Cry1Ab蛋白对拟环纹豹蛛生长发育的影响
2015-01-20李秦晋魏宝阳湖南农业大学生物科学技术学院湖南长沙4028湖南体育职业学院湖南长沙4009
李秦晋, 魏宝阳 (.湖南农业大学生物科学技术学院,湖南长沙 4028;2.湖南体育职业学院,湖南长沙 4009)
Cry1Ab蛋白对拟环纹豹蛛生长发育的影响
李秦晋1,2, 魏宝阳1(1.湖南农业大学生物科学技术学院,湖南长沙 410128;2.湖南体育职业学院,湖南长沙 410019)
[目的] 探明Cry1Ab蛋白通过培养基-果蝇-拟环纹豹蛛这一食物链的传递,最终在拟环纹豹蛛(Pardosapseudoannulata)体内的富集情况以及毒蛋白随食物链的传递在各营养级中的变化规律。[方法]采用酶联免疫技术(ELISA)测定Cry1Ab蛋白在拟环纹豹蛛和果蝇中的含量。[结果]Cry1Ab蛋白对不同龄期拟环纹豹蛛的体长、背甲宽、中眼域宽、腿长等生长指标不存在显著性差异,但对体重产生一定的影响,使得其体重略有下降的趋势。在0~10 d Cry1Ab蛋白在拟环纹豹蛛体内不断累积,但10 d后拟环纹豹蛛体内Cry1Ab蛋白含量逐渐降低。[结论] Cry1Ab蛋白在拟环纹豹蛛体内具有富集效应。
拟环纹豹蛛;果蝇;Cry1Ab蛋白;食物链
尽管转基因抗虫作物可以带来巨大效益[1-2],但关于转基因作物的生态安全性评价受到了世界各国政府部门和科技人员的重视,许多学者对转基因作物的利弊及其潜在的生态环境问题进行了分析[3-8]。其中抗虫转Bt基因作物对害虫天敌等非靶标昆虫的潜在影响是当今转基因生物安全性的研究热点之一[9-13]。
拟环纹豹蛛(Pardosapseudoannulata)是稻田蜘蛛中的优势种群[14],农田作物害虫最主要的捕食性天敌之一[15-22]。作为害虫的重要捕食性天敌,Bt蛋白会随水稻-害虫-蜘蛛这一食物链传递,水稻中的Bt毒蛋白有可能会在蜘蛛体内积累,对其繁殖、生长和发育产生潜在影响。鉴于此,笔者模拟水稻食物链,构建实验室Bt培养基-果蝇-蜘蛛这一食物链,以此探明Bt毒蛋白沿食物链的传递对蜘蛛毒害的生态效应,明确这种效应对蜘蛛生长、发育和繁殖的影响,同时也为Bt作物生态安全性评价提供参考。
1 材料与方法
1.1 试验材料在田间采集拟环纹豹蛛带回室内饲养备用。
1.2 主要试剂Cry1Ab蛋白标准品(上海佑隆生物科技有限公司),EnviroLogix Cry1Ab/Cry1Ac(ELISA)蛋白检测试剂盒(上海佑隆生物科技有限公司),SOD、GSH-Px试剂盒(南京建成生物工程研究所),其他试剂均为国产分析纯。
1.3 主要仪器电热恒温水浴锅(DK-98-IIA),天津市泰斯特仪器有限公司;可见分光光度计(V-5000型),上海元析仪器有限公司;台式低温冷冻离心机,日本日立仪器有限公司;酶标仪(MK3型),热电(上海)仪器有限公司;电子分析天平(TMP-510),湘仪天平仪器设备有限公司;恒温恒湿培养箱(LRH-250-S),广东省医疗器械厂。
1.4 试验方法
1.4.1拟环纹豹蛛的喂养。在同一非Bt水稻田块采集带有卵囊的拟环纹豹蛛,待卵囊孵化5 d后,将幼蛛接入2.5 cm的大型试管内(保证试验蜘蛛均来自同一卵囊),每管一头,进行单头饲养,每支试管中均置一团湿润的脱脂棉,并每天加水3~4滴于脱脂棉上,为其提供水分。试验组为每天喂食在含有Cry1Ab蛋白培养基上饲养5~10 d的果蝇,正常组为每天喂食在普通培养基上饲养5~10 d的果蝇。
每只幼蛛蜕皮后2 d,对其进行称重,并从中取出2只幼蛛用75%的乙醇浸泡2h后进行生理指标的测定。
1.4.2Cry1Ab蛋白的提取。分别将试验组和正常组的拟环纹豹蛛麻醉,每组取3只拟环纹豹蛛,将其麻醉后称重,待其将要苏醒时立即置于匀浆器中,加入250 μl提取液,在冰浴中将拟环纹豹蛛研磨成匀浆。将匀浆后的样品液于高速冷冻离心机中以4 000 r/min(4 ℃)离心10 min,取上清液用于ELISA检测Cry1Ab蛋白含量,-20 ℃储存备用。
1.4.3Cry1Ab蛋白含量测定(ELISA)。
1.4.3.1Cry1Ab蛋白标准曲线的制作。取标准CrylAb蛋白用样品提取缓冲液依次稀释成1、2、4、6、8 ng 5个梯度,按照试剂盒说明的方法进行测定,制作标准曲线。
1.4.3.2酶联免疫技术(ELISA)。取5 μl待测样品,加入45 μl提取液,将待测样品稀释10倍,在微孔板的每孔中加入50 μl酶标记Cry1Ab/Cry1Ac后,立刻加入50 μl空白对照(提取液)、阳性对照和样品稀释液,轻摇混匀20~30 s;微孔板上加盖防止液体蒸发,室温下温育2 h后小心地揭去盖,摇匀微孔内液体,并加入洗涤缓冲液冲洗,用力弃去微孔中的液体,重复3次;接着在每孔中加入300 μl洗涤液洗涤,并在纱布上拍干,每孔中加入100 μl底物溶液,轻摇混匀20~30 s,微孔板上加盖,室温下温育15~30 min,每孔中加入100 μl 终止液,充分混匀后用酶标仪在450 nm波长下读取OD值,用测试孔OD-空白孔OD,对照标准曲线得到Cry1Ab蛋白的含量(ng),将结果换算成可溶性蛋白中含有的Cry1Ab蛋白量(ng/mg)。
1.5 数据分析采用WPS Excel软件进行数据处理 ,并用SPSS 17.0软件对数据进行统计分析。数据均以平均值±标准误(Mean±SE)表示。
2 结果与分析
2.1 生长指标由表1可知,Bt蛋白对不同龄期拟环纹豹蛛的体长、中眼域宽和背甲宽等生长指标不存在显著性差异。

表1 不同龄期拟环纹豹蛛生长指标 cm
2.2 重量由表2可知,Bt蛋白对不同龄期拟环纹豹蛛的重量有一定影响,使得其体重略有下降趋势。在5龄时,体重的差异性最大(4龄、5龄存在显著性差异)。
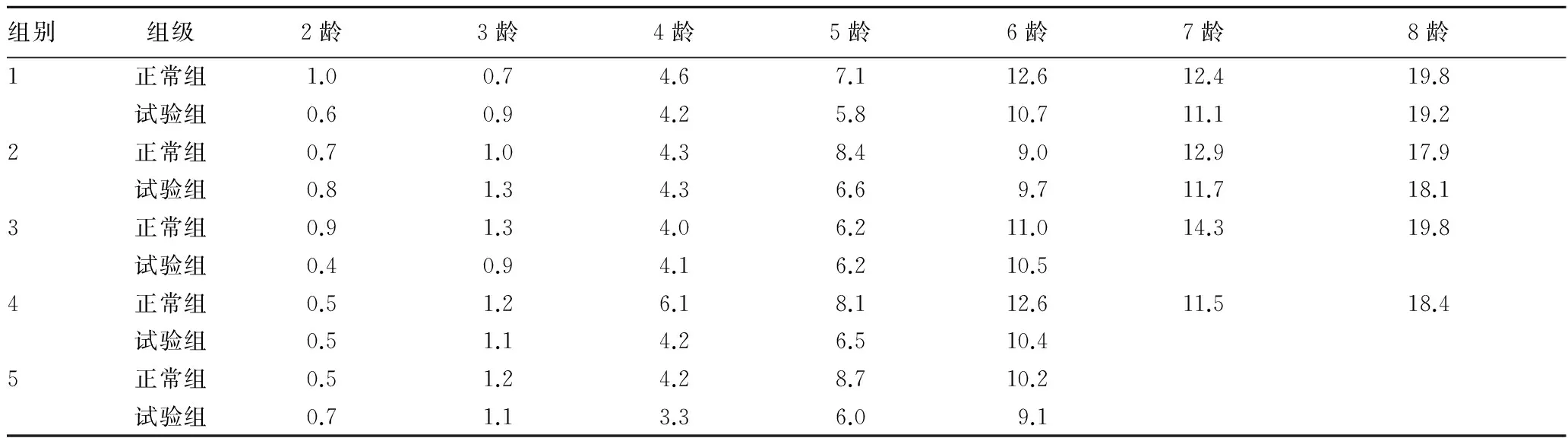
表2 不同龄期拟环纹豹蛛重量 mg
2.3 蛋白含量由表3可知,在0~10 d Cry1Ab蛋白在果蝇体内的含量随着培养时间的延长而不断增加,表明Cry1Ab蛋白的确在果蝇体内有富集作用,但在10 d后Cry1Ab蛋白含量下降,但在15 d时含量仍高于对照。 由表4可知,第1~7天Cry1Ab蛋白在拟环纹豹蛛体内均具富集作用(P<0.05),到第7天时达到最高值,达极显著差异(P<0.01),然后逐渐降低,但仍显著高于对照(P<0.05)。

表3 不同培养时间果蝇体内Cry1Ab蛋白含量
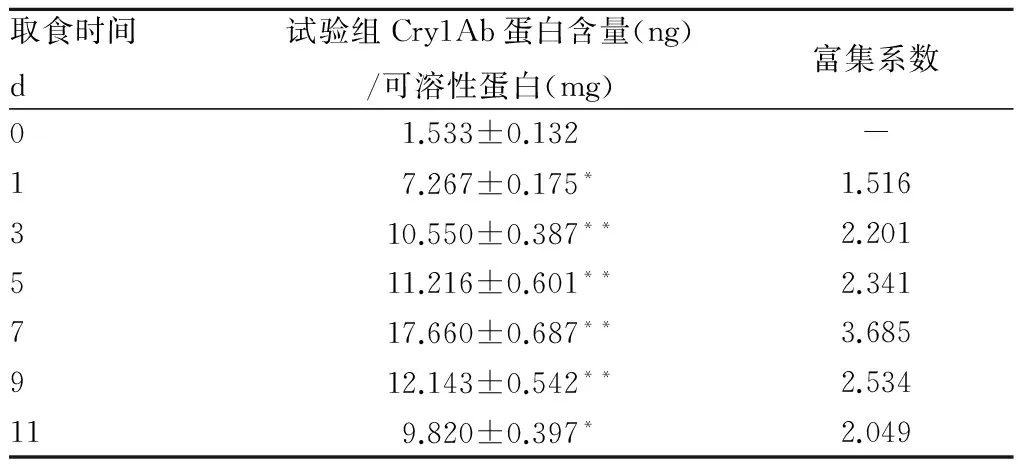
表4 不同龄期拟环纹豹蛛体内Cry1Ab蛋白含量
注:*表示差异显著(P<0.05),**表示差异极显著(P<0.01)。
3 讨论
有关抗虫转Bt基因作物对其非靶标植食者及天敌的正面或负面影响已有较多报道[2,23-25],但仅分析了温室或田间条件下转基因作物对供试对象生物学、生态学特性的影响,而极少探讨Bt杀虫蛋白能否在转基因植物-非靶标植食者-天敌三者间进行转移与富集。该试验对培养基中的CrylAb毒蛋白在果蝇体内的积累作用进行了研究,发现CrylAb毒蛋白在果蝇体内的含量随取食时间的延长先增加后下降。而喂食含有CrylAb毒蛋白果蝇的拟环纹豹蛛体内CrylAb毒蛋白也呈先增加后下降的趋势。CrylAb毒蛋白能够在果蝇及拟环纹豹蛛体内产生富集效应,而后蛋白含量下降,可能是由于随取食时间的延长,CrylAb毒蛋白对果蝇和拟环纹豹蛛造成了影响,使个体生活力下降,取食量下降,摄入的CrylAb毒蛋白较少所致;也可能是由于培养基中的CrylAb蛋白已耗尽,而果蝇体内的CrylAb毒蛋白被降解而造成。目前的研究是在较短时间内得出的,长期作用结果如何,以及具体机理尚有待进一步跟踪研究。
该试验以Bt非靶标生物果蝇作为蜘蛛的食物来源,探讨Bt毒素在Bt培养基-果蝇-蜘蛛这一食物链中的传递和累积规律,以及Bt蛋白的富集对蜘蛛生长发育的影响,可在一定程度上揭示转Bt蛋白对稻田蜘蛛长期安全性的影响。其次,通过同一母体的幼蛛在Bt毒蛋白胁迫下对其生长发育影响,最大可能地消除蜘蛛个体间的差异,使得试验更具参考意义
[1] HILDER V A,BOULTER D.Genetic engineering of crop plants for insect resistance-a critical review[J].Crop Prot,1999,18:177-191.
[2] SHELTON A M,ZHAO J Z,ROUSH R T.Economic,ecologic,food safety,and social consequences of the development of Bt transgenic plants[J].Ann Rev Entomol,2002,47:845-881.
[3] 修伟明,刘红梅,李刚,等.转Bt基因抗虫玉米发展状况及对非靶标昆虫的影响[J].玉米科学,2009,17(4):1-6.
[4] WOLFENBARGER L L,NARANJO S E,LUNDGREN,J G,et al.Bt crop effects on functional guilds of non-target arthropods:a meta- analysis[J].PLoS One,2008,3(5):2118.
[5] 钱迎倩,魏伟,桑卫国,等.转基因作物对生物多样性的影响[J].生态学报,2001,21(3):337-343.
[6] 史晓利,杨益众,蔡建华,等.Bt毒蛋白在转基因抗虫玉米中的表达及在亚洲玉米螟中的转移积累[J].应用生态学报,2009,20(11):2773-2777.
[7] CHEN M,ZHAO J Z,YE G Y,et al.Impact of insect-resistant transgenic rice on target insect pests and non-target arthropods in China[J].Insect Science,2006,13:409-420.
[8] CHEN M,SHELTONH A,YE G Y.Insect-resistant genetically modified rice in China:from research to commercialization[J].Annual Review of Entomology,2011,56:81-101.
[9] POPPY G M.Corps:environmental risks and non-target effects[J].Trends Plant Sci,2000,5:4-6.
[10] 李丽莉,王振营,何康来,等.转基因抗虫作物对非靶标昆虫的影响[J].生态学报,2004,24(8):1797-1806.
[11] ROMEIS J,MEISSLE M,BIGLER F.Transgenic crops expressing Bacillus thuringiensis toxins and biological control[J].Nat Biotechnol,2006,24:63-71.
[12] MARVER M,MCCREEDY C,REGETZ,et al.A meta-analysis of effects of Bt cotton and maize on non-target invertebrates[J].Science,2007,316:1475-1477.
[13] SHELTON A M,SEARS M K.The monarch butterfly controversy:scientific interpretations of a phenomenon[J].The Plant Journal,2001,27(6):483-488.
[14] ANDREA CHIPMAN.Fears over Europe’s GM crop plan[J].Nature,2010,466:542-543.
[15] 刘雨芳,古德祥,张古忍.广东省稻田生态系统中蜘蛛群落多样性研究[J].蛛形学报,2003,12(1):27-31.
[16] 孙继英,彭光旭,胡波,等.拟环纹豹蛛种群遗传多样性与其生境的关系[J].应用生态学报,2007,18(5):1081-1085.
[17] 田俊策,陈洋,李兆亮,等.转cry1Ab基因水稻和两种杀虫剂对非靶标天敌拟环纹豹蛛免疫影响的评价[J].中国生物防治学报,2011,27(4):559-563.
[18] 王洪全.稻田蜘蛛利用研究[M].北京:科技文献出版社,1981:1-28.
[19] 宋大祥.中国农田蜘蛛[M].北京:农业出版社,1987:376.
[20] RIECHERT S E,BISHOP L.Pery control by an assemblage of generalist predators:Spiders in garden test systems[J].Ecology,1992,71:1441-1450.
[21] BARRION A T,LITSINGER J A.Riceland of South and South east Asia[M].Wallingford,UK:CAB International,1995:701.
[22] 王智.拟环纹豹蛛的生物生态学研究[J].昆虫学报,2007,50(9):927-932.
[23] HIGH S M,COHEN M B,SHU Q Y,et al.Achieving successful deployment of Bt rice[J].Trends Plant Sci,2004,6:286-292.
[24] O’CALLAHGANA M,GLAREl T R,BURGESS E P J,et al.Effects of Plnats genetically modified of insect resistance on non-target organisms[J].Annu Rev Entomol,2005,50:271-292.
[25] 刘志诚,叶恭银,傅强,等.转Cry1Ab基因水稻对拟水狼蛛捕食作用间接影响的评价[J].中国水稻科学,2003,27(2):175-178.
Effect of Cry1Ab Protein on Growth ofPardosepseudoannulata
LI Qin-jin1,2,WEI Bao-yang1(1.College of Bioscience & Biotechnology,Hunan Agriculture University,Changsha,Hunan 410128;2.Hunan Sports Vocational College,Changsha,Hunan 410019)
[Objective] The aim was to prove food chain transfer of Cry1Ab protein through the medium-fruit flies and intends toP.pseudoannulata, the final accumulate inP.pseudoannulatabody as well as toxic protein variation in each trophic level with the transfer of the food chain.[Method]Using enzyme-linked immunosorbent assay (ELISA) the content of Cry1Ab protein in theP.pseudoannulataandDrosophilamelanogasterwas determined.[Result]The results showed that,there was no signifant difference that Cry1Ab protein on body length,carapace width,the width of the eye domain,legs and other growth indicators ofP.pseudoannulatain different ages,but had a certain impact on body weight,so its weight was slightly a downward trend.Cry1Ab protein within 0-10 d accumulated inP.pseudoannulatabody,but after 10 days Cry1Ab protein content intended to decrease.[Conclusion] The study had shown the Cry1Ab protein enrichment effect ofP.pseudoannulatabody.
Pardosapseudoannulata;Drosophilamelanogaster; Cry1Ab protein; Food chain
国家自然科学基金项目(31472017);湖南省科技厅重点项目(2014FJ2003)。
李秦晋(1975- ),男,湖南益阳人,讲师,在读博士,从事农业生态学研究。
2015-03-31
S 188
A
0517-6611(2015)14-013-03
