不同降雨量(5、10、20 mm)条件下以10 d为降水周期的多重干湿交替过程的响应,结果表明:3个降雨量条件下的多重干湿交替过程中,土壤呼吸速率均在降雨后迅速增大,并在降雨0.5 h后达到最大值,尔后,随着土壤含水量的下降而逐渐减小并恢复到降雨前水平。随着干湿交替过程的依次进行,最大和平均土壤呼吸速率及累积碳释放量均呈现出逐渐减小的趋势。3个干湿循环周期平均呼吸速率和土壤碳释放量均随着降雨量的增加逐渐增大,土壤呼吸速率峰值表现为第1个干湿循环周期土壤呼吸速率峰值随着降雨量的增加而增大,而第2、3个周期各降雨处理下的土壤呼吸速率峰值显示出随着降雨前期土壤含水量的增加而减小的趋势。说明干湿交替过程对土壤呼吸具有显著的激发作用,且干湿交替程度(即降雨前后土壤含水量的变化)以及土壤前期所经历的干湿交替过程是影响荒漠生态系统土壤呼吸对干湿交替响应的重要因素。
土壤呼吸; 降水脉动; 干湿交替; 固沙植被区; 腾格里沙漠
占全球陆地表面1/3以上的干旱、半干旱区长期以来被认为是陆地生态系统中重要的碳源[1-2],其每年约有224 g C m-2a-1释放到大气中[3]。土壤呼吸作为该区最主要的碳释放途径,其随着温度、水分等环境因子的改变而发生的变化往往是引起大气CO2浓度波动以及土壤有机碳储量变化的重要原因[4],且有可能导致生态系统源汇功能的转变[5]。其中,与降水密切相关的土壤水分有效性是荒漠生态系统土壤呼吸的首要驱动因子[6]。研究表明,降雨所引起的土壤碳释放量能够占年土壤总碳释放量的16%—21%[7],是干旱区碳估算的重要参数。
荒漠区降水主要以不连续的偶然事件为显著特征,且存在强烈的时空变异,降水的这种特征使表层土壤常常经历干湿交替的水分环境[8]。早在1958年,Birch就发现经历长期干旱的土壤再湿润后促进了土壤CO2的释放[9]。近年来,大量研究表明,干旱土壤再湿润后会引起短暂的碳释放脉冲。荒漠区土壤长期处于干旱状况,土壤呼吸作用微弱,这种土壤再湿润所引起的碳释放脉冲可能极大地推动该区碳循环过程。然而,土壤呼吸对干湿交替响应的研究大部分都集中在农田、森林、草地生态系统以及地中海气候区[7,10-12],而对于常常经历剧烈干湿交替的温带荒漠区却少有研究;荒漠生态系统中仅有的研究也只限于单次降雨所引起的干湿交替[5],而对反复干湿交替过程所引起的土壤呼吸的动态变化鲜见报道[13]。
本研究以腾格里沙漠东南缘的天然固沙植被区土壤为研究对象,模拟了不同降雨量条件下的反复干湿交替过程,通过分析该过程中土壤呼吸的动态变化过程,揭示了天然固沙植被区土壤碳释放与降水引起的土壤干湿交替之间的关系,量化了不同降雨量条件下的多重干湿交替所引起的碳释放量,为进一步探讨干旱区土壤碳释放对全球变化背景下降水格局改变的响应提供依据。
1 材料与方法
1.1 研究区概况
研究区位于腾格里沙漠东南缘沙坡头沙漠研究试验站以西22 km的红卫天然固沙植被区(37°32′—37°26′ N,105°02′ —104°30′ E,海拔1300 m)。该区是草原化荒漠向荒漠化草原的过渡地带,年平均气温10.0℃;年平均降水量为186.2 mm,年内降水分布不均,80%左右的降水集中在7—9月;年潜在蒸发量为2300—2500 mm;植被区灌木、半灌木主要有柠条(Caraganakorshinskii)、狭叶锦鸡儿(C.stenophylla)、驼绒藜(Ceratoideslatens)、油蒿(Artemisiaordosica)、刺叶柄棘豆(Oxytropisaciphylla)等,优势草本植物有茵陈蒿(A.capillaris)、冷蒿(A.frigida)、小画眉草(Eragrostispoaeoides)、雾冰藜(Bassiadasyphylla)、细叶苦菜(Ixerisgracilias)、小车前(Plantagominuta)、叉枝鸦葱(Scorzoneradivaricata)等。主要土壤类型为灰钙土和风沙土。0—10 cm土壤理化性质见表1。

表1 土壤理化性质
1.2 样品采集
2013年6月,距离上一次降水事件30d时(降低上一次降雨对实验的影响),在实验区灌丛间地势平坦的开阔地按照随机原则用内径10.4 cm、高10 cm的PVC管采集9个原状土。为了保证样品的完整性,采样前,用喷壶在土壤表面均匀喷洒少量蒸馏水,待水分渗入土壤表层后,开始取样。将样品带回实验室放在阴凉通风处备用。
1.3 室内培养与土壤呼吸测定
将样品置于智能生化培养箱(SHP-250)中,在25℃条件下进行恒温暗培养,预培养3 d(使土壤达到稳定状态)后,开始试验。根据沙坡头站多年平均降水数据显示,雨季(7—9月)单次降水量以< 10 mm 的降水出现频次最多,降水周期约为6—15 d,因此本试验设置了3个降水梯度(5、10 mm和20 mm),每个降水梯度下设置3个以10 d为降水周期的干湿循环周期,每个处理设3个重复。模拟降水采用喷壶喷洒的方式,向土壤表面均匀喷洒蒸馏水,加水当天记作第0天,第10天、20天分别采用相同方法向土壤表面喷洒蒸馏水。第0天加水后至第10天加水前记作第1个循环周期,第10天加水后至第20天加水前记作第2个循环周期,第20天加水后至第30天记作第3个循环周期。
土壤呼吸速率采用Li-6400-09土壤呼吸室(Li-COR, INC, US)进行测定。分别于加水后0.5、2、5、12、24 h进行测定,之后每隔24 h测定1次,直到下一次加水后进行新一轮测定。同时采用1/100电子天平称量样品重量,实验结束后用烘干法(105℃条件下烘干24 h)测定并计算土壤质量含水量。
1.4 数据处理
采用时间加权的方法,计算实验期间土壤累积碳释放量及每一个干湿交替周期土壤平均呼吸速率和累积碳释放量。采用重复测定方差分析(Repeated Measure of ANOVA)和Duncan检验进行方差分析;采用双因素方差分析(Two-way ANOVAs)检验干湿交替次数和降雨量对最大呼吸速率、平均呼吸速率、总碳释放量的影响。所有统计分析均采用SPSS 16.0(Chicago USA),绘图采用Origin 8.5。
2 结果与分析
2.1 干湿交替对土壤呼吸速率的影响
土壤干湿交替显著地激发了土壤呼吸(P<0.05, 表2),各降雨量条件下,土壤呼吸速率均在降雨后0.5 h达到最大值,随后逐渐下降(图1)。3种降雨量条件下的土壤呼吸速率峰值均随着干湿交替次数的增加而逐渐减小,且随着降雨量的增加,土壤呼吸速率峰值减小的比例增大。不同降雨量处理之间,土壤呼吸速率峰值呈现出不同规律。其中,第1个周期土壤呼吸速率峰值随降雨量的增加而依次增大(P<0.05),而第2、3个周期土壤呼吸速率峰值却呈现出随着降雨前土壤含水量的增加而减小的趋势。干湿交替次数及其与降雨量的交互作用对土壤呼吸速率峰值影响显著(P<0.05, 表3)。
第1个循环周期,3种降雨量条件下土壤呼吸速率峰值分别为5.65、6.74、7.50 μmol m-2s-1,是模拟降雨前(0.13 μmol m-2s-1)的43—58倍,随后土壤呼吸速率逐渐下降,并分别在48、96 h以及192 h内恢复到降雨前水平;第2个循环周期土壤呼吸速率峰值分别为4.17、3.81、3.44 μmol m-2s-1,较第1个循环周期分别下降了26%、43%和54%,是第1次模拟降雨前的26—32倍,分别在24、72 h和168 h恢复到降雨前水平;第3个循环周期土壤呼吸速率峰值分别为3.17、3.56、2.92 μmol m-2s-1,较第1个循环周期分别降低了44%、47%和61%,是第1次模拟降雨前的22—27倍,分别在48、48 h和144 h恢复到降雨前水平。

表2 降雨量和干湿交替次数对总体土壤呼吸速率影响的重复测定方差分析
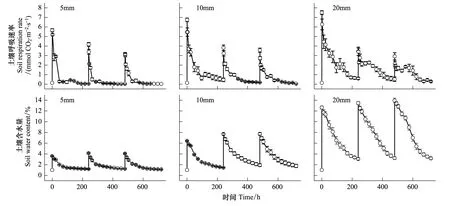
图1 干湿交替条件下土壤呼吸速率和土壤含水量随时间的变化特征Fig.1 Dynamics of soil respiration rates and soil moisture under repeated drying and rewetting cycles
降雨量与干湿交替次数均显著影响平均土壤呼吸速率(表3,P<0.05)。各降雨量条件下的干湿循环周期平均土壤呼吸速率均显著高于降雨前(P<0.05)。第1个循环周期各降雨量条件下平均土壤呼吸速率分别为0.67、1.37、2.14 μmol m-2s-1,是降雨前土壤呼吸速率的5—16倍;第2个循环周期各降雨量条件下平均土壤呼吸速率分别为0.29、0.69、1.52 μmol m-2s-1,较第1个循环周期下降了57%、50%、29%;第3个循环周期各降雨量条件下平均土壤呼吸速率分别为0.34、0.58、1.11 μmol m-2s-1,较第1个循环周期下降了49%、58%、48%。,3种降雨量条件下土壤平均呼吸速率均随着干湿交替次数的增加呈现出逐渐下降的趋势(第2、3个循环周期差异不显著)。随着降雨量的增加,3个循环周期平均土壤呼吸速率均逐渐增大(图2)。其中,第1个循环周期5 mm和20 mm差异显著(P<0.05),而与10 mm之间差异均不显著;第2、3个循环周期5 mm和10 mm差异不显著,而与20 mm之间差异均显著(P<0.05)。
2.2 干湿交替对土壤累积碳释放量的影响
干湿交替次数和降雨量均显著影响土壤累积碳释放量(P<0.05, 表3)。3个干湿循环土壤累积碳释放量随时间的变化均表现出先快后慢的特征,且随着降雨量的增加,土壤累积碳释放量逐渐增大(图3)。第1个循环周期各降雨量条件下,土壤累积碳释放量分别为6.97、14.24、22.20 g C/m2;第2个循环周期各降雨量条件下,土壤累积碳释放量分别为2.97、7.20和15.76 gC/m2,较第1个循环周期减少了57%、49%和29%;第3个循环周期各降雨量条件下,土壤累积碳释放量分别为3.57、5.99、11.53 g C/m2,较第1个循环周期减少了49%、58%和48%。3个循环土壤碳释放量均随着降雨量的增加逐渐增大。其中,第1个循环周期5 mm和20 mm差异显著(P<0.05),而与10 mm之间差异均不显著;第2、3次循环周期5 mm和10 mm差异不显著,而与20 mm之间差异均显著(P<0.05)。相同降雨量条件下,随着干湿交替次数的增加,土壤累积碳释放量呈下降趋势,但第2、3次干湿循环之间差异不显著(图4)。
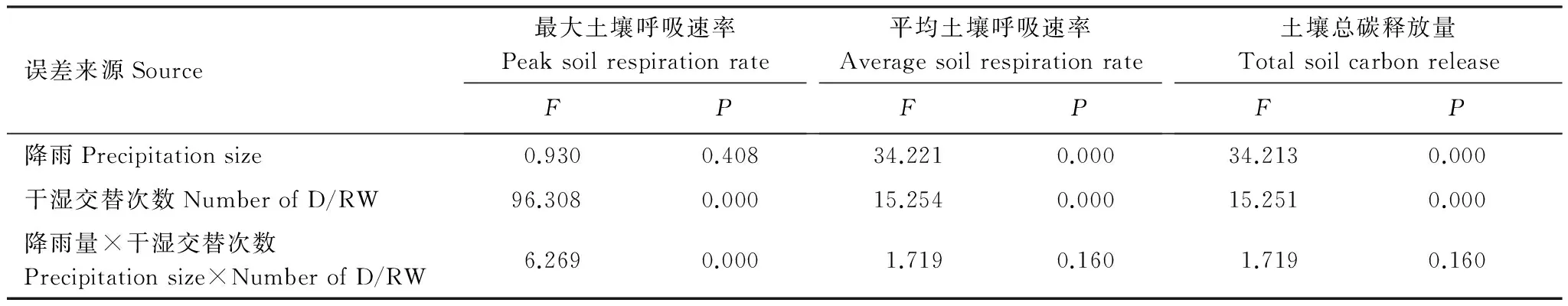
表3 降雨量和干湿交替次数对土壤呼吸速率峰值、平均值和总碳释放量的影响
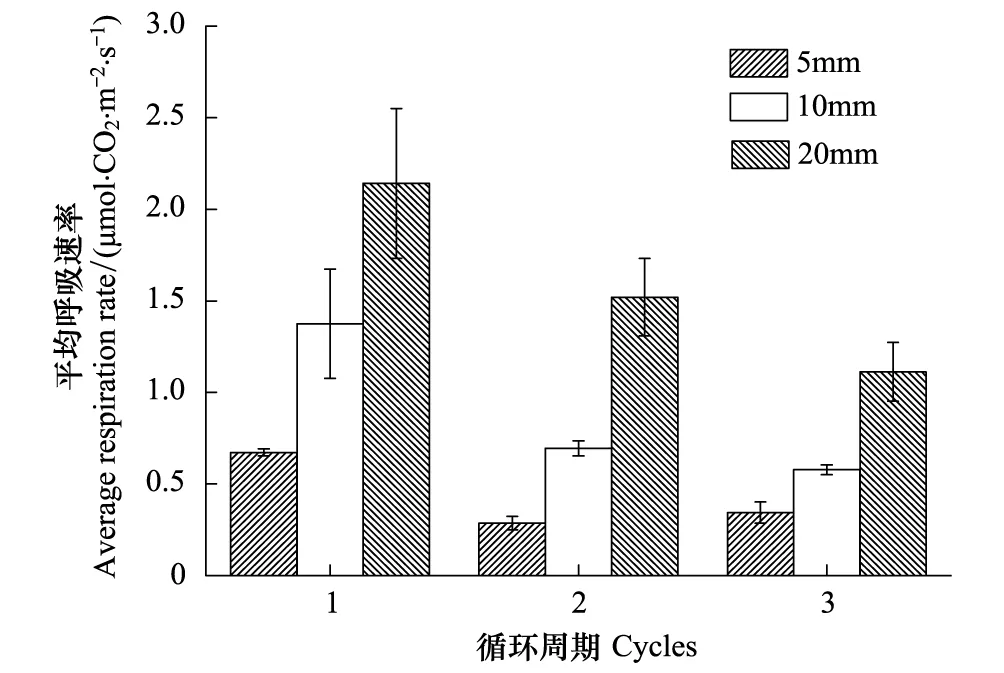
图2 不同降雨量条件下干湿循环过程平均土壤呼吸速率Fig.2 Average soil respiration rates of different drying and rewetting cycles
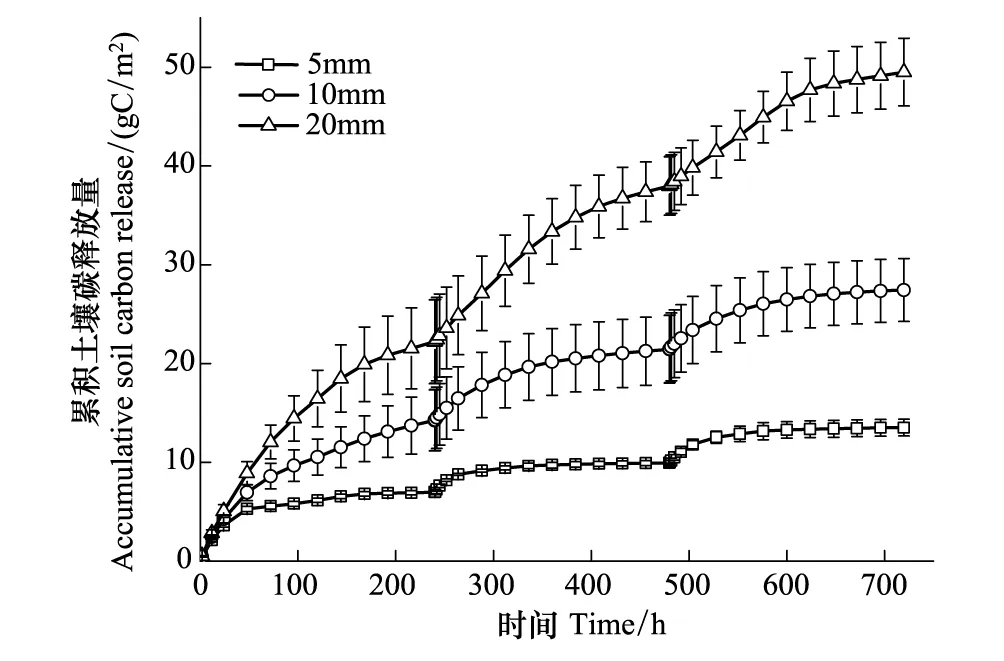
图3 干湿交替条件下累积土壤碳释放量随时间的变化特征 Fig.3 Dynamics of accumulated soil carbon release under drying and rewetting cycles
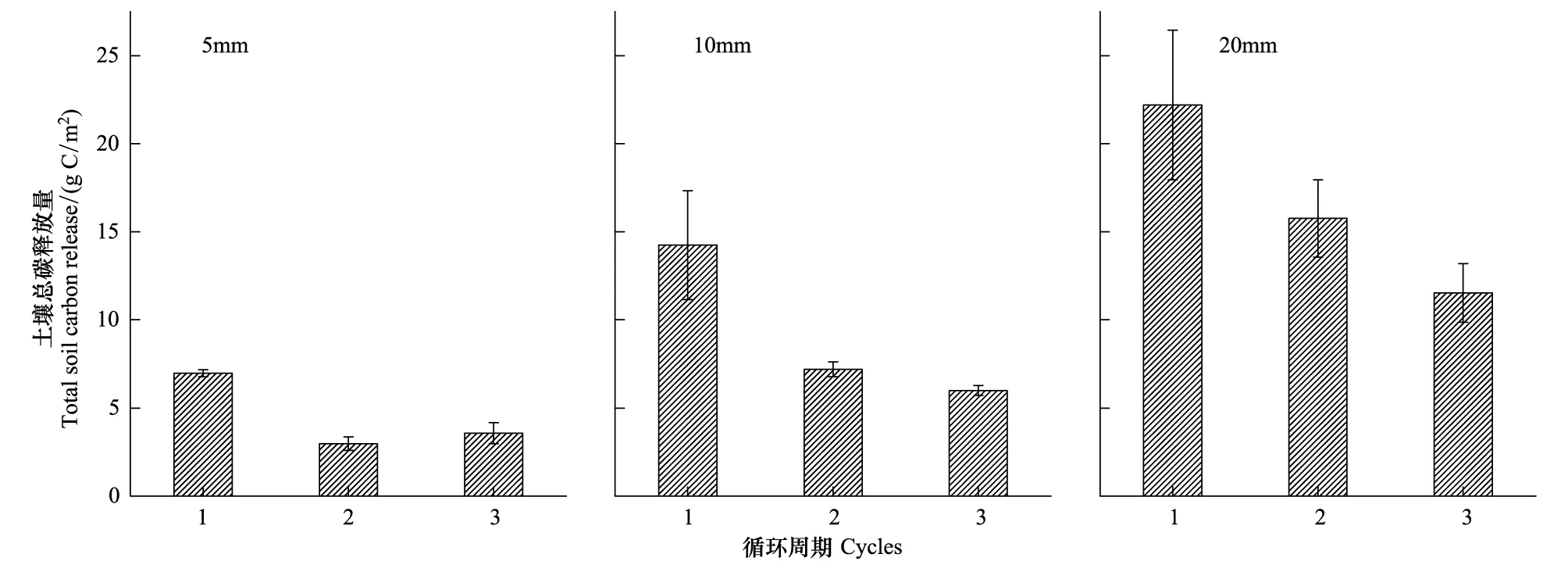
图4 不同降雨量条件下3个干湿循环过程土壤总碳释放量Fig.4 Total carbon release of three drying and rewetting cycles with different precipitation
2.3 土壤呼吸速率与土壤含水量的关系
用对数函数分别拟合3个循环周期土壤呼吸速率与土壤含水量的关系,拟合方程分别为:y=-6.22+4.18ln(x+3.37)(R2=0.61,P<0.01),y=-2.49+2.00ln(x+2.05) (R2=0.62,P<0.01)和y=-0.81+1.13ln(x+0.78) (R2=0.54,P<0.01)。3个循环周期土壤呼吸速率均随着土壤含水量的增加而增大,而随着干湿循环次数的增加,这种增大的趋势逐渐减缓(图5)。
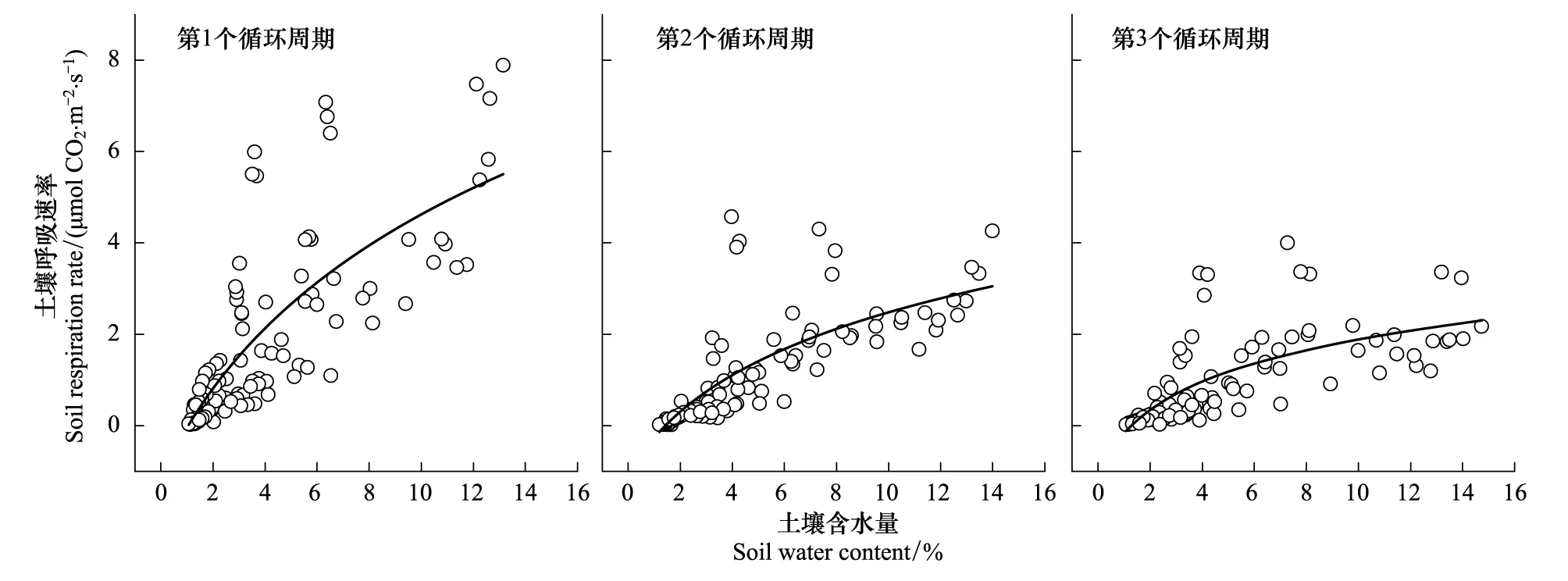
图5 土壤呼吸速率与土壤含水量的关系Fig.5 The relationship between soil respiration rate and soil moisture of three drying and rewetting cycles
3 讨论
降水的不连续性导致荒漠生态系统中土壤常常经历干湿交替的水分环境,从而影响一系列地下生态学过程,并进一步影响土壤呼吸动态。本实验结果表明,降水引起的土壤干湿交替显著促进了土壤呼吸,使土壤呼吸速率在短时间内升高数倍,这与Sponseller在亚热带荒漠生态系统所做的研究结果一致[5]。目前认为干湿交替对土壤呼吸的激发主要是由于土壤再湿润后微生物可利用的有机基质增加,其来源主要有两方面,即经历了干湿交替的土壤颗粒所释放的前期受到保护的有机质[9],以及水势的迅速变化导致微生物释放的渗透调节溶质或微生物死亡后细胞溶解释放的微生物量[14]。干湿交替过程使土壤中可供微生物利用的有机质迅速增加,土壤微生物活性增强[15],这是土壤呼吸速率以及碳释放量随干湿交替过程变化的根本原因。随着降雨量的增加,干湿交替所引起的土壤碳释放量逐渐增大。这是由于小降雨事件仅能湿润土壤表层,且水分迅速蒸发,降雨无法到达下层土壤而激活微生物。降雨量的增加使更多的水分渗透到下层土壤,一方面,水分的入渗排出了土壤孔隙中在干旱期积累的CO2,另一方面,土壤中水分的增多导致水势的变化更加剧烈,微生物需要释放更多的渗透调节溶质以达到与土壤水势的平衡,而一些渗透调节能力较弱的微生物在渗透压冲击下导致细胞破裂,释放出体内有机质。水分和有机质有效性的增加激发了微生物活性,导致土壤释放量进一步增加。
土壤呼吸对干湿交替的响应不仅受到降雨量的影响,还与降雨前期土壤干旱程度密切相关。本研究结果显示,第2、3个循环周期各降雨处理下的土壤呼吸速率峰值并未随着降雨量的增加而增大,这可能是降雨前期土壤含水量的差异所导致。Cable等人在Sonoran沙漠的研究表明,土壤处于干旱状况时,降雨对土壤呼吸速率的激发作用达到864%,而降水前期土壤处于湿润的水分环境时,降雨后土壤呼吸速率仅增加了25%[16]。这可能是由于前期土壤含水量较低的土壤再湿润后,其微生物量的周转及胞内溶质的矿化更加强烈[17]。此外,较干旱的土壤孔隙较大,降水入渗后排出的CO2也为碳释放脉冲做出部分贡献。本实验中第2、3个干湿循环周期土壤呼吸速率峰值并未随降雨量的升高而升高,可能是由降雨量较高的土壤其再次降雨前含水量相对于降雨量较低的土壤更高,其呼吸速率峰值受到限制。因此,干湿交替的程度,即降雨前后土壤含水量差异的大小是影响土壤呼吸对干湿交替响应的一个重要原因。
干湿交替显著促进了土壤呼吸速率,并提高了土壤累积碳释放量,但随着干湿交替次数的增加,这种促进作用逐渐减弱,这与Muhr等人在森林生态系统的研究结果一致[18]。一方面,这可能是由于经过重复的干湿交替过程,土壤中的可利用有机质减少[19],或由于土壤团聚体稳定性的提高,从中释放的有机质减少[20]。另一方面,重复的干湿交替导致土壤微生物生物量、群落结构以及生理活性的改变也是引起CO2释放脉冲减小的重要原因。Butterly在农田生态系统中的研究结果表明,在第1次干湿交替过程中土壤微生物量减少了50%—75%,但幸存的微生物与CO2释放的减少量不成比例,且随着干湿交替的进行,土壤中可溶解的有机物减少,因此认为碳释放脉冲的减小可能是土壤微生物量和可利用有机物的减少共同引起的[10]。Franzluebbers则认为,干湿交替改变了土壤微生物群落的组成[21],这可能也是引起CO2释放脉冲减小的原因之一。经过干湿交替后重新建立的微生物群落,其活性降低,生理状况发生改变,可能降低了它们对水势变化的敏感性[22],因而减少了由水势变化所引起的渗透调节物质的释放以及渗透压冲击导致的微生物细胞溶解。
荒漠区土壤长期处于干旱状况,土壤呼吸作用微弱,干湿交替所引起的土壤碳释放激增效应改变了这种状况,加速了土壤有机碳的释放,使土壤碳释放出现短暂的脉冲现象。然而随着干湿交替的反复发生,这种脉冲逐渐减小,这可能意味着反复的干湿交替将会引起土壤有机碳库的减小,并进一步加剧受水分和养分双重限制的干旱区土壤养分的贫瘠化。全球变化所引起的降水格局的改变,即干旱和强降水事件的增多[23],可能进一步增强土壤干湿交替过程,这意味着降水前后土壤含水量变化更加强烈。根据本实验研究结果推测,这将使土壤碳释放的激增效应更加强烈,这是否会进一步影响温带荒漠土壤碳释放动态及土壤有机碳储量,还有待于做进一步研究,并扩展到更大的时空尺度。然而,考虑到野外条件下通过降尘以及植被凋落物等方式向土壤输入新碳,由于室内实验的局限性,土壤缺少新碳的输入,因此野外条件下土壤呼吸对干湿交替的响应程度可能会有所不同,还需要进一步做野外实验以达到更高的精确度。
[1] Reynolds J F, Smith D M S, Lambin E F, Turner II B L, Mortimore, M, Batterbury S P J, Downing T E, Dowlatabadi H, Fernández R J, Herrick J E, Huber-Sannwald E, Jiang H, Leemans R, Lynam T, Maestre F T, Ayarza M, Walker B. Global desertification:building a science for dryland development. Science, 2007, 316(5826):847-851.
[2] Liu X Z, Wan S Q, Su B, Hui D F, Luo Y Q. Response of soil CO2efflux to water manipulation in a tallgrass prairie ecosystem. Plant and Soil, 2002, 240(2):213-223.
[3] Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B, 1992, 44(2):81-99.
[4] Raich J W, Potter C S, Bhagawati D. Interannual variability in global soil respiration. Global Change Biology, 2002, 8(8):800-812.
[5] Sponseller R A. Precipitation pulses and soil CO2flux in a Sonoran Desert ecosystem. Global Change Biology, 2007, 13(2):426-436.
[6] Huxman T E, Snyder K A, Tissue D, Leffler A J, Ogle K, Pockman W T, Sandquist D R, Potts D L, Schwinning S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia, 2004, 141(2):254-268.
[7] Lee X H, Wu H J, Sigler J, Oishi C, Siccama T. Rapid and transient response of soil respiration to rain. Global Change Biology, 2004, 10(6):1017-1026.
[8] Noy-Meir I. Desert ecosystems:environment and producers. Annual Review of Ecology and Systematics, 1973, 4(1):25-51.
[9] Birch H F. The effect of soil drying on humus decomposition and nitrogen availability. Plant and Soil, 1958, 10(1):9-31.
[10] Butterly C R. Drying/Rewetting Cycles in Southern Australian Agricultural Soils:Effects on Turnover of Soil Phosphorus, Carbon and the Microbial Biomass [D]. Australia:The University of Adelaide, 2008.
[11] Xiang S R, Doyle A, Holden P A, Schimel J P. Drying and rewetting effects on C and N mineralization and microbial activity in surface and subsurface California grassland soils. Soil Biology and Biochemistry, 2008, 40(9):2281-2289.
[12] Unger S, Máguas C, Pereira J S, David T S, Werner C. Interpreting post-drought rewetting effects on soil and ecosystem carbon dynamics in a Mediterranean oak savannah. Agricultural and Forest Meteorology, 2012, 154-155:9-18.
[13] 欧阳扬, 李叙勇. 干湿交替频率对不同土壤CO2和N2O释放的影响. 生态学报, 2013, 33(4):1251-1259.
[14] Harris R F. Effect of water potential on microbial growth and activity. Water potential relations in soil microbiology, 1981 (waterpotentialr), 23-95.
[15] Orchard V A, Cook F J. Relationship between soil respiration and soil moisture. Soil Biology and Biochemistry, 1983, 15(4):447-453.
[16] Cable J M, Ogle K, Williams D G, Weltzin J F, Huxman T E. Soil texture drives responses of soil respiration to precipitation pulses in the Sonoran Desert:implications for climate change. Ecosystems, 2008, 11(6):961-979.
[17] Fierer N, Schimel J P. Effects of drying-rewetting frequency on soil carbon and nitrogen transformations. Soil Biology and Biochemistry, 2002, 34(6):777-787.
[18] Muhr J, Goldberg S D, Borken W, Gebauer G. Repeated drying-rewetting cycles and their effects on the emission of CO2, N2O, NO, and CH4in a forest soil. Journal of Plant Nutrition and Soil Science, 2008, 171(5):719-728.
[19] Chow A T, Tanji K K, Gao S D, Dahlgren R A. Temperature, water content and wet-dry cycle effects on DOC production and carbon mineralization in agricultural peat soils. Soil Biology and Biochemistry, 2006, 38(3):477-488.
[20] Denef K, Six J, Bossuyt H, Frey S D, Elliott E T, Merckx R, Paustian K. Influence of dry-wet cycles on the interrelationship between aggregate, particulate organic matter, and microbial community dynamics. Soil Biology and Biochemistry, 2001, 33(12-13):1599-1611.
[21] Franzluebbers K, Weaver R W, Juo A S R, Franzluebbers A J. Carbon and nitrogen mineralization from cowpea plants part decomposing in moist and in repeatedly dried and wetted soil. Soil Biology and Biochemistry, 1994, 26(10):1379-1387.
[22] Mikha M M, Rice C W, Milliken G A. Carbon and nitrogen mineralization as affected by drying and wetting cycles. Soil Biology and Biochemistry, 2005, 37(2):339-347.
[23] IPCC. IPCC Fourth Assessment Report-Climate Change:the Physical Science Basis. Cambridge:Cambridge University Press, 2007.
Response of soil respiration to repeated cycles of drying and rewetting in soils of the sand-fixed region of the Tengger Desert
ZHAO Rong, LI Xiaojun*, ZHAO Yang, YANG Haotian
ShapotouDesertResearchandExperimentStation,ColdandAridRegionsEnvironmentalandEngineeringResearchInstitute,ChineseAcademyofSciences,Lanzhou730000,China
In arid and semi-arid regions, the climate is characterized by long dry periods interspersed by occasional precipitation, owing to which the soil surface undergoes frequent cycles of drying and rewetting (D/RW). This influences underground carbon (C) mineralization and produces a transient CO2pulse after each rewetting. Although the rewetting event is short-lived, the CO2production that it triggers may constitute a significant portion of the carbon released annually because little carbon is released in dry periods. The aim of this paper is to understand the response of CO2efflux to repeated D/RW cycles and to elucidate the mechanisms that regulate carbon cycling in dry lands. As a case study, we examined the soil from a sand-fixed vegetation area in the southeastern fringe of the temperate Tengger Desert of northern China. We subjected soil samples to laboratory incubation at a constant temperature of 25°C, with simulated rainfalls occurring at 10-day intervals. Three treatment conditions differed in terms of precipitation volume (5, 10, or 20 mm). Soil respiration rate and soil water content were measured 1 h before rewetting (baseline); 0.5, 2, 12, and 24 h after rewetting; and every 24 h until the next cycle. We analyzed the changes in soil CO2efflux and moisture after each cycle by comparing peak and average respiration rates and total CO2production. The results showed that all cycles stimulated soil respiration, which peaked at 0.5 h after rainfall and then gradually decreased to baseline level. Respiration rates and carbon release both increased with increasing precipitation. As the D/RW cycles proceeded, the peak and average soil respiration rates and amounts of carbon released tended to decrease. Although peak respiration rates following the first rewetting increased with precipitation volume, peak respiration rates of the second and third rewetting of soils with different precipitation amounts did not vary with precipitation volume but did correlate with antecedent moisture.Our results indicate that rewetting of dry soil significantly accelerates soil CO2efflux, but the magnitude of this effect gradually decreases as D/RW cycles are repeated. Furthermore, the intensity of drying and rewetting and the soil water present before precipitation both influence the soil respiration response in temperate desert soil. This implies that the soil respiration response depends not only on rainfall but also on antecedent soil conditions. Climate models predict that precipitation patterns will likely intensify in mid-latitude regions, with increases in total precipitation, drought period duration, and frequency of extreme precipitation events. The water environment of these soils will become more severe, which will intensify the effects of pulsed precipitation events and further impact soil C storage and fertility due to the relatively low levels of organic C content.
soil respiration; precipitation pulse; drying and rewetting cycles; sand-fixing vegetation area; Tengger Desert
国家自然科学基金项目(41171078)
2014-04-09; < class="emphasis_bold">网络出版日期:
日期:2014-12-18
10.5846/stxb201404090690
*通讯作者Corresponding author.E-mail: xiaojunli@lzb.ac.cn
赵蓉, 李小军, 赵洋, 杨昊天.固沙植被区土壤呼吸对反复干湿交替的响应.生态学报,2015,35(20):6720-6727.
Zhao R, Li X J, Zhao Y, Yang H T.Response of soil respiration to repeated cycles of drying and rewetting in soils of the sand-fixed region of the Tengger Desert.Acta Ecologica Sinica,2015,35(20):6720-6727.
