不同光强下濒危植物伯乐树幼苗叶片光合生理参数比较
2015-01-08叶飞英陈子林郑伟成刘菊莲周钰鸿章剑玉潘成椿季国华
叶飞英,陈子林,郑伟成,刘菊莲,周钰鸿,章剑玉,潘成椿,季国华
(1.浙江九龙山国家级自然保护区 管理局,浙江遂昌323300;2.浙江大盘山国家级自然保护区 管理局,浙江 磐安322300;3.温州科技职业学院,浙江温州325006)
不同光强下濒危植物伯乐树幼苗叶片光合生理参数比较
叶飞英1,陈子林2,郑伟成1,刘菊莲1,周钰鸿2,章剑玉3,潘成椿1,季国华1
(1.浙江九龙山国家级自然保护区 管理局,浙江遂昌323300;2.浙江大盘山国家级自然保护区 管理局,浙江 磐安322300;3.温州科技职业学院,浙江温州325006)
为探讨伯乐树Bretschneidera sinensis幼苗的适宜光强环境,并为伯乐树幼苗的人工繁育及种群复壮提供参考,达到保护这一濒危物种的目的,比较研究了不同光强下(1 450.00,550.00和200.00 μmol·m-2·s-1)伯乐树幼苗的光合日变化、光响应曲线、二氧化碳响应曲线以及形态指标等相关参数。结果表明:气孔导度、蒸腾速率和光合速率的日变化均以全光处理下的较高;光强相对较高环境下(1 450.00和550.00 μmol·m-2·s-1)伯乐树幼苗叶片的表观光合速率、最大光合速率、最大羧化速率和羧化效率等光合参数值均显著较高(P<0.05),幼苗表现出良好的生长态势,说明伯乐树幼苗喜光。同时,不同光强处理下伯乐树幼苗叶片的光补偿点分别为21.22,19.93和14.95 μmol·m-2·s-1,其值均较低,说明伯乐树幼苗也较耐弱光。较高光强有利于伯乐树的光合作用,植株长势健壮,株高和地径相对较大。建议在进行伯乐树幼苗人工繁育时,不宜过度遮荫光;在原生地抚育时,应适当增加林窗的光照强度。图4表2参20
植物生理学;伯乐树;幼苗;发光强度;光合作用
伯乐树Bretschneidera sinensis又名钟萼木,伯乐树科Bretschneideraceae的落叶乔木,是中国特有的单种科植物,也是第三纪热带植物区系的孑遗种,对研究被子植物的系统发育及古地理、古气候等具有重大的科学价值。由于伯乐树星散分布,个体数量极少,被列为国家一级保护植物[1]。因此,亟须开展种群复壮的相关研究。近年来由于人为生态环境的破坏和砍伐,自然条件下结实少及野生动物取食,导致伯乐树自然状态更新较差,林分内很少见其幼树[2]。因此,伯乐树幼苗的繁育对其种群复壮具有重要意义。目前,关于伯乐树幼苗繁育主要集中在形态观察及生物学特性比较[2-7],而少有从生理生态学方面对其适宜生境的研究[3,8]。乔琦等[3,8]在野外调查中发现伯乐树1年生幼苗的夏季死亡率较高,死亡原因之一是遭受到亚热带夏季强光照造成的高温胁迫和干旱胁迫;同时研究认为,不同光环境下对夏季幼苗的存活率和生长量存在显著区别,表现在全光环境下易造成幼苗的死亡,而极度弱光环境会导致生长迟缓,只有适度遮光有利于幼苗的形态生长,其中自然遮光下的生长量各项指标(苗高、地茎、叶片数和侧枝数等)和幼苗成活率达到最优;而马冬雪等[7]认为,伯乐树播种育苗遮光对苗木生长没有明显影响,但在低海拔丘陵地区播种育苗需要适当遮光。然而,前者的试验在温室中进行,而后者并未从生理角度予以深入探讨。从植物对环境的适应性来看,随着生长光强的变化,植物能够在形态及生理方面产生可塑性反应,以适应变化的光环境[9]。由于濒危植物种群的分布区狭小、缺乏基因交流及生境长期的退化,生理生态学适应能力已经弱化,因此,有不少濒危植物的分布与其对光照的适应相关[10]。如Liu等[11]对濒危植物七子花Heptacodium miconioides的研究表明:其喜光而不耐强光,强光和弱光都对其光合作用以及抗氧化酶都非常不利,因而七子花从适应光的角度,选择中等海拔区域高大乔木较少的群落中,以满足其生长和生理的需要。张旺锋等[12]对濒危植物银杉Cathaya argyrophylla的研究表明:适度遮光有利于银杉抵御冬季光抑制。香果树Emmenopterys henryi为中性偏阳树种,喜温暖湿润气候,幼树喜阴湿,成树喜光,且母树在光照不足的条件下难以开花结实[13-14]。为了明确伯乐树幼苗对光强的响应特性,本研究从光合生理角度并结合形态指标研究了伯乐树幼苗对光强的响应,旨在探讨伯乐树幼苗的适宜光强环境,以期为伯乐树幼苗的人工繁育及其种群复壮提供参考。
1 材料与方法
1.1 植物材料及处理
种子于2010年10月采自浙江九龙山国家级自然保护区。清水洗去种子的肉质外种皮,置于室内稍做晾干。将洗净的种子置于湿砂中储藏,m(种子)∶m(湿砂)=1∶1,沙子含水量为5.11%~6.07%,定期翻动种子并检查温度与水分状况。2011年3月将萌动的种子分别播种在大盘山国家级自然保护区的试验地。试验地海拔高度约为500.00 m,土壤类型为山地黄壤。利用遮阳网设置不同光强,分别为0层、1层和2层。测定于2011年6月25-30日(晴天)进行,此时伯乐树处于旺盛生长期。由于山顶试验地伯乐树幼苗长势较差,未予以测量。运用便携式光合作用测量仪(Li-6400)探头分别测量晴天中午11:00时左右的光强,其光强分别约为1 450.00,550.00和200.00 μmol·m-2·s-1。
1.2 测量及计算方法
先随机选取25株长势较为一致的幼苗,测量其地径和高度。由于其枝条尚未完全开展,故未对冠幅等形态指标进行测量。随后选择其中的5株植物进行光合参数的测量,叶片选自下而上第2片复叶完全展开的顶叶作为测量对象。仪器采用美国Licor公司生产的Licor-6400型便携式光合作用测定系统。
光合日变化的测定于6月30日8:00-18:00进行,2 h测量1次。采用净光合速率(Pn),蒸腾速率(Tr),胞间二氧化碳浓度(Ci),大气二氧化碳浓度(Ca),光合有效辐射(RPAR)等的日进程,进而计算水分利用效率(EWUE)=Pn/Tr[15]等。
光响应曲线的测定于6月26-27日进行。设定温度为28.00℃,二氧化碳摩尔分数为400 μmol·mol-1,空气相对湿度为50%~70%,应用Li-6400-02B红蓝光光源提供不同的光合有效辐射强度(RPARμmol·m-2· s-1), 分别在 RPAR为 1 500.00, 1 200.00, 1 000.00, 800.00, 600.00, 400.00, 200.00, 150.00,100.00,50.00,20.00,0.00 μmol·m-2·s-1下测定不同含水量下伯乐树叶片的净光合速率(μmol·m-2·s-1)。
二氧化碳响应曲线的测定于6月28-29日进行。设定温度为28.00℃,有效辐射强度为1 000.00 μmol·m-2·s-1作为测定光强,采用Li-6400液化二氧化碳钢瓶提供不同的二氧化碳体积分数,分别在二氧化碳摩尔分数为400.00,300.00,200.00,150.00,100.00,50.00,20.00,400.00,600.00,800.00,1 000.00,1 200.00 μmol·mol-1的条件下测定叶片净光合速率,拟合得出最大光合速率(Amax),表观羧化速率(ECE),光下呼吸速率(R),二氧化碳补偿点(PCCP)和二氧化碳饱和点(PCSP)。
1.3 数据分析及作图
运用模型对光响应曲线和二氧化碳响应曲线进行拟合[16-17]。运用SPSS 12.0进行显著性检验,运用Excel 2010做图。
2 结果与分析
2.1 光合参数的日变化
对伯乐树1年生幼苗的叶片光合参数日变化进行动态观测结果表明:不同光强下的伯乐树叶片气孔导度(Gs),蒸腾速率(Tr)和光合速率(Pn)均以全光处理下的较高,遮光处理下较低(图1),且遮光处理整体上无显著差异;不同光强下的伯乐树叶片水分利用效率(EWUE)整体表现为先升后降的趋势,其中较高光强下(1 450.00和550.00 μmol·m-2·s-1)的水分利用效率在14:00时有所下降,推测是高温和强光造成了叶片的 “午休”现象。
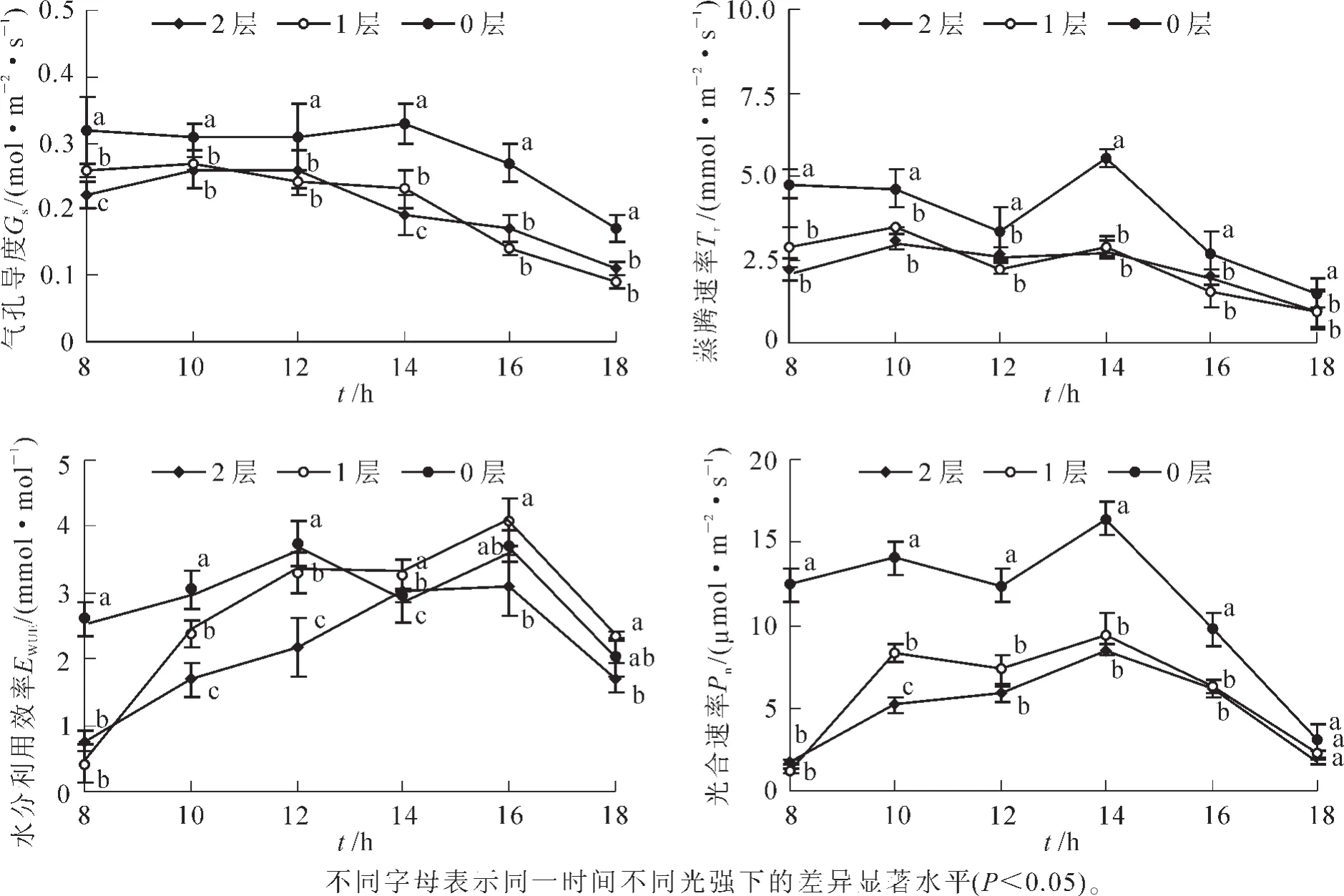
图1 不同光强下伯乐树幼苗气孔导度(Gs),蒸腾速率(Tr),水分利用效率(EWUE)和光合速率(Pn)等的日变化Figure 1 Day changes of Gs,Tr,EWUEand Pnof Bretschneidera sinensis seedlings under different light intensities
2.2 伯乐树幼苗叶片的光响应曲线
伯乐树幼苗叶片光合速率对光的响应曲线见图2。由图2可以看出:当光强较弱时,不同光强下的伯乐树幼苗光合速率差异不明显;当光强大于200.00 μmol·m-2·s-1时,较高光强下(1 450.00和550.00 μmol·m-2·s-1)的光合速率显著高于弱光(P<0.05)。对光响应曲线进行拟合得出的最大光合速率(Pmax)也显示,全光和1层处理下的最大光合速率分别为13.94和12.00 μmol·m-2·s-1,而2层处理下的最大光合速率为4.14 μmol·m-2·s-1,其差异达到显著水平(P<0.05)。从伯乐树幼苗叶片的光补偿点(PLCP)来看,不同光强处理下的光补偿点分别为21.22,19.93和14.95 μmol·m-2·s-1,其值均较低(表1)。

表1 不同光强下伯乐树幼苗光响应参数的比较Table 1 Effects of light intensities on the photoresponse parameters of Bretschneidera sinensis seedlings
2.3 伯乐树幼苗叶片的二氧化碳响应曲线
伯乐树幼苗叶片光合速率对二氧化碳的响应曲线见图3。由图3可以看出:随着二氧化碳摩尔分数的升高,光合速率逐渐升高,其光合速率以全光下最高,1层遮阳网下次之,2层遮阳网下的光合速率最低。对二氧化碳响应曲线拟合的结果表明,不同光强下的最大羧化速率(Amax)分别为39.74,31.82和23.17 μmol·m-2·s-1(表2)。经检验,不同处理之间的差异均达到显著水平(P<0.05)。
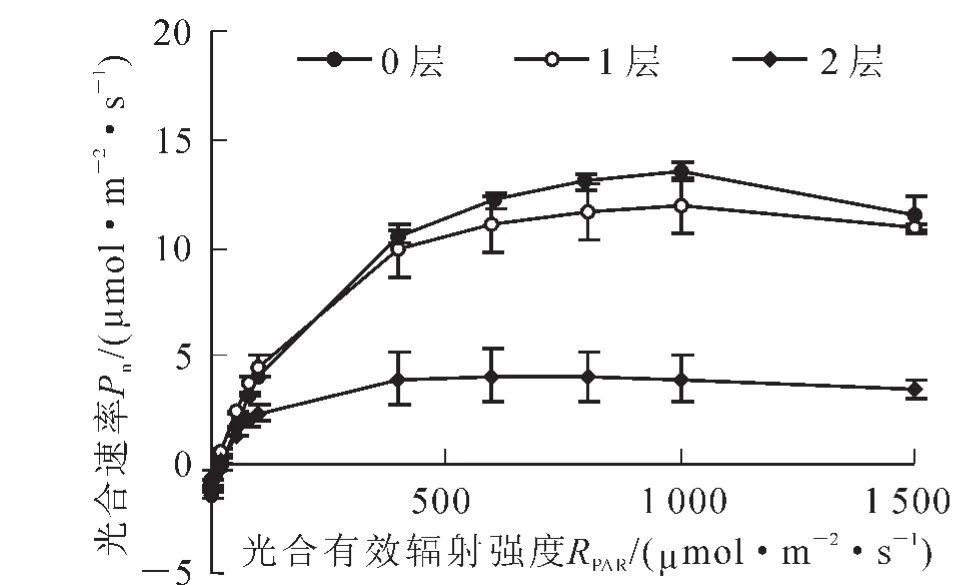
图2 不同光强下伯乐树幼苗叶片光响应曲线Figure 2 Photoresponse curve of Bretschneidera sinensis seedlings under different light intensities
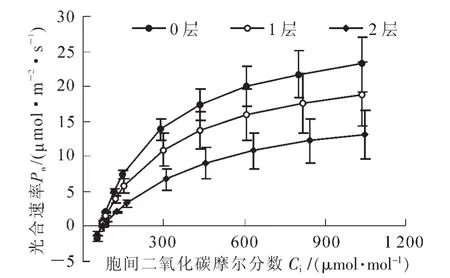
图3 不同光强下伯乐树幼苗叶片二氧化碳响应曲线Figure 3 CO2-response curve of B.sinensis seedlings under different light intensities
2.4 伯乐树幼苗形态指标
由图4可以看出:全光下的伯乐树幼苗地径最大(P<0.05),而1层和2层遮阳网处理之间无显著差异;同样,伯乐树幼苗高度的结果表明,全光下的伯乐树幼苗最高。
3 讨论
植物叶片光合作用对光的响应曲线可深入了解植物的最大光合速率、光饱和点、光补偿点等光合参数,是研究光合作用特性的最主要内容之一[18]。光饱和点的高低反映了光合机构暗反应过程对同化力最大需求量的多少,以及植物利用光强的能力;光补偿点的高低反映了对弱光的利用能力高低[19]。本研究结果表明:高光强下的伯乐树幼苗叶片光饱和点较高,其对应的光合速率日变化及最大光合速率均较高,说明相对高的光强更有利于伯乐树幼苗的光合作用。这与伯乐树幼苗实际生长情况相一致。野外调查也表明伯乐树为高大的乔木树种,在原生分布点多处于群落上层,说明伯乐树为喜光树种。然而,本研究结果与乔琦等[8]研究结果不一致。分析认为有可能是试验地气候及测量时间的不同而造成。本研究地位于浙江省大盘山国家级自然保护区内海拔约500.00 m的山坡。该区温度适宜,雨水充沛,试验期间最大光强为1 450.00 μmol·m-2·s-1。另外,不同光强处理下的伯乐树幼苗光补偿点均较低,说明伯乐树幼苗较耐荫。但弱光下伯乐树幼苗植物相对矮小,长势较慢。这与乔琦等[8]研究结果相一致。
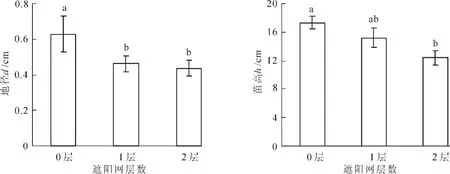
图4 不同光强下伯乐树幼苗地径和高度Figure 4 Ground diameters and height of Bretschneidera sinensi seedlings under different light intensities
光合作用是绿色植物生物量积累的基础。植物光合作用所需要的碳源主要是空气中的二氧化碳。由二氧化碳响应曲线的拟合结果来看,全光下的伯乐树幼苗二氧化碳羧化效率最高,说明其核酮糖-1,5-二磷酸羧化酶/加氧酶的量及酶活性均较高[19]。这与最大羧化速率值相一致,说明较高光强更有利于伯乐树进行二氧化碳同化,从而积累较多有机物以增加其生物量。这与伯乐树幼苗形态指标结果相一致。
另外,本研究结果显示:光合速率的日变化反映出伯乐树幼苗叶片有 “午休”现象,说明强光高温不利于伯乐树幼苗生长。同样,伍铭凯等[20]研究也认为伯乐树不耐高温,易遭日灼。因此,推测本研究与乔琦等[8]研究结果不一致的原因也有可能是温度不同造成的。乔琦等[8]研究在华南植物园的温室内进行。夏季温室温度远高于山地林地;同时,也认为气温和土温过高、土壤湿度较低等也易使部分幼苗遭受干旱胁迫而大量死亡。因此,建议探讨伯乐树的幼苗光响应特性应考虑温度因素。
综合气体交换参数的日变化、光响应曲线和二氧化碳响应曲线的试验结果表明,光强相对较高下的伯乐树幼苗叶片的表观光合速率、最大光合速率、最大羧化速率和二氧化碳羧化效率等光合参数值均较高,幼苗表现出良好的生长态势。建议在进行伯乐树幼苗人工繁育时,不宜过度遮光;在原生地抚育时,应适当增加林窗的光照强度。
[1] 国家林业局,农业部.国家重点保护野生植物名录:第1批[J].中华人民共和国国务院公报,2000(13):39-47.
State Forestry Administration,Ministry of Agriculture of the People’s Republic of China.List of wild plants under state protection group:first[J].Gazet State Counc P R China,2000(13):39-47.
[2] 唐邦权,杨怡.伯乐树实生苗播种育苗技术[J].特种经济动植物,2012(11):31-32.
TANG Bangquan,YANG Yi.Planting seedling technology of Bretschneidera sinensis seedling[J].Spec Econ Anim Plant,2012(11):31-32.
[3] 乔琦,秦新生,郑希龙,等.伯乐树1年生幼苗的光响应特征[J].福建林业科技,2013,40(1):63-70.
QIAO Qi,QIN Xinsheng,ZHENG Xilong,et al.Study on the light response characteristics of 1-year-old seedlings of Bretschneidera sinensis[J].J Fujian For Sci Technol,2013,40(1):63-70.
[4] 张纪卯.伯乐树人工林的生长规律[J].亚热带农业研究,2011,7(1):22-25.
ZHANG Jimao.Study on the growth of Bretschneidera sinensis Hemsl.plantation[J].Subtropi Agric Res,2011,7(1):22-25.
[5] 郭祥泉,周立华,熊自华,等.孑遗树种钟萼木幼树生长特性探讨[J].亚热带植物科学,2012,41(2):32-36.
GUO Xiangquan,ZHOU Lihua,XION Zihua,et al.Study on the growth characteristic of young trees of Bretschneidera sinensis[J].Subtrop Plant Sci,2012,41(2):32-36.
[6] 张季,田华林,王玉奇,等.3个不同地理种源的伯乐树种子和苗期生长差异比较[J].热带农业科学,2011,31(5):12-15.
ZHANG Ji,TIAN Hualin,WANG Yuqi,et al.Comparison of seeds and seedling growth for 3 different geographical provenances of Bretschneidera sinensis[J].Chin J Tropic Agric,2011,31(5):12-15.
[7] 马冬雪,刘仁林.伯乐树幼苗特性与不同年龄阶段苗木生长规律研究[J].安徽农业科学,2012,40(17):9344-9346.
MA Dongxue,LIU Renlin.A study on characteristics and growth laws of Bretschneidera sinensis Hemsi.during young seedling period[J].J Anhui Agric Sci,2012,40(17):9344-9346.
[8] 乔琦,秦新生,邢福武,等.珍稀植物伯乐树一年生更新幼苗的死亡原因和保育策略[J].生态学报,2011,31(16): 4709-4716.
QIAO Qi,QIN Xinsheng,XING Fuwu,et al.Death causes and conservation strategies of the annual regenerated seedlings of rare plant,Bretschneidera sinensis[J].Acta Ecol Sin,2011,31(16):4709-4716.
[9] LUSK C H.Leaf area accumulation helps juvenile evergreen trees tolerate shade in a temperate rainforest[J].Oecologia,2002,132(2):188-196.
[10] 张文辉,祖元刚,刘国彬.10种濒危植物种群的生态学特征及致危因素分析[J].生态学报,2002,22(9):1512-1520.
ZHANG Wenhui,ZU Yuangang,LIU Guobin.Population ecological characteristics and analysis on endangered cause of ten endangered plant species[J].Acta Ecol Sin,2002,22(9):1512-1520.
[11] LIU Peng,YANG Y S,XU Gendi,et al.Physiological response of rare and endangered seven-son-flower(Heptacodium miconioides)to light stress under habitat fragmentation[J].Environ Exp Bot,2006,57(1/2):32-40.
[12] 张旺锋,樊大勇,谢宗强,等.濒危植物银杉幼树对生长光强的季节性光合响应[J].生物多样性,2005,13(5):387-397.
ZHANG Wangfeng,FAN Dayong,XIE Zongqiang,et al.The seasonal photosynthetic responses of seedlings of the endangered plant Cathaya argyrophylla to different growth light environments[J].Chin Biodiv,2005,13(5):387-397.
[13] 刘军.国家Ⅱ级重点保护植物香果树的保护与利用[J].甘肃科技,2003(19):151-152.
LIU Jun.Protection and utilization of national protected plants(Ⅱ)Emmenopterys henry[J].Gansu Sci Technol, 2003(19):151-152.
[14] 谢玉芳,潘林,杨玉芳,等.香果树育苗技术[J].江苏林业科技,2004,31(2):39-40.
XIE Yufang,PAN Lin,YANG Yufang,et al.Breeding technology of Emmenopterys henryi Oliv.[J].J Jiangsu For Sci Technol,2004,31(2):39-40.
[15] NIJS I,FERRIS R,BLUM H.Stomatal regulation in a changing climate:a field study using free air temperature increase(FATL)and free air CO2enrichment(FACE)[J].Plant Cell Environ,1997,20(8):1041-1050.
[16] YE Zipiao.A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J].Photosynthetica,2007,45(4):637-640.
[17] YE Zipiao,YU Qiang.A coupled model of stomatal conductance and photosynthesis for winter wheat[J].Photosynthetica,2008,46(4):637-640.
[18] 叶子飘,于强.冬小麦旗叶光合速率对光强度和CO2浓度的响应[J].扬州大学学报:农业与生命科学版,2008,29(3):33-37.
YE Zipiao,YU Qiang.Photosynthetic response to irradiance and CO2concentration for flag leaves of winter wheat[J].J Yangzhou Univ Agric Life Sci Ed,2008,29(3):33-37.
[19] 叶子飘.光合作用对光和CO2响应模型的研究进展[J].植物生态学报,2010,34(6):727-740.
YE Zipiao.A review on modeling of responses of photosynthesis to light and CO2[J].Chin J Plant Ecol,2010,34(6):727-740.
[20] 伍铭凯,杨汉远,龙舞,等.伯乐树种子育苗试验[J].贵州林业科技,2006,34(4):39-42.
WU Mingkai,YANG Hanyuan,LONG Wu,et al.A study on the seedling cultivation of Bretschneidera sinensis Hemsl.[J].J Guizhou For Sci Technol,2006,34(4):39-42.
Photosynthetic physiological response of Bretschneidera sinensis seedlings to light intensity
YE Feiying1,CHEN Zilin2,ZHENG Weicheng1,LIU Julian1,ZHOU Yuhong2, ZHANG Jianyu3,PAN Chengchun1,JI Guohua1
(1.Management Office,Jiulong Mountain National Nature Reserve,Suichang 323300,Zhejiang,China;2. Management Office,Dapanshan National Nature Reserve,Pan’an 322300,Zhejiang,China;3.Wenzhou Vocational College of Science and Technology,Wenzhou 325006,Zhejiang,China)
Bretschneidera sinensis,an endemic species in China and one China’s national first-level protected wild plants,is a particularly rare,severely endangered,and protected plant whose conservation for biodiversity is needed.This study determined the effect of different light intensities on the photosynthetic physiological response of B.sinensis and offered some suggestions for conservation.The photosynthetic characteristics of B. sinensis seedlings grown outdoors were studied with different light intensities(1 450,550,and 200 μmol·m-2· s-1).Results showed that daily changes in stomatal conductance,transpiration rate,and photosynthetic rate with 1 450 μmol·m-2·s-1were higher than with 550 and 200 μmol·m-2·s-1.As light intensity decreased,the maximum photosynthetic rate(Pmax),the light saturation point(LSP),the carboxylation rate(CU),and the maximum rate of carboxylation (Amax)all decreased.Therefore,light intensity should be considered carefully when biodiversity conservation of B.sinensis to be carried out.[Ch,4 fig.2 tab.20 ref.]
plant physiology;Bretschneidera sinensis;seedling;light intensity;photosynthetic physiological
S718.3;Q948.1
A
2095-0756(2015)05-0716-06
10.11833/j.issn.2095-0756.2015.05.009
2014-09-14;
2015-04-04
林业国家级自然保护区中央财政补助资金资助项目(浙财农字[2010]174号);浙江省温州市科技计划项目(S20130003)
叶飞英,助理工程师,从事自然资源保护研究。E-mail:315126864@qq.com。通信作者:郑伟成,从事自然资源保护与研究。E-mail:jlszwc@126.com
