不同坡向条件下小叶杨光合光响应曲线研究
2015-01-03王富刚徐伟洲亢福仁尚爱军
王富刚,徐伟洲,2,亢福仁,尚爱军,张 静
(1.榆林学院 生命科学学院,陕西 榆林719000;2.西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌712100)
半干旱黄丘陵区是黄土高原植被破坏最为严重的地区,人工林生态系统建设是该地区生态植被恢复与退耕还林的重要方式,其关键技术在于树种的选择,应最终营造以灌木为主,灌、草、乔相结合的水土保持林[1-2]。坡向条件是黄土丘陵区最主要的地形因子,它通过改变光照、温度、水分和土壤等生态因子,进而对植物生长发育、生产力以及生态系统功能等产生重要影响[3]。因此,根据区域不同立地条件对类型树种进行合理布局也显得尤为重要[4]。植物光合作用是植物体内最重要的代谢过程,光响应曲线反映的是净光合速率随光强变化的特性。通过拟合光响应曲线得到的各种生理参数对了解植物光反应过程的光合效率非常重要[5],也是研究植物光合生理生态过程对外界环境条件变化响应的基础[6]。
小叶杨(Populussimonii)具有耐旱、耐寒、耐瘠薄等优良特性,是黄土丘陵地区人工植被恢复常选用的乡土树种,也是该地区主要水土保持树种之一,在退耕还林与植被恢复工程中有较大的开发潜力。目前,关于小叶杨生理生态特征方面的研究取得了一批成果[7-8],但关于黄土丘陵区不同坡向条件下的光合生理生态特征比较研究尚未见报道。随着我国退耕还林草工程的实施,进一步系统研究黄土丘陵区不同坡向生境条件下小叶杨生理与生长的空间差异有利于该区人工植被的合理配置[9]。因此,本研究系统比较了该地区阴坡、半阴坡、半阳坡和阳坡条件下小叶杨光合光响应、蒸腾速率与水分利用效率等光合生理特征,旨在为该地区小叶杨人工林的合理配置和水土保持林的科学营造提供理论依据。
1 材料与方法
1.1 试验区概况
本研究区位于陕西省吴起县境内,属典型的黄土高原梁状丘陵沟壑区,地处东经107°52′57″—108°32′49″,北纬36°33′33″—37°24′27″,海拔1 500~1 650m,年均气温7.0℃,≥10℃积温3 086℃,无霜期146d左右,多年降雨量为478.3mm,且7—9月占60%以上,多年平均陆面蒸发量为1 016.7mm,气候属半干旱温带大陆性季风气候,主要土壤类型为黄绵土,土地类型以梁峁坡地为主,约占总面积的77.1%。
该区处于森林草原向典型草原过渡地带,由于气候、地质地貌、土壤肥力等的制约,以及人为活动的影响,天然植被已破坏殆尽,现存植被均系20世纪70年代的人工营造,主要的造林树种有小叶杨(Populussimonii),河北杨(Ponulusnopeiensis)、山杏(Armeniaca sibirica.)、山桃(Amygdalusdavidiana)、沙棘(Hippo-phaerhamnoides)、柠条(Caraganakorshinskii)、沙 柳(Salixpsammophila)、杠柳(Periplocasepium)等,以灌木为主,小乔木林比例低,且呈片断零星分布。
1.2 试验设计与测定项目
本试验选样地点位于吴起县长城乡杨庄,试验以该地区30a育龄小叶杨人工栽培灌木林为研究对象,试验于2012年在山坡下部分阴坡、半阴坡、半阳坡和阳坡选取人工小叶杨林发育较好的地段分别设置10m×10m标准样地4块,共计设置样地16块,坡度在13o~16o。
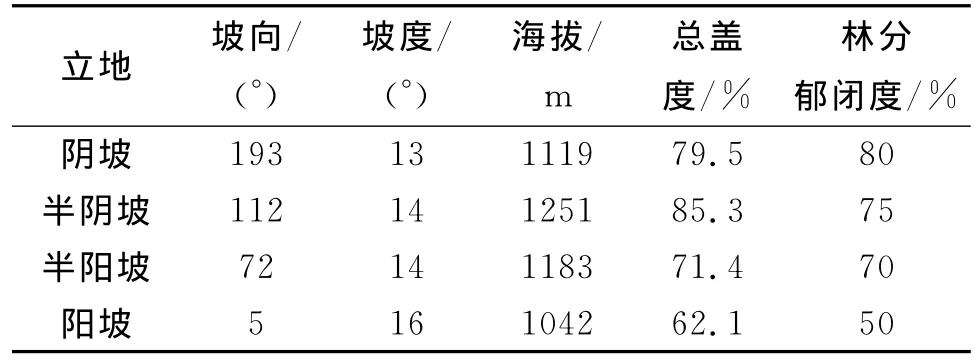
表1 不同立地条件下样地的基本概况
试验选择晴朗天气,分别于5月27—31日,7月21—25日、9月11—15日采用Li-6400便携式光合仪测定小叶杨叶片光合光响应曲线。测定时段为每日9:00—11:30,每次随机从每块标准样地中选择生长状况良好的3株小叶杨完全展开叶进行测定,测定过程中将叶温设置为25oC,叶室内CO2浓度设定为380μmol/mol,用Li-6400自带红蓝光源将光合有效辐射[PAR,μmol/(m2·s)]梯度设置为 2 000,1 500,1 200,1 000,800,600,400,200,100,50,20,0 μmol/(m2·s),改变光强后最少稳定时间为50s,当测定结果变异率小于0.05时由仪器自动记录,可测定的参数有净光合速率[Pn,μmol/(m2·s)]、蒸腾速率[Tr,mmol/(m2·s)]等。叶片水分利用效率[WUE,umol/mmol]按照 WUE=Pn/Tr计算。
1.3 光响应曲线拟合方法
测定所得植物光合数据经过直角双曲线修正模型进 行 拟 合[10],可 获 得 最 大 净 光 合 速 率 [Pmax,μmolCO2/(m2·s)]、暗呼吸速率[Rd,μmolCO2/(m2·s)]、光补偿点[LCP,μmol/(m2·s)]、光饱和点[LSP,μmol/(m2·s)],该模型拟合方程表达式为:
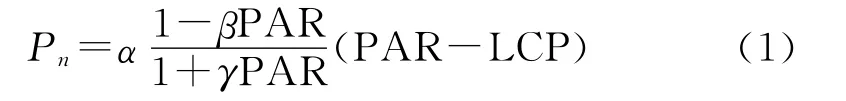
式中:Pn——净 光 合 速 率 [μmol/(m2·s)];α、β、γ——不依赖光强的系数;PAR——光合有效辐射[μmol/(m2·s)],在0~200μmol/(m2·s)范围内通过线性回归求其Pn-PAR曲线初始斜率,即为表观量子效率[AQE,μmolCO2/μmol];LCP——光补偿点[μmol/(m2·s)]。
1.4 数据处理
试验数据采用Excel 2003进行绘图,用SPSS 16.0进行相关统计分析,其中双因素方差分析用于比较测定时期、坡向条件及其二者间可能存在的交互作用,不同处理下光响应参数均值显著性差异采用TukeyHSD进行多重比较(p<0.05)。
2 结果与分析
2.1 光合光响应实测曲线比较
2.1.1 净光合速率 由图1可看出,不同坡向条件小叶杨叶片Pn均随着PAR的增强而升高,但其光响应曲线变化规律在不同测定时期存在明显差异。5月下旬,在PAR小于50μmol/(m2·s)条件下,小叶杨的光响应曲线变化规律在4种坡向间均无显著性差异,而在PAR大于50μmol/(m2·s)条件下出现显著性差异(p<0.05);7月下旬和9月中旬,当PAR大于100μmol/(m2·s)时,不同坡向条件下小叶杨的光合光响应曲线变化规律才出现显著性差异(p<0.05)。
5月下旬,不同坡向下的光响应曲线均存在光饱和现象,而其在7月下旬和9月中旬时均无明显的光饱和。就不同测定时期饱和峰值而言,阴坡、半阴坡与半阳坡条件下的饱和峰值均显著高于阳坡条件(p<0.05),且前三者间无显著性差异(图1)。
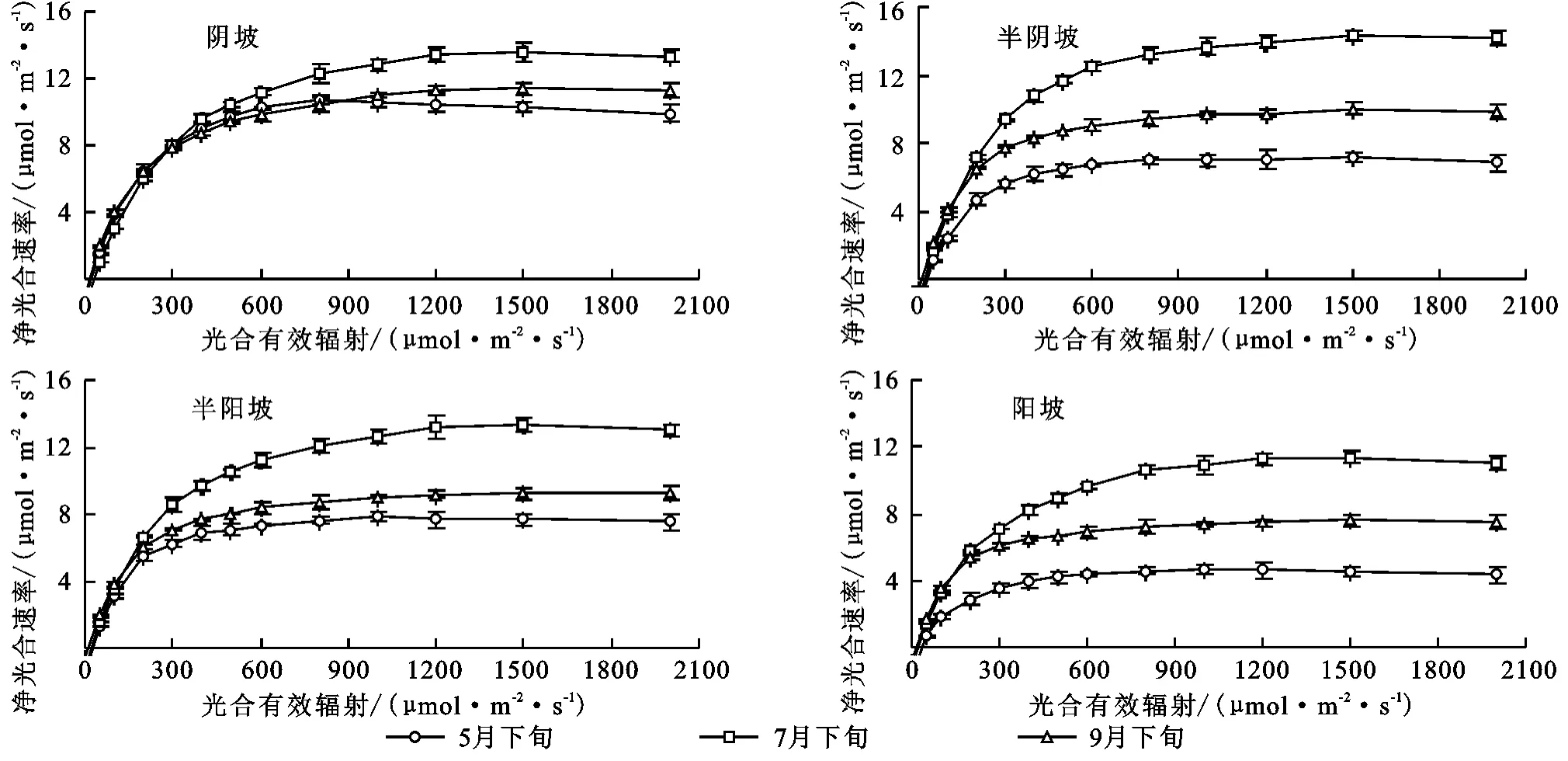
图1 不同坡向条件下小叶杨叶片净光合速率
2.1.2 蒸腾速率 不同测定时期时,4种坡向条件下小叶杨的Tr均随着PAR的增强而升高,但5月下旬和9月中旬时的升高幅度明显小于7月下旬。5月下旬,半阴坡和半阳坡条件下的Tr均显著高于阴坡和阳坡条件(p<0.05),且而两者间不显著性差异;7月下旬,阴坡条件下的Tr均显著高于其他3种坡向条件(p<0.05),而半阴坡、半阳坡和阳坡均在高光强(PAR大于1 400μmol/(m2·s))下才出现明显差异;9月中旬,以半阴坡条件下小叶杨的Tr显著最低(p<0.05),且其余3种坡向条件间无显著性差异(p<0.05)(图2)。
2.1.3 水分利用效率 就不同测定时期而言,随着光合有效辐射的增强,不同坡向条件下小叶杨的WUE均呈先急剧上升而后缓慢降低的变化趋势。5月下旬,以阴坡条件下的WUE显著高于阳坡、半阳坡和半阴坡(p<0.05),且后3者间无显著性差异;7月下旬,以半阴坡条件下小叶杨的WUE显著高于其余3种坡向条件(p<0.05);9月中旬,阴坡和半阴坡条件下的叶片WUE均明显高于阳坡与半阳坡条件(p<0.05)(图3)。
2.2 曲线拟合结果比较
2.2.1 最大净光合速率 由表2得出,不同坡向条件与测定时期下拟合方程的决策系数均在0.99以上,说明直角双曲线修正模型可以较准确的拟合小叶杨叶片光合光响应变化规律(表2)。阴坡条件下,5月下旬和9月中旬的小叶杨叶片Pmax显著低于7月下旬(p<0.05);而阳坡、半阴坡和半阳坡条件下,不同测定时期的Pmax显著顺序均为7月下旬>5月下旬>9月中旬(p<0.05)(表2)。
就不同测定时期而言,5月下旬和9月中旬时,小叶杨叶片Pmax以阴坡条件下明显大于半阴坡和半阳坡,且阳坡条件下显著最低(p<0.05);7月下旬,阴坡、半阴坡和半阳坡的Pmax显著高于阳坡,且前3者坡向间无显著性差异(表2)。坡向条件、测定时期及其二者间的交互作用均对小叶杨的Pmax产生显著影响(表3)。

图2 不同坡向条件下小叶杨叶片蒸腾速率

图3 不同坡向条件下小叶杨叶片水分利用效率
2.2.2 表观量子效率 阴坡条件下,小叶杨的AQE以5月下旬和7月下旬显著低于9月中旬(p<0.05),且前两者间无显著性差异;半阴坡和半阳坡条件下的AQE在3个测定时期间无显著性差异;阳坡条件下,以5月下旬和9月中旬的AQE显著高于7月下旬(p<0.05)(表2)。在5月下旬,阴坡和半阴坡条件下的小叶杨AQE均显著低于半阳坡和阳坡(p<0.05),且前两者或后两者间无显著性差异;7月下旬,半阳坡条件下的AQE显著高于阳坡条件(p<0.05),且均与阴坡与和阴坡条件无显著性差异;9月中旬,阴坡条件下的AQE显著高于其余3种坡向条件(p<0.05)(表2)。坡向条件、测定时期及其二者间的交互作用均对小叶杨的AQE具有显著影响(表3)。
2.2.3 暗呼吸速率 阴坡条件下,不同测定时期的Rd的显著顺序为7月下旬>5月下旬>9月中旬(p<0.05);而半阴坡、半阳坡和阳坡条件下的Rd的显著性顺序均为5月下旬>7月下旬>9月中旬(p<0.05);就不同坡向条件而言,5月下旬时以阴坡显著低于半阴坡、半阳坡和阳坡条件,而7月下旬的变化趋势恰好与其相反(p<0.05);9月中旬,阴坡与阳坡条件下的Rd显著高于半阴坡和半阳坡条件(p<0.05)(表2)。交互分析表明,除坡向条件对小叶杨Rd无显著性影响外,测定时期及其与坡向条件的交互作用均对其有显著性影响(表3)。
2.2.4 光饱和点 在4种坡向条件下,不同测定时期小叶杨的LSP均表现出相同的变化规律,以5月下旬显著低于7月下旬和9月中旬(p<0.05),且后两者测定时期间无显著性差异(表2);5月下旬和9月中旬时,阴坡、半阴坡、半阳坡和阳坡条件下的LSP均无显著性差异;7月下旬,半阳坡的LSP显著高于阴坡(p<0.05),且均与其余两种坡向条件间无显著性差异(表2)。坡向条件、测定时期及其二者间的交互作用均对小叶杨的LSP具有显著影响(表3)。
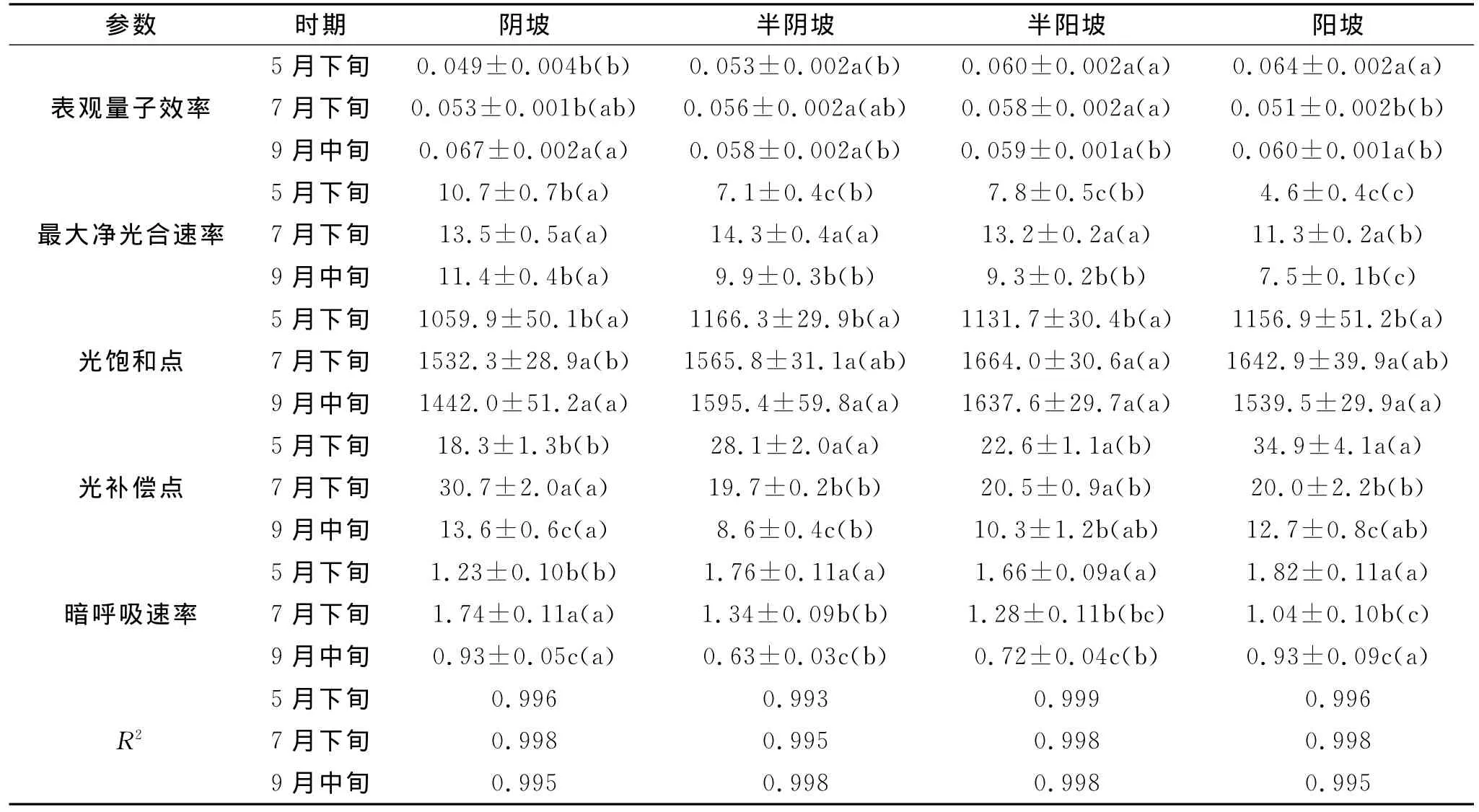
表2 不同坡向条件下小叶杨光合光响应曲线拟合结果

表3 不同坡向、时期及其交互作用对叶片光合光响应曲线各参数的影响
2.2.5 光补偿点 阴坡条件下,不同测定时期小叶杨的LCP的显著性顺序为7月下旬>5月下旬>9月中旬(p<0.05);半阴坡和阳坡条件下,其不同测定时期的显著顺序为5月下旬>7月下旬>9月中旬(p<0.05);半阳坡条件下,以5月下旬和7月下旬的LCP显著高于9月中旬(p<0.05),且前两者间无显著性差异(表2)。坡向条件、测定时期及其二者间的交互作用均对小叶杨的LCP具有显著影响(表3)。
3 讨 论
立地条件中坡向的差异会导致太阳辐射强度、温度、土壤湿度、雨量等生态因子的变化,从而对植物个体生理生长过程产生影响,进而引起植物群落生态系统结构发生变化[11]。植物的光合速率和蒸腾速率用来表征不同植物的重要生理指标。本研究中,不同坡向条件下小叶杨的Pmax和Tr均以7月下旬显著最高,表明小叶杨在7月下旬处于生长旺盛期,这也有可能归因于该地区降雨多集中在7—9月份引起土壤水分条件相对较为充分。
在适宜的环境条件下,通常用叶片的Pmax来表示叶片的潜在光合能力,此值越高说明植物在强光下越不易产生光抑制反应[12]。本研究中,不同坡向条件下的小叶杨叶片光响应曲线均没有出现明显的光饱和现象,表明该灌木树种适宜于种植在阳光相对充分的立地环境条件中;另一方面,不同坡向条件下小叶杨的光合能力在3种测定时期均以阳坡条件下显著最低,且在5月下旬和9月中旬以阴坡条件下显著最高,这表明小叶杨适宜于种植在阴坡、半阴坡或半阳坡条件,这与前人在黄土高原丘陵沟壑区对不同生境条件下小叶杨人工林分布调查状况一致[13]。
植物叶片AQE反映了其对光能的利用情况,尤其是对弱光的利用能力,这有助于明确植物光合机构是否运转正常,水分胁迫通常会造成植物的AQE降低[14]。本研究中,5月下旬,阳坡和半阳坡条件下小叶杨对弱光的利用能力显著高于阴坡和半阴坡条件(p<0.05);7月下旬和9月中旬阳坡条件下其叶片对弱光的利用效率显著低于其他3种坡向条件(p<0.05),这可能归因于在土壤水分相对较为充分的7—9月份,阴坡、半阴坡和半阳坡由于相对较高的土壤储水量增加了其对弱光的利用能力,进而保证植物维持较高的量子产额[7]。
叶片WUE指植物消耗单位水量生产出的同化量,是评价植物对环境适应的综合生理生态指标,也是确定植物体生长发育所需水分供应的重要指标之一[15]。本研究中,在5月和7下旬,分别以阴坡和半阴坡条件下的叶片 WUE显著最高(p<0.05);而9月中旬时,阴坡和半阴坡条件下的叶片WUE均明显高于阳坡和半阳坡条件(p<0.05),表明小叶杨在阴坡和半阴坡条件下不仅保持了相对较强的叶片光合能力,而且均有较高的水分利用能力。
植物的LCP和LSP分别体现了其对弱光和强光的适应能力,能够反映其对于光照条件需求范围[16]。本研究中,不同坡向条件下小叶杨的的LCP范围为8.6~30.7μmol/(m2·s),这基本与典型阳生植物的9~27μmol/(m2·s)相符,LSP范围为1059.9~16664.0μmol/(m2·s),高于典型阳性植物的360~900μmol/(m2·s),表明小叶杨光能利用范围较广且净光合作用有效时间较长,属于典型阳生灌木树种,这表明小叶杨具有良好的光适应能力,这与前人对该树种在不同坡位条件的研究结果一致[13]。
4 结 论
(1)非直角双曲线模型拟合方程决策系数R2均在0.99以上,表明该模型能很好的拟合不同坡向条件下小叶杨的光合光响应曲线。
(2)不同坡向条件下小叶杨的光合光响应曲线均无明显的光饱和现象,表明在其生育期均没有出现光抑制现象,说明小叶杨适宜种植在阳光和水分相对较为充分的立地环境。
(3)阴坡条件下,5月下旬和9月中旬的小叶杨叶片Pmax显著低于7月下旬;而阳坡、半阴坡和半阳坡条件下,不同测定时期的Pmax显著顺序均为7月下旬>5月下旬>9月中旬。
(4)综合比较分析不同坡向条件下叶片光合能力、蒸腾速率和水分利用效率等生理生态特征,可以推测出就坡向条件而言,其在黄土丘陵区最佳种植顺序为:阴坡>半阴坡>半阳坡>阳坡。
[1] 张淑勇,周泽福,张光灿,等.半干旱黄土丘陵区4种天然次生灌木光合生理和水分利用特征[J].林业科学,2008,44(12):140-146.
[2] 万雪琴,胡庭兴,张健,等.坡耕地退耕还林后的植被恢复[J].林业科学,2005,41(2):191-194.
[3] 聂莹莹,李新娥,王刚.阳坡—阴坡生境梯度上植物群落α多样性与β多样性的变化模式及与环境因子的关系[J].兰州大学学报:自然科学版,2010,46(3):73-79.
[4] 李俊辉,李秧秧.立地条件和树龄对刺槐叶形态及生理特性的影响[J].水土保持研究,2012,19(4):176-181.
[5] 徐伟洲,徐炳成,段东平,等.不同水肥条件下白羊草光合生理生态特征研究.光响应曲线[J].草地学报,2010,18(6):773-779.
[6] 张中峰,黄玉清,莫凌,等.岩溶植物光合—光响应曲线的两种拟合模型比较[J].武汉植物学研究,2009,27(3):340-344.
[7] 时慧君,杜峰,张兴昌.毛乌素沙地几种主要植物的光合特性[J].西北林学院学报,2010,25(4):29-34.
[8] 张颖,朱铭强,李浩,等.半干旱黄土丘陵区4种灌木树种的光合特性研究[J].西北农林科技大学学报:自然科学版,2012,40(3):65-70.
[9] 温仲明,从怀军,焦峰,等.黄土丘陵沟壑区小叶杨林生长的空间差异分析:以吴旗县为例[J].水土保持通报,2005,25(1):15-17.
[10] 叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356-1361.
[11] 祝顺波,许文年,刘刚,等.坡度对紫花苜蓿和狗牙根光合生理特性的影响[J].水土保持研究,2012,19(2):218-221.
[12] Farquhar G D,von Caemmerer S,Berry J A.A biochemical model of photosynthetic CO2assimilation in leaves of C3species[J].Planta,1980,149(1):78-90.
[13] 张婷,张文辉,郭连金,等.黄土高原丘陵区不同生境小叶杨人工林物种多样性及其群落稳定性分析[J].西北植物学报,2007,27(2):340-347.
[14] Olsson T,Leverenz J W.Non-uniform stomatal closure and the apparent convexity of the photosynthetic photon flux density response curve[J].Plant,Cell &Environment,1994,17(6):701-710.
[15] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
[16] 李扬,黄建辉.库布齐沙漠中甘草对不同水分和养分供应的光合生理响应[J].植物生态学报,2009,33(6):1112-1124.
