土壤微生物群落构建理论与时空演变特征
2015-01-20贺纪正王军涛
贺纪正,王军涛
中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085
土壤微生物群落构建理论与时空演变特征
贺纪正*,王军涛
中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085
土壤微生物作为陆地生态系统的重要组成部分,直接或间接地参与几乎所有的土壤生态过程,在物质循环、能量转换以及污染物降解等过程中都发挥着重要作用。对土壤微生物时空演变规律及其形成机制的研究,不仅是微生物演变和进化的基础科学问题,也是预测微生物及其所介导的生态功能对环境条件变化响应、适应和反馈的理论依据。讨论了土壤微生物群落的定义、测度方法和指标,认为群落是联系动植物宏观生态学与微生物生态学的基础,群落构建机制是宏观和微观生态学都需要研究的核心科学问题;从生态学的群落构建理论出发,阐述了包括生态位理论/中性理论、过程理论和多样性-稳定性理论在土壤微生物时空演变研究中的应用,以及微生物群落在时间和空间上的分布特征及其尺度效应;确立了以微生物群落构建理论为基础、不同时空尺度下土壤微生物群落演变特征为主要内容的微生物演变研究的基本框架。
土壤微生物;微生物地理学;微生物演变;生物多样性;群落构建
土壤微生物作为陆地生态系统的重要组成部分,直接或间接地参与几乎所有的土壤生态过程,在物质循环、能量转换以及污染物降解等过程中都发挥着重要作用[1]。对土壤微生物时空演变规律及形成机制的理解是全面认识其在生态系统所执行功能的必要条件[2]。与动植物相比,由于微生物体积微小、数量庞大、代谢速率快和变异性强,一度被认为是不存在地理分布格局的。近年来,微生物领域研究的进展揭示了动植物具有的部分时空演变规律,比如种-面积关系(Taxa-area Relationship),距离-衰减关系(Distance-decay Relationship)以及广域种-稀有种的分化等在微生物中都存在[3-5]。这些结果同时也表明,尽管微生物具有体积小、数量多这些特性,但其与其他生物的分布规律仍然具有共性。因此,建立对于动植物和微生物都适用的理论架构[6]、在有效的理论框架指导下探索微生物群落的时空演变特征及形成机制并揭示生物时空演变的一般规律是生态学未来发展的必然方向。
个体是生命特征的最小表现单元,但是在人类现有的认知和研究能力范围内,自然状态下土壤中的微生物从来不是以个体或种群形态单独存在的,群体或者群落才是其存在的基本单元[7]。在这样的单元中,微生物之间以及微生物与土壤环境之间进行着物质能量流动和信息交换,不同的微生物密切联系作为一个整体共同行使群落的功能。对于微生物时空分布的研究实际上是探索不同时间和空间条件下微生物群落的变化规律,即群落是微生物地理学研究的基本单位。而在宏观生态学理论中,群落的构建机制是生物多样性产生和维持的核心研究内容[8]。因此,对于群落的共同关注也是动植物生态学和微生物生态学理论寻求统一的重要基础。
长久以来,由于缺乏理论的指引,微生物生态学的研究大多只能是就事论事、分析表象,而不具有预见性。微生物时空演变的理论也一度仅仅以荷兰微生物学家Beijerinck提出的“环境条件决定微生物”为指导[9]。事实上,土壤微生物时空演变的研究是由生态学到微生物生态学领域的延伸。微生物生态学的理论工具并非普通微生物学研究的思维定式,其理论发展的定位应是以生态学的理论与指标体系为基础,以地理学和生物学等相关学科理论为支撑。本文讨论了土壤微生物群落的定义、测度方法和指标,并从生态学中群落构建的角度出发,阐述了包括生态位/中性理论、过程理论和多样性-稳定性理论在内的群落构建的理论,以及微生物群落在时间和空间上的分布特征及其尺度效应;指出土壤微生物生态学研究最终要能够在时空尺度上对微生物的多样性和功能进行预测,以及实现这一目标还需要多学科努力。
1 土壤微生物群落的定义及测度
生态学中对于群落的定义为“特定时空条件下,生活于具有明显表观特征的生境下、相互关联的不同类群生物的有序集合体”,其基本特征包括外貌、种类组成与结构(如捕食关系等)、群落环境、分布范围和边界特征等[10]。虽然这一定义中实际上包含了一定范围内的所有动物、植物和微生物,但事实上不同生态学家说到群落的时候,往往指的只是自己研究的对象,即动物群落或者植物群落。这种说法上的差异在动物和植物研究中基本不存在问题,但是对于土壤微生物,以下问题需要注意(表1):(1)样本的代表性决定了群落。微生物绝大多数是肉眼不可见的,因此对于土壤微生物群落来说,外在形貌、边界特征、分布范围这些都是比较模糊的概念。实际的研究中是以极少量土壤样本(0.25克到几克)中的群落代表目标范围内的微生物群落。比如不同土壤类型之间微生物群落差异是很大的,但同一类型的土壤在养分梯度或者不同土层上也存在差异[11]。因此,土壤微生物群落的界定是以研究目的为导向,而样本的代表性决定了土壤微生物群落之间的差异。(2)对微生物群落的表征取决于研究的问题。对微生物群落的表征可以面向包括真核生物和原核生物的所有微生物,也可以只针对执行特定功能的某些类群,即群落可以分为发生型和功能型两类。(3)对于群落内部的不同微生物之间的作用关系(即群落结构)目前并不能给出非常明确而全面的证据,因此我们只能先验性地认为微生物群落内各类群之间存在联系,而后在此基础上进行验证。微生物之间的作用关系,可以通过分析代谢途径反映或者通过生物统计方法(比如网络分析[12]等)计算不同分类单元在数量上的变化规律从而揭示两者可能存在的联系。
对微生物群落的物种组成进行测度的基础在于对微生物“种”的界定。由于微生物不具备明显的生殖隔离等特征,因此分子生物学中以序列的相似度定义微生物物种。最经典的判定方法为DNA序列杂交率70%以上则为同一物种,但该方法在现在的研究中并未得到广泛应用。实际操作中,通常以基于rRNA基因或者编码特定酶的功能基因的可操作分类单元(Operational Taxonomic Unit, OTU)表征物种水平的分类单元。一般认为相似度大于97%的rRNA基因[13]序列属于同一物种(即同一OTU);对于功能基因,这一数据会视具体对象而有所变化,比如目前一般认为氨单加氧酶amoA基因的为85%,而固氮酶nifH基因的则为90%。同样,微生物之间的亲缘关系也是以基因序列的相似度来度量。当然,这种基于人为设定DNA序列相似度的界定方式在一定程度上并不能反映真实的物种差异[2]。相比之下对土壤宏基因组DNA进行全基因组测序并拼接的方法能更准确揭示物种组成[14],但由于土壤微生物种类庞大,全基因组拼接的准确度和难度都很高,因此并不常用。总体来说,基于OTU的物种分类方法虽然不是最完美的但却是当前最有效的微生物物种测度指标。
在确立微生物物种的基础上,可进一步对多样性和多度指标进行考量,比如测度群落内部的α多样性(丰富度以单个样本中的OTU数量表征,均匀度以Simpson均匀度指数[15]或者Gini系数表征[16],多样性以Shannon指数或者基于亲缘关系的Faith指数[17]等表征),反映群落之间差异的β多样性(又称周转率,包括Sørensen距离和Jaccard距离、Bray-Curtis距离以及基于微生物亲缘关系的UniFrac距离[18]等),以及γ多样性(即洲际尺度上的α多样性)。多度有两种表征方法:一种是通过对特定基因进行定量PCR分析后折算为每克土壤中该基因的拷贝数表征;另一种则是通过测序方法,以不同分类水平上的序列条数占总序列条数的百分比表征(即多度格局[19])。
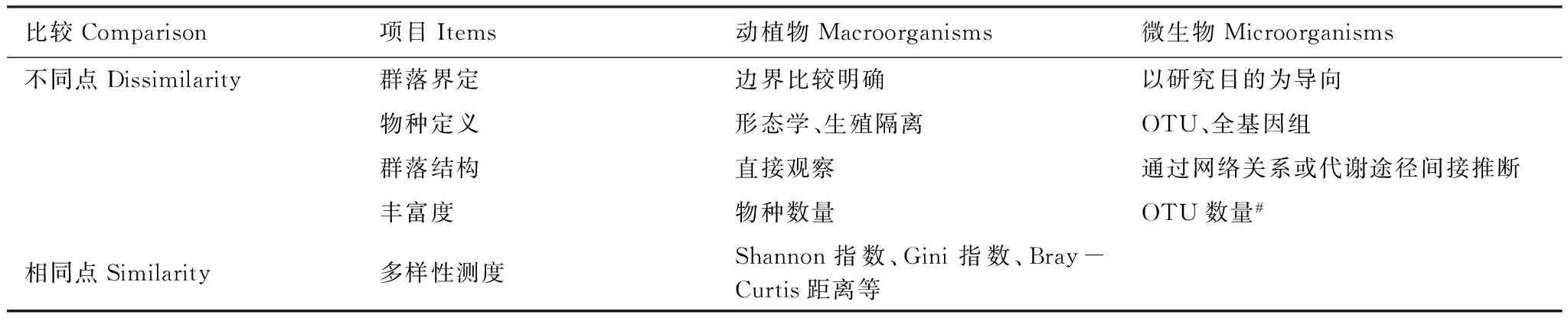
表1 动植物群落与微生物群落及测度方法的异同
2 土壤微生物群落构建的理论基础
理论的完善是学科发展的充要条件。微生物生态学理论的发展包含了两方面的内容:一方面是在现有的生态学研究体系下发展适用于微生物的理论框架;另一方面则是微生物领域的进展对原有生态学理论的补充和反馈。群落的构建机制是宏观生态学体系的核心议题[8],同时群落也是微生物生态学研究的基本对象[7],因此土壤微生物时空演变机制实质上也是在不同空间和时间尺度下微生物的群落构建及演变机制。微生物群落构建理论基础主要有三种,即:生态位理论/中性理论和过程理论,多样性-稳定性理论。
2.1 生态位理论/中性理论
生态位理论与中性理论:生态位理论体现的是群落中不同生物对于各种环境条件和压力适应能力的综合表现相互区别[8],其生理基础在于生物对于环境条件的耐受范围(即生态幅);中性理论忽略环境选择作用但强调中性突变对于群落内生物多样性形成的重要性[7],其生理依据是中性突变学说,即分子水平上无害的随机变异与漂变等过程产生生物多样性。前者强调的是确定性过程的作用,后者强调的是随机过程的影响。因此,尽管生态位理论和中性理论并非同时产生,但往往被放在一起进行比较,是矛盾对立但又互补统一的两个方面。
生态位理论和中性理论发展时间相对较长,因此,与之相关的理论体系最为丰富。比如微生物地理学中最著名的Bass-Becking假说“Everything is everywhere, but the environment selects”[9]强调环境条件对于微生物的选择作用,事实上就是生态位理论的体现。而中性过程可以被认为是匀质环境下微生物群落产生种-面积关系或者距离-衰减关系的基础[4]。以生态位理论/中性理论为基础,还进一步衍生出了生态-历史理论框架[20]:以小的时空尺度(生态位过程起主要作用)定义“生境(Habitat)”、以大的时空尺度(中性过程起主要作用)定义“域(Province)”。生态-历史理论框架通过将微生物的地理格局归因于生境与域的组合框架,即单生境单域(零假设,即微生物无地理分布格局)、单生境复域、复生境单域和复生境复域,从而将Bass-Becking假设也纳入了该框架之下(即复生境单域)。多个研究中都利用这一框架对于微生物的地理格局成因做出解释,比如在对三个隔离的站点(域)中不同施肥处理(生境)对土壤微生物群落多样性影响的研究中,发现历史因素的作用大于环境因子的作用[21]。另一项针对水体微生物的研究则发现,趋磁细菌的分布格局与盐度呈显著相关而与空间距离无显著关系[22],即环境因子而非历史因素主导微生物的地理格局。历史-生态理论框架的广泛应用也表明了经典的生态位理论/中性理论在微生物生态研究中仍然充满活力。
2.2 过程理论
生态位理论/中性理论从经典生物地理学的角度解释微生物的地理分布格局。这种由现象到理论机制的研究顺序贯穿了生物地理学发展的历史。虽然这是探究自然界的普遍模式,然而在解释生物多样性地理格局的时候,却造成了一种比较混乱的局面:对于同一种格局,可以存在不同的理论解释;格局的普遍性不断被确认,而理论解释却无法在同一种格局中通用,即理论的应用缺乏普适性[23]。针对这一问题,2010年,Vellend根据种群遗传学的选择、漂变、突变和基因流动(即扩散)这四种在基因水平上的演化动力的概念,提出了一个基于过程的群落生态学理论框架[24]。他指出,群落的构建是由选择、漂变、物种形成和扩散这四个基本过程所决定的,所有的群落模式都可以用这四种过程不同程度作用的组合来解释[23]。Hanson等人[25]在原来的历史-生态论[20]的基础上融合了该理论框架,提出生态和进化尺度上的选择(Selection)、漂变(Drift)、变异(Mutation)和扩散(Dispersal)这四个过程是微生物地理格局产生与维持的原因,现阶段解释微生物地理格局的各种理论事实上都是对这四个过程不同程度上的强调。
过程理论认为生态过程和中性过程的边界并不清晰,两者也不应该被完全隔离,选择、漂变、变异和扩散这四个过程在生态和进化水平上都发挥作用。变异在基因、基因型和个体水平上发生,是生物多样性产生的基础;选择决定了对环境适应性更强的变异被保留,也在群落内改变不同物种的相对多度并达到平衡状态;漂变的结果使得某一特定变异消亡,或者通过建立者效应而形成单一的优势种群;扩散通过减少种群群落之间的差异而抵消格局异质性的产生。基于这一框架,室内与室外空气微生物群落的差异被认为是由于局域尺度上的扩散限制导致的,而非室内环境条件的选择性[26];而区域尺度上对于土壤氨氧化微生物的研究强调了土壤pH作为环境选择条件对于其地理格局的驱动作用[27];针对淡水、沉积物、土壤等不同生境之间的微生物群落差异的研究也强调了选择过程对于微生物群落构建的重要性[28]。
2.3 多样性-稳定性理论
与过程理论拆分具体过程不同,群落构建的多样性-稳定性理论关注的是微生物群落整体所处的状态。群落的多样性-稳定性理论最早在20世纪50年代由MacArthur等人提出,指的是群落抵御外界压力而维持原来的稳定状态[29]。根据该理论,土壤微生物群落可认为是一个动态变化的自组织系统,通过遗传来维持其组成的稳定性,通过演替而适应外界环境。而由于不同微生物对于环境条件的耐受程度不同,可以定义微生物表现出抵抗力(x)和恢复力(y)两种属性(x,y),分别指微生物群落抵御外界特定环境压力维持自身稳定的能力和回复到原始稳定状态的能力[29]。依据抵抗力/恢复力的高(+)低(-),可将微生物分为四种类型,即抵抗力高恢复力高类型(+,+)、抵抗力高恢复力低类型(+,-)、抵抗力低恢复力高类型(-,+)和抵抗力低恢复力低类型(-,-)。基于这种设定,提出了时间和空间上微生物群落构建机制的概念模型(图1)。在时间格局中,假定微生物存在一个相对稳定的初始状态,当环境条件发生改变的时候,由于不同类群对该压力抵抗力和恢复能力的差异,各自的响应也不相同:(+,+)在最终稳定的群落中会占据优势;(+,-)保持不变;(-,+)先减少后增加并恢复原状;(-,-)可能被新出现的适应性突变体取代。在空间格局中,则可以假定微生物在空间上的分布是均匀的,微生物不存在地理分布格局(即零假设)。由于不同类群在同一环境梯度上的响应不同,因此稳定之后实际分布的群落中:(+,+)在最终稳定的群落中会占据优势;(+,-)和(-,+)的类群最终都能保持相对不变;(-,-)可能被新出现的适应性类群(突变体)取代。

图1 时空尺度上的环境压力下土壤微生物群落构建的概念模型 Fig.1 Conceptual model of soil microbial community assembly under certain environmental stress within spatiotemporal scales(引自[29],有修改)抵抗力高恢复力高(+,+)、抵抗力高恢复力低(+,-)、抵抗力低恢复力高(-,+)和抵抗力低恢复力低(-,-)四种不同类型的微生物分别用实心三角形、空心三角形、空心圆形和实心圆形表示
这一概念模型解释了耐受性不同的微生物类群对同一环境压力的响应,而微生物群落整体的稳定则是不同类群的抵抗力和恢复力的加和。用多样性-稳定性理论解释微生物时空演变特征的最终结果,则是(1)在时间水平上,微生物的群落随着环境条件的变化而发生变化,比如在经历了长期重金属胁迫的环境压力之后耐受性微生物类群明显增加[10];(2)在空间水平上,土壤微生物表现出与环境梯度密切相关的空间分布特征,比如我们在对全国不同森林土壤微生物的调查发现,亚热带常绿阔叶林土壤中耐受低pH环境压力的酸杆菌类群比例高于其他森林土壤(未发表数据)。
上述三种理论基础分别从不同角度阐述了微生物群落构建的机制,反映了自然科学研究 “形而下”的发展规律。这三种理论基础在生态学发展过程中都占据了重要的位置。其中,又以生态位理论/中性理论发展历史最长、衍生的理论体系最为丰富,因此生态位/中性理论可以说是现代生态学理论的重要基石。相比而言,过程理论和多样性-稳定性理论用于解释群落构建机制的时间较短(2010年以后),所以目前还没有很多的相关理论发展起来,但在将来相信会有更多的延伸。
3 土壤微生物群落时空演变的尺度效应
尽管在群落构建理论中没有明确限定范围,但自然界的所有格局与规律都具有尺度依赖性[30],我们观察到的土壤微生物时空分布规律都是建立于特定的时间和空间尺度乃至分类尺度上的。因此,尺度效应是地理学与生态学等研究中必须考虑的关键问题。在不同的研究尺度上,驱动微生物群落构建机制的差异导致了群落演变规律的变化。
3.1 空间尺度
空间尺度可以简单区分为大尺度、中尺度和小尺度[20],也可以根据具体的研究范围划分为微小尺度、局域尺度、生态系统尺度、区域尺度、洲际尺度、全球尺度等。微生物在不同空间尺度上的分布特征是土壤微生物地理研究的主要方面。由于土壤微生物的大小在微米级以下,因此其空间分布可以涵盖不同的研究尺度:在厘米以下的微小尺度范围内,土壤孔隙(团聚体)结构、微生物间的相互作用、根际效应等[31]就可以导致微生物分布格局的差异;在米到公里的范围内[12],土壤的异质性、植被的差异、地形等因素对微生物的分布产生影响;在几百乃至上千公里的更大的尺度上[32],土壤发育条件、气候、乃至地理隔离等条件都影响到微生物的空间分布。
不同空间尺度下土壤微生物的分布特征可能不同。最典型的例子是局域尺度(几百米到几千米)上海拔分布特局和区域尺度上(几百公里到几千公里)纬度分布格局的差异。海拔梯度和纬度梯度具有相似的环境条件,即随着海拔/纬度的升高,温度呈现线性降低并伴随降水、植被、土壤等环境条件的变化。海拔梯度上的植物多样性存在着降低或者单峰格局,而土壤微生物随海拔梯度的单峰或者降低格局也被证实[33-34]。在大的区域尺度上,生物多样性的纬度格局是生态学中一个重要的问题[35]。物种丰富度由热带到寒带逐渐降低这一基本格局对于大型动植物是确定的[36],但是对无脊椎的动物,包括蚯蚓、甲虫等则缺乏适用性。现有的证据表明,生物多样性的纬度格局似乎存在一种尺度效应,即体积小的生物在中纬度地区的多样性最高[37]。由于缺乏在完整的纬度梯度上的详实数据,土壤微生物的全球地理分布格局尚不确定。但根据现有的部分结果推测[38],土壤微生物的纬度多样性格局很可能与大型动植物不一致。
从群落构建理论角度看,造成不同尺度下土壤微生物群落空间分布格局差异的原因在于群落构建机制的差异。这一解释以生态位/中性理论的应用最为广泛:目前生态学家趋向于认为生态位理论和中性理论都对群落构建产生影响,但这种影响具有明显的尺度依赖性,即小尺度下生态位理论的作用大于中性理论,而大尺度下中性理论的作用大于生态位理论[7]。在认可微生物扩散限制的前提下,根据过程理论的体系,扩散过程在小尺度比在大尺度更容易实现。
3.2 时间尺度
土壤微生物群落随着时间的演变同样具有明显的尺度效应。时间尺度上的研究跨度可以从小时、天、月到季节、年乃至更久。在小的时间尺度上,微生物群落动态的驱动因子是间歇性脉冲式的,从而引发微生物的快速响应。比如,在经历了长期的干旱之后,降水导致的土壤含水量的增加会发生在几分钟到几个小时或者几天的时间里,这使得某些亲缘关系密切的特定的微生物突然复苏并持续增多,这种快速响应与氮矿化和土壤二氧化碳释放等的变化密切相关[39]。而在稍长的时间内,土壤微生物群落对环境条件的响应具有明显的季节性差异:春季和秋季对土壤养分的响应最为积极,而夏季与植被之间关系密切[40]。
由于大的历史时间具有不可重复性,因此单从生态学的角度看,对于土壤微生物群落时间演替的研究只能在较短的时间尺度(天、月、年)上进行,而无法对土壤微生物群落在大的时间尺度上的演变规律进行直接研究。这种思路和技术上的限制阻碍了对大尺度上微生物群落的时间演替规律与驱动机制的理解。土壤学理论为这种困境提供了部分解决方案。道库恰耶夫的成土理论指出,土壤是由特定地形下的成土母质,经过气候和生物长时间的作用而形成的[41]。而微生物是土壤发育初始阶段的主要生物驱动力。因此,可以用空间代替时间的方法,通过对不同的土壤演替序列(Chronosequence)中微生物群落的分析来反映其在大的时间尺度上的演替规律。例如,可以冰川退缩后暴露出来的母质作为土壤发育初始过程的起点,通过测量冰川的退化距离而计算土壤的发育时间,通过对不同发育程度的土壤中微生物群落的分析来揭示微生物群落在大的时间尺度上的演替规律与机制。鉴于土壤发育的基本过程,可以推测大致上存在这样的时间格局:在演替的初始阶段,土壤食物网络由简单的异养微生物、光合以及固氮微生物组成;随着时间推移,群落越来越复杂并趋于稳定,伴随食物链延长,植物对通过菌根真菌获取养分的依赖性增加[42]。
对一些具有特殊地质历史背景的生境的研究在一定程度上也能反映出大的时间尺度上土壤微生物群落的演变特征。例如,在地质历史上青藏高原南部大概在700万年之前从海底隆起,是非常年轻的山脉[43]。对青藏高原南部色季拉山土壤微生物群落组成的研究结果表明[34],土壤古菌中海洋底栖微生物类群Marine Benthic Group A (MBGA)的相对多度非常高(平均大于60%),表明经历几百万年之后土壤微生物群落可能仍然遗存了部分对于海洋环境的适应特性,历史效应可能在当代仍得以保留。这一结果暗示了在长期进化历史中,中性过程对微生物群落演替的影响仍然很大。此外,对古老冰芯的研究也是反映微生物群落在地质历史时期演变特征的有效途径,由于与外界环境之间缺乏基因交流,这一生境是反映微生物进化的比较理想的模式[44]。
4 问题与展望
在陆地生态系统中,土壤作为地上和地下生物群落联系的枢纽,是各种复杂生态过程发生的场所。动植物与微生物群落之间、微生物群落内部、微生物群落与土壤环境之间的相互作用,以及生物与环境条件本身在时空尺度上的变化,都会对微生物群落的时空演变产生影响。从哲学角度上说,我们永远不可能准确而完整地认识微生物群落时空演变的所有特征[45]。因此,根据当前微生物群落演变的理论以及其在时空尺度上的驱动机制,发展相关生态模型来简化研究问题并突出主要矛盾就显得尤为迫切,例如通过对决定微生物时空演变的因子解析,建立物种分布模型,最终实现对大的空间尺度和时间尺度上的微生物群落演变规律的预测[46]。
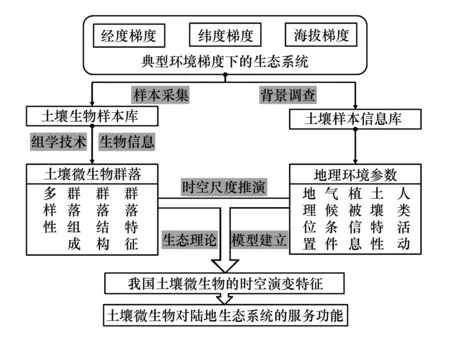
图2 我国土壤微生物时空演变格局的研究框架 Fig.2 A research framework on the spatiotemporal patterns of soil microbes in China
土壤微生物时空演变研究是以对微生物功能的关注为根本驱动力的,其主要内涵是土壤微生物维持陆地生态系统生物多样性和功能所发挥的作用。而这一问题实际指向的是植物和微生物的共进化演变格局。土壤微生物和地上植物之间存在着紧密的相互作用:植物影响微生物的主要途径包括对土壤的营养输入和对内生菌、根际微生物的直接影响[47];微生物对植物的影响主要为分解凋落物,通过调控土壤营养来改变植被的生长和群落结构。氮素是土壤中的常见限制性营养元素,而植物本身不能固氮,因此固氮微生物对植物初级生产力有着更加明显的影响。植物的内生菌、根际微生物可以更直接地影响植物,也可以通过介导植物和非根际微生物之间的物质和能量交流来影响植物和非根际微生物。两者相互影响和相互改变,形成正反两方面的反馈,共同驱动了各自群落结构的演变,也影响了土壤元素循环过程和组分。因此,在当前全球气候变化背景下,关注土壤微生物和植物的格局及其在时空尺度上演变规律的联系,有助于我们更为准确地评估气候变化的环境和生态效应。
我国幅员辽阔,不同地区气候差异显著。沿着东西方向的经度梯度降水发生明显变化,草原、灌丛、荒漠等生态系统植被差异明显;南北方向的纬度梯度上涵盖了由热带雨林到寒温带落叶针叶林在内的完整森林带谱,是研究纬度生物多样性格局的理想环境;垂直方向上有着丰富的山地生态系统,更是有着青藏高原这样的“世界屋脊”。不同的生态系统自然变异及受人为干扰程度各不相同,这就为研究不同尺度下气候环境因子与人为干扰因子对土壤微生物时空分布的影响提供了理想环境。在这些环境梯度下,丰富的气候条件和土壤类型孕育了纷繁复杂的土壤微生物,使得中国在土壤微生物的时空分布研究中具有独特的研究背景。同时在从南到北的热量梯度样带、从东到西的降雨梯度样带上具备了长达30年的长期定位试验站,这些都为研究土壤微生物随时间上的演替规律提供了理想平台。在客观条件上,高通量测序技术的迅猛发展为大规模的土壤微生物群落调查提供了技术支撑;另一方面,通过对土壤样本采集、样本处理与保藏等一系列过程的统一规范从而建立土壤生物样本库[48],为大的空间尺度以及时间序列上的土壤微生物群落演变研究提供基础。通过这些努力,有望阐明我国主要生态系统中土壤微生物的时空分布格局、多样性的形成机制及其潜在生态功能,为提升生态系统生产力、维持生态系统服务提供理论依据(图2)。
[1] 贺纪正, 陆雅海, 傅伯杰. 土壤生物学前沿. 北京: 科学出版社, 2015.
[2] 贺纪正, 葛源. 土壤微生物生物地理学研究进展. 生态学报, 2008, 28(11): 5571-5582.
[3] Bell T, Ager D, Song J I, Newman J A, Thompson I P, Lilley A K, Van Der Gast C J. Larger islands house more bacterial taxa. Science, 2005, 308(5730): 1884-1884.
[4] Cho J C, Tiedje J M. Biogeography and degree of endemicity of fluorescent Pseudomonas strains in soil. Applied and Environmental Microbiology, 2000, 66(12): 5448-5456.
[5] Green J, Bohannan B J M. Spatial scaling of microbial biodiversity. Trends in Ecology & Evolution, 2006, 21(9): 501-507.
[6] Prosser J I, Bohannan B J M, Curtis T P, Ellis R J, Firestone M K, Freckleton R P, Green J L, Green L E, Killham K, Lennon J J, Osborn A M, Solan M, Van Der Gast C J, Young J P W. The role of ecological theory in microbial ecology. Nature Reviews Microbiology, 2007, 5(5): 384-392.
[7] 曹鹏, 贺纪正. 微生物生态学理论框架初探. 生态学报, 2015, 35(22): 1-14.
[8] 牛克昌, 刘怿宁, 沈泽昊, 何芳良, 方精云. 群落构建的中性理论和生态位理论. 生物多样性, 2009, 17(6): 579-593.
[9] O′Malley M A. The nineteenth century roots of ′everything is everywhere′. Nature Reviews Microbiology, 2007, 5(8): 647-651.
[10] 李博. 生态学. 北京: 高等教育出版社, 2000.
[11] Cao P, Zhang L M, Shen J P, Zheng Y M, Di H J, He J Z. Distribution and diversity of archaeal communities in selected Chinese soils. FEMS Microbiology Ecology, 2012, 80(1): 146-158.
[12] Wang J T, Zheng Y M, Hu H W, Zhang L M, Li J, He J Z. Soil pH determines the alpha diversity but not beta diversity of soil fungal community along altitude in a typical Tibetan forest ecosystem. Journal of Soils and Sediments, 2015, 15(5): 1224-1232.
[13] Vandamme P, Pot B, Gillis M, De Vos P, Kersters K, Swings J. Polyphasic taxonomy, a consensus approach to bacterial systematics. Microbiological Reviews, 1996, 60(2): 407-438.
[14] 贺纪正, 袁超磊, 沈菊培, 张丽梅. 土壤宏基因组学研究方法与进展. 土壤学报, 2012, 49(1): 155-164.
[15] Simpson E H. Measurement of diversity. Nature, 1949, 163(4148): 688-688.
[16] Wittebolle L, Marzorati M, Clement L, Balloi A, Daffonchio D, Heylen K, De Vos P, Verstraete W, Boon N. Initial community evenness favours functionality under selective stress. Nature, 2009, 458(7238): 623-626.
[17] Faith D P. Conservation evaluation and phylogenetic diversity. Biological Conservation, 1992, 61(1): 1-10.
[18] Lozupone C A, Hamady M, Kelley S T, Knight R. Quantitative and qualitative β diversity measures lead to different insights into factors that structure microbial communities. Applied and Environmental Microbiology, 2007, 73(5): 1576-1585.
[19] 马克明. 物种多度格局研究进展. 植物生态学报, 2003, 27(3): 412-426.
[20] Martiny J B H, Bohannan B J M, Brown J H, Colwell R K, Fuhrman J A, Green J L, Horner-Devine M C, Kane M, Krumins J A, Kuske C R, Morin P J, Naeem S, Øvreås L, Reysenbach A -L, Smith V H, Staley J T. Microbial biogeography: putting microorganisms on the map. Nature Reviews Microbiology, 2006, 4(2): 102-112.
[21] Ge Y, He J Z, Zhu Y G, Zhang J B, Xu Z H, Zhang L M, Zheng Y M. Differences in soil bacterial diversity: driven by contemporary disturbances or historical contingencies?. The ISME Journal, 2008, 2(3): 254-264.
[22] Lin W, Wang Y Z, Li B, Pan Y X. A biogeographic distribution of magnetotactic bacteria influenced by salinity. The ISME Journal, 2012, 6(2): 475-479.
[23] 朱璧如, 张大勇. 基于过程的群落生态学理论框架. 生物多样性, 2011, 19(4): 389-399.
[24] Vellend M. Conceptual synthesis in community ecology. The Quarterly Review of Biology, 2010, 85(2): 183-206.
[25] Hanson C A, Fuhrman J A, Horner-Devine M C, Martiny J B H. Beyond biogeographic patterns: processes shaping the microbial landscape. Nature Reviews Microbiology, 2012, 10(7): 497-506.
[26] Adams R I, Miletto M, Taylor J W, Bruns T D. Dispersal in microbes: fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. The ISME Journal, 2013, 7(7): 1262-1273.
[27] Hu H W, Zhang L M, Dai Y, Di H J, He J Z. pH-dependent distribution of soil ammonia oxidizers across a large geographical scale as revealed by high-throughput pyrosequencing. Journal of Soils and Sediments, 2013, 13(8): 1439-1449.
[28] Wang J J, Shen J, Wu Y C, Tu C, Soininen J, Stegen J C, He J Z, Liu X Q, Zhang L, Zhang E L. Phylogenetic beta diversity in bacterial assemblages across ecosystems: deterministic versus stochastic processes. The ISME Journal, 2013, 7(7): 1310-1321.
[29] 贺纪正, 李晶, 郑袁明. 土壤生态系统微生物多样性——稳定性关系的思考. 生物多样性, 2013, 21(4): 411-420.
[30] 吕一河, 傅伯杰. 生态学中的尺度及尺度转换方法. 生态学报, 2001, 21(12): 2096-2105.
[31] Marschner P, Yang C H, Lieberei R, Crowley D E. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biology and Biochemistry, 2001, 33(11): 1437-1445.
[32] Fierer N, Jackson R B. The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(3): 626-631.
[33] Singh D, Takahashi K, Kim M, Chun J, Adams J M. A hump-backed trend in bacterial diversity with elevation on Mount Fuji, Japan. Microbial Ecology, 2012, 63(2): 429-437.
[34] Wang J T, Cao P, Hu H W, Li J, Han L L, Zhang L M, Zheng Y M, He J Z. Altitudinal distribution patterns of soil bacterial and archaeal communities along Mt. shegyla on the tibetan plateau. Microbial Ecology, 2014, 69(1): 135-145.
[35] Willig M R, Kaufman D M, Stevens R D. Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis. Annual Review of Ecology, Evolution, and Systematics, 2003, 34: 273-309.
[36] Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220-227.
[37] De Deyn G B, Van Der Putten W H. Linking aboveground and belowground diversity. Trends in Ecology & Evolution, 2005, 20(11): 625-633.
[38] Tedersoo L, Bahram M, Põlme S, Kõljalg U, Yorou N S, Wijesundera R, Ruiz L V, Vasco-Palacios A M, Thu P Q, Suija A, Smith M E, Sharp C, Saluveer E, Saitta A, Rosas M, Riit T, Ratkowsky D, Pritsch K, Põldmaa K, Piepenbring M, Phosri C, Peterson M, Parts K, Pärtel K, Otsing E, Nouhra E, Njouonkou A L, Nilsson R H, Morgado L N, Mayor J, May T W, Majuakim L, Lodge D J, Lee S S, Larsson K H, Kohout P, Hosaka K, Hiiesalu I, Henkel T W, Harend H, Guo L D, Greslebin A, Grelet G, Geml J, Gates G, Dunstan W, Dunk C, Drenkhan R, Dearnaley J, De Kesel A, Dang T, Chen X, Buegger F, Brearley F Q, Bonito G, Anslan S, Abell S, Abarenkov K. Global diversity and geography of soil fungi. Science, 2014, 346(6213): 1256688.
[39] Placella S A, Brodie E L, Firestone M K. Rainfall-induced carbon dioxide pulses result from sequential resuscitation of phylogenetically clustered microbial groups. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(27): 10931-10936.
[40] Regan K M, Nunan N, Boeddinghaus R S, Baumgartner V, Berner D, Boch S, Oelmann Y, Overmann J, Prati D, Schloter M, Schmitt B, Sorkau E, Steffens M, Kandeler E, Marhan S. Seasonal controls on grassland microbial biogeography: Are they governed by plants, abiotic properties or both?. Soil Biology and Biochemistry, 2014, 71: 21-30.
[41] Jenny H. Factors of Soil Formation: A System of Quantitative Pedology. New York: Dover publications, 1941.
[42] Bardgett R D, Van Der Putten W H. Belowground biodiversity and ecosystem functioning. Nature, 2014, 515(7528): 505-511.
[43] Wang C S, Zhao X X, Liu Z F, Lippert P C, Graham S A, Coe R S, Yi H S, Zhu L D, Liu S, Li Y L. Constraints on the early uplift history of the Tibetan Plateau. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(13): 4987-4992.
[44] Price P B. A habitat for psychrophiles in deep Antarctic ice. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97(3): 1247-1251.
[45] Jørgensen S E, Bendoricchio G. Fundamentals of Ecological Modelling. 3th ed. UK: Elsevier, 2001.
[46] Fierer N, Ladau J, Clement J C, Leff J W, Owens S M, Pollard K S, Knight R, Gilbert J A, McCulley R L. Reconstructing the microbial diversity and function of pre-agricultural tallgrass prairie soils in the united states. Science, 2013, 342(6158): 621-624.
[47] Wardle D A, Bardgett R D, Klironomos J N, Setälä H, Van Der Putten W H, Wall D H. Ecological linkages between aboveground and belowground biota. Science, 2004, 304(5677): 1629-1633.
[48] Cary S C, Fierer N. The importance of sample archiving in microbial ecology. Nature Reviews Microbiology, 2014, 12(12): 789-790.
Mechanisms of community organization and spatiotemporal patterns of soil microbial communities
HE Jizheng*, WANG Juntao
StateKeyLaboratoryofUrbanandRegionalEcology,ResearchCenterforEco-EnvironmentalSciences,ChineseAcademyofSciences,Beijing100085,China
As an important component of terrestrial ecosystems, soil microbial communities are directly or indirectly involved in almost all soil ecological processes, including material cycles, energy conversion, and pollutant transformation. Studies of the spatiotemporal patterns of changes in soil microbial communities and the underlying mechanisms would help us determine the basic principle of microbial evolution, and would provide a scientific basis to predict the response, adaptation, and feedback of the microbial communities to environmental changes. In this review, we discuss the definition, analysis methods, and indices of soil microbial community. We argue that community could be the basic unit in macro-ecology and microbial ecology. Studies of community organization mechanisms are at the core of both the macro-and microbial ecology. Based on the community organization theories in ecology, ecological niche and neutral theories, process-based theories, and diversity-stability theories were introduced and applied to understand the soil microbial spatiotemporal patterns and the scale effects. A framework for the study of spatiotemporal patterns of changes in soil microbial communities was proposed based on the community organization theories under different spatial and temporal scales.
soil microbial community; microbial biogeography; microbial evolution; biodiversity; community organization
国家自然科学基金(41230857);典型生态系统土壤微生物生态空间特征评价(STSN-21-02)
2015-06-06; < class="emphasis_bold">网络出版日期:
日期:2015-07-29
10.5846/stxb201506061143
*通讯作者Corresponding author.E-mail: jzhe@rcees.ac.cn
贺纪正,王军涛.土壤微生物群落构建理论与时空演变特征.生态学报,2015,35(20):6575-6583.
He J Z, Wang J T.Mechanisms of community organization and spatiotemporal patterns of soil microbial communities.Acta Ecologica Sinica,2015,35(20):6575-6583.
