华山松大小蠹对非寄主挥发物的触角电位和行为反应
2015-01-02张维平陈志辉谢寿安吕淑杰刘绥鹏
张维平,陈志辉,谢寿安*,吕淑杰,刘绥鹏,黎 华
(1.西北农林科技大学 林学院,陕西 杨陵712100;2.丹凤县野生动物保护站,陕西 丹凤726200;3.国营北川羌族自治县林场,四川 北川622750)
华山松大小蠹(Dendroctonus armandi Tsai et Li)属鞘翅目(Coleoptera)、齿小蠹科(Ipidae)、大小蠹属(Dendroctonus)昆虫,又名凝脂小蠹,为我国特有种,主要分布于陕西、四川、湖北、甘肃、河南。该虫主要危害30a以上的华山松健康立木,入侵后引起树势急剧衰弱,为其他小蠹及蛀干害虫入侵奠定了基础,故属于秦岭华山松的先锋虫种,是造成我国陕西秦岭林区、大巴山南北坡华山松大量枯死的主要原因[1]。
小蠹虫在与寄主植物长期协同进化过程中形成了一定的相互关系,许多小蠹虫均具有特定的取食习性,仅攻击一种或几种密切相关的寄主树木[2-3],这些寄主树木广泛分散在混交林中,时空上分布不均衡[4],因此,寻找到正确的生境、寄主树木并定位适合侵染的寄主个体是小蠹虫扩散传播的首要环节[5]。
小蠹虫能够识别寄主树木释放的挥发性物质,作为寻找寄主的嗅觉信号[2]。但与适宜的寄主树木相比,小蠹虫在寻找寄主的过程中会遇到更多的不适宜的寄主和非寄主树木。对树木的驱避可能是根据寄主某些特性的失调和对非寄主刺激物如非寄主挥发物的消极反应而确定的[6-7]。在远处就能识别和回避不适宜的寄主和非寄主,对植食性小蠹减少被捕食和能量损失的风险会是有益的[8-9]。
针叶树小蠹虫的寄主和非寄主释放挥发性次生化学物质通常是不同的,对于针叶树,挥发物主要是单萜,而非寄主被子植物相对释放大量的六碳的醇、醛和酯类化合物,称为“绿叶气体”(green leaf volatiles,GLVs)[10]。张 庆 贺[10-11]等 和 T.M.Poland[12]等发现非寄主挥发物中的1-己醇、顺-3-己烯-1-醇、3-辛醇和1-辛烯-3-醇可以抑制重齿小蠹(Ips duplicates)和 纵 坑 切 梢 小 蠹 (Tomicus piniperda)的聚集,顺-3-己烯-1-醇、反-2-己烯-1-醇和1-己醇能明显降低二齿星小蠹(Pityogenes bidentatus)对其聚集信息素顺式-马鞭草烯醇和诱杀烯醇的聚集反应。目前非寄主挥发物已经在云杉八齿小蠹(Ips typographus)、山松大小蠹(D.ponderosae)、南部松小蠹(D.frontalis)等小蠹虫的生态防治中得到了广泛应用[13-15],而有关华山松大小蠹对非寄主挥发物反应的研究尚未见报道。
为探明非寄主挥发物在华山松大小蠹寄主选择过程中的作用,研究了华山松大小蠹对其非寄主挥发物的8种标准化合物的反应,筛选其中具有明显反应的化合物,为研制和开发驱避剂型,开展华山松大小蠹的综合治理以及生态调控奠定基础。
1 材料与方法
1.1 供试虫源
2014年6月在秦岭火地塘华山松林内(海拔1 915m,33°27'10.0″N,108°28′49.2″E)伐取被华山松大小蠹侵害的华山松,带回实验室保存。华山松大小蠹开始羽化后,每日收集单独保存,于4℃的冰箱内储存。
1.2 供试化合物
将各样品配制为液状石蜡溶液,以0.01~100(以10倍递增)进行剂量反应测试。所用样品的名称、纯度及来源见表1。
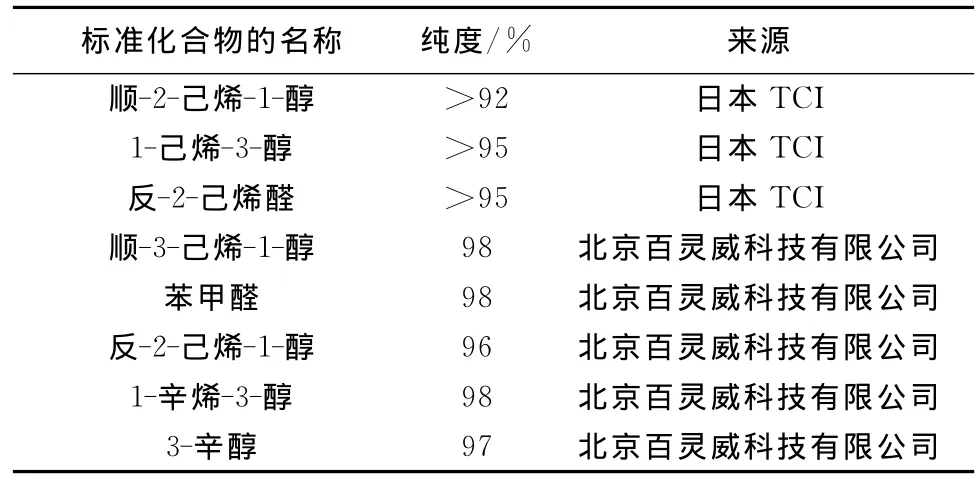
表1 8种供试化合物的名称、纯度及来源Table 1 Name,purity and source of 8standard chemical compounds
1.3 触角电位测定
用解剖刀将华山松大小蠹触角自基部切下,用Spectra 360导电胶将其固定在PR(Gain10×10)电极上,气味管末端与触角之间的距离约1cm。用微量取样器抽取10μL样品溶液,均匀地滴在2cm×0.5cm的滤纸条上,放入10cm长的样品管中,样品管末端连接气体刺激控制装置,其通有持续性湿润气流(40mL·min-1),刺激气流流速为20mL·min-1。待基线稳定后给予刺激,每次刺激时间为0.5s,刺激间隔至少30s,以保证触角感受能够完全恢复,每只待测小蠹虫只用一根触角。试验每处理测试触角8根,每样品平均刺激5次。选用1μg·μL-1的顺-3-己烯-1-醇作为对照,因为顺-3-己烯-1-醇是植物中常见的绿叶气味,在昆虫与植物关系中具有重要作用[16],常被选为昆虫触角电位反应的测试标样[17-19],且已证明对多种小蠹虫具有反应活性[19-20]。相同样品按浓度自低向高进行测定。将每一样品观测值除以前后2次对照测定值的平均值即得该样品的触角电位反应相对值。
1.4 行为反应测定
采用Y型嗅觉仪测定华山松大小蠹成虫对8种非寄主挥发物的行为反应。Y型嗅觉仪主臂长15cm,两侧臂长10cm,侧臂间夹角为75°,主臂和侧臂内径均1.5cm。两侧臂分别用硅胶管与500 mL锥形瓶相连。锥形瓶中放置含有待测样品的2 mL样品瓶,锥形瓶用硅胶管依次与流量计和加湿瓶相连。空气流速调节为0.5~0.6L·min-1;测量时间为8:00-18:00;测量温度为25℃±1℃。测量时将华山松大小蠹由Y型管主臂管口引入,如果试虫进入气味源的支管并爬过分支臂的连接处的距离超过5cm,记作华山松大小蠹对该处理臂做出了选择,即对该处理具有趋向反应,否则记为对该气味无选择。每测5头将Y型管2分支臂对调,每个处理测量完成后更换Y型管和锥形瓶,用酒精彻底清洗,自然晾干。每种样品只测试1、10、100μg·μL-13种较高浓度下的行为反应,每组测试的个体数达30头,重复3次。
1.5 数据统计分析
试验数据采用SPSS 19.0软件进行统计分析,利用Duncan多重分析法比较华山松大小蠹成虫对8种挥发物行为反应和触角电位反应的差异。利用T检验分析比较雌虫和雄虫触角电位反应之间的差异。试验数据用Sigmaplot 12.0软件进行作图。

2 结果与分析
2.1 对不同非寄主挥发物的触角电位反应
从表2可以看出,在100μg·μL-1浓度下,顺-2-己烯-1-醇引起华山松大小蠹雌虫的触角电位反应最强,与其他7种挥发物达到显著性差异水平(p<0.05),顺-3-己烯-1-醇次之,1-辛烯-3-醇引起的触角电位反应最弱。华山松大小蠹雄虫对顺-3-己烯-1-醇的触角电位反应最强,并与其他7种挥发物达到极显著性差异水平(p<0.01),其次为顺-2-己烯-1-醇和苯甲醛,与其余挥发物具有显著性差异,3-辛醇和1-辛烯-3-醇引起的华山松大小蠹雄虫的触角电位反应最弱。T检验表明,华山松大小蠹成虫对8种挥发物(100μg·μL-1)的触角电位反应不存在性别差异。
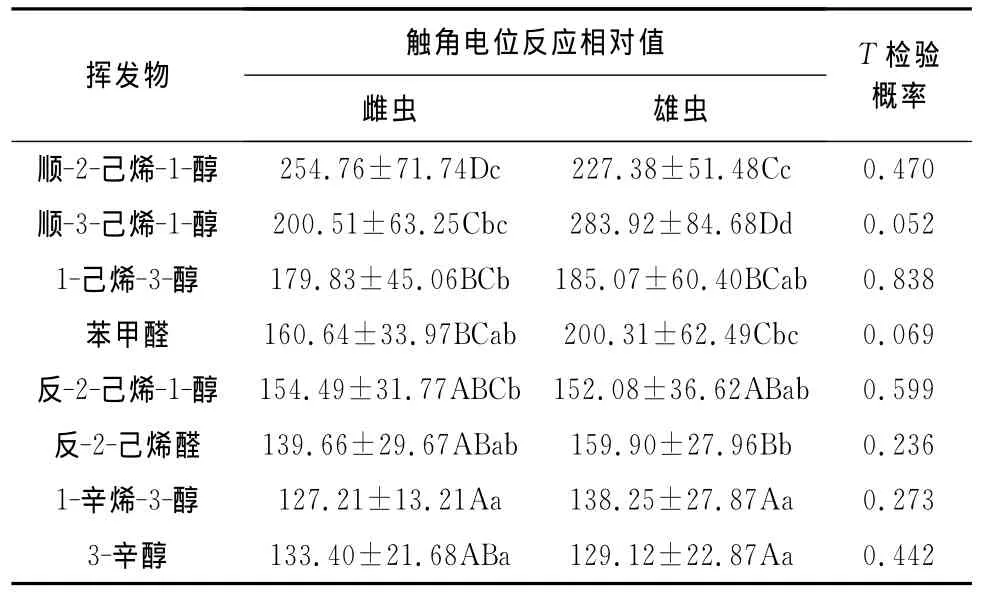
表2 华山松大小蠹成虫对非寄主挥发物的触角电位反应Table 2 Electroantennographic responses of adult D.armandi to nonhost volatiles 100μg·μL-1
2.2 对不同浓度非寄主挥发物的触角电位反应
由图1可知,所选择的8种非寄主挥发物在特定的浓度下均能引起华山松大小蠹雌雄虫一定的触角电位反应。8种非寄主挥发物的剂量反应均表现出相同的规律:随着挥发性化合物浓度的升高,触角电位反应值增大,最大值均出现在100μg·μL-1浓度。在0.01μg·μL-1和0.1μg·μL-1浓度下,反式-2-己烯-1-醇和3-辛醇引起华山松大小蠹雌雄虫的触角电位反应较强;在10μg·μL-1和100μg·μL-1浓度下,雌雄虫对顺-2-己烯-1-醇和顺-3-己烯-1-醇的触角电位反应较强,对1-辛烯-3-醇和3-辛醇的触角电位反应较弱。T检验表明,在0.01、0.1、1、10μg·μL-1浓度下,华山松大小蠹对非寄主挥发物的触角电位反应仍不存在性别差异。
2.3 对8种非寄主挥发物的室内趋向反应
由图2可知,室内趋向反应表明,除顺-3-己烯-1-醇和反-2-己烯-1-醇驱避反应不明显外,华山松大小蠹对其余6种非寄主挥发物均表现出不同程度的驱避作用,随着挥发物浓度的增加,挥发物的驱避作用增强。在1μg·μL-1浓度下,3-辛醇与其他挥发物相比呈现显著性差异(p<0.05),表现出明显的驱避作用,正趋向率低于30%;顺-3-己烯-1-醇、1-己烯-3-醇和1-辛烯-3-醇三者之间没有显著性差异,但与顺-2-己烯-1-醇和反-2-己烯-1-醇具有显著性差异,表现出较弱的驱避作用,正趋向率低于40%。在10μg·μL-1浓度下,顺-3-己烯-1-醇、1-己烯-3-醇、苯甲醛和反-2-己烯醛的驱避作用增强;1-辛烯-3-醇驱避作用明显增强。在100μg·μL-1浓度下,除顺-2-己烯-1-醇和反-2-己烯-1-醇以外,其他6种挥发物均能引起华山松大小蠹成虫明显的驱避反应,正趋向率低于30%,并且挥发物之间没有显著性差异。
3 结论与讨论
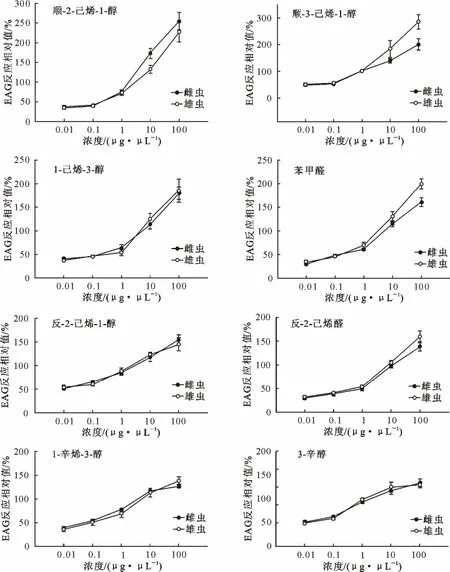
图1 华山松大小蠹雌雄虫对8种非寄主挥发物不同浓度的触角电位反应Fig.1 Electroantennographic responses of D.armandi to different concentrations of eight nonhost volatiles
非寄主挥发物在小蠹虫的寄主选择过程中担当重要的角色,目前已发现十几种小蠹虫对非寄主挥发物具有反应[11]。本研究测定的8种非寄主挥发物在特定浓度下均引起华山松大小蠹的触角电位反应。华山松大小蠹对非寄主挥发物的触角电位反应不具有性别差异,这与之前对小蠹虫非寄主挥发物的研究结果一致[11,20]。此外,华山松大小蠹对8种非寄主挥发物的触角电位反应值随浓度的升高而增强,与华山松大小蠹对寄主挥发物的反应有差异,高浓度的寄主挥发物会抑制其EAG反应[21-22],这说明华山松大小蠹对非寄主挥发物与寄主挥发物的反应机制可能存在差异。华山松大小蠹对反-2-己烯-1-醇、反-2-己烯醛、1-辛烯-3-醇和3-辛醇的触角电位反应虽有一定的差异,但是差异并不极显著(p>0.01),表明华山松大小蠹对多种非寄主挥发物的反应具有相似性,这可能是因为在小蠹虫寄主搜寻的过程中,非寄主挥发物作为一类化学信息物质是在生境水平上发挥驱避作用,所以小蠹虫需要识别并避开非寄主阔叶树普遍释放的挥发物,而不是针对某种非寄主的特定挥发物[23]。室内趋向反应表明,测定的挥发物中顺-3-己烯-1-醇、1-辛烯-3-醇、3-辛醇、1-己烯-3-醇、苯甲醛和反-2-己烯醛在一定浓度下对华山松大小蠹成虫表现出明显的驱避作用,说明华山松大小蠹在寄主选择过程中可能会避开产生该类物质的非寄主树木。3-辛醇引起华山松大小蠹的触角电位反应最弱,但其对华山松大小蠹驱避作用最强,这可能是因为触角电位反应只是触角感觉器官的参与,不能代表整个有机体的复杂系统[24],此外小蠹虫对非寄主挥发物的触角电位反应和行为反应二者之间的关系也具有复杂性,触角电位反应值与驱避作用并不是单纯的正相关或负相关。
对于小蠹虫寄主选择过程的研究是开发信息化合物防控小蠹虫技术的关键环节,近年来,通过对寄主华山松的挥发物的筛选,已经明确了α-蒎烯、β-蒎烯、莰烯、3-蒈烯等对华山松大小蠹的引诱效果,并已初步应用于野外防治[25-27]。将非寄主挥发物作为驱避剂与寄主挥发物或者性信息素结合应用的“推-拉”策略要优于单纯用引诱剂(“拉”)的效果,有助于推动小蠹虫综合管理水平的提高。
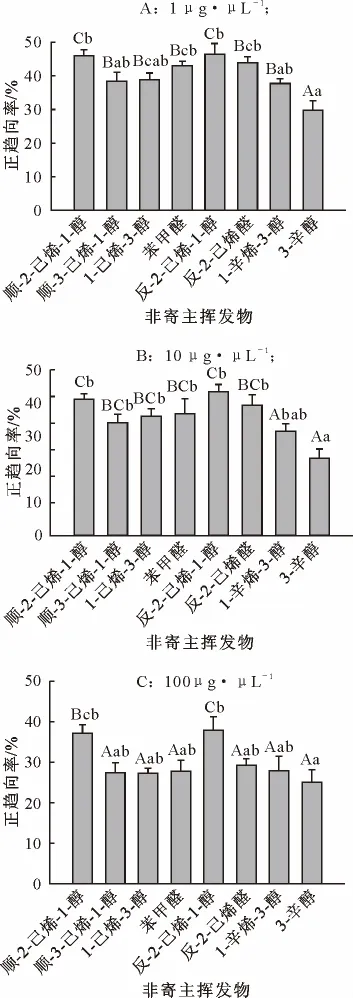
图2 华山松大小蠹成虫对非寄主挥发物的行为反应Fig.2 Behavioral responses of D.armandi to nonhost volatiles
本试验测定了华山松大小蠹对几种非寄主挥发物的电生理反应及室内趋向反应,表明顺-3-己烯-1-醇、1-辛烯-3-醇、3-辛醇、1-己烯-3-醇、苯甲醛和反-2-己烯醛对其有较强的驱避作用,其中3-辛醇的驱避作用最强,说明利用这5种化合物可能配制出有效的驱避剂,但是否可以达到理想林间引诱效果及应如何配比还待进一步研究。此外,这几种化合物对华山松大小蠹聚集信息素及寄主挥发物的抑制作用是否存在,有待于通过进一步的研究确认。
[1] 李成德.森林昆虫学[M].北京:中国林业出版社,2004.
[2] WOOD D L.The role of pheromones,kairomones and allomones in the host selection and colonization of bark beetles[J].Annu Rev.Entomol,1982,27:411-446.
[3] KELLEY S T,FARRELL B D.Is specialization a dead end?The phylogeny of host use in Dendroctonus bark beetles(Scolytidae)[J].Evolution,1998,52:1731-1743.
[4] SEQUEIRA A S,NORMARK B B,FARRELL B D.Evolutionary assembly of the conifer fauna:distinguishing ancient from recent association in bark beetles[J].Proceedings of the Royal Society Biological Series B,2000,267:2359-2366.
[5] RAFFA K F,BENYMAN A A.Interacting selective pressures in conifer bark beetle systems:a basis for reciprocal adaptations[J].American Naturalist,1987,129:234-262
[6] HRDY I.Insect chemical ecology[M].Prague:Acadamia Payne,1991:277-280.
[7] SCHROEDER L M.Olfactory recognition of nonhosts aspen and birch by conifer bark beetles Tomicus piniperda and Hy lurgops palliates[J].J.Chem.Ecol.,1992,18:1583-1593.
[8] MITTON J B,STURGEON K B.Bark beetles in North American conifers:A system for the study of evolutionary biology[M].Austin:University of Texas Press,1982:140-182.
[9] ATKINS M D.Lipid loss with flight in the douglas-fir beetle[J].The Canadian Entomologist,1969,101(2):164-165.
[10] ZHANG Q H,SCHLYTER F.Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles[J].Agricultural and Forest Entomology,2004,6(1):1-19.
[11] ZHANG Q H,LIU G T.Olfactory responses of Ips duplicatus from Inner Mongolia,China to nonhost leaf and bark volatiles[J].Journal of Chemical Ecology,2001,27(5):995-1009.
[12] POLAND T M,DEGROOT P,BURKE S,et al.Semiochemical disruption of the Pine Shoot Beetle,Tomicus piniperda (Coleoptera:Scolytidae)[J].Environmental Entomology,2004,33(2):221-226.
[13] UNELIU CR,SCHIEBE C,BOHMAN B.Non-host volatile blend optimization for forest protection against the Eurpean Spruce bark beetle,Ips typographus[J].Plos One,2014,9(1):1-10.
[14] WILLIAM PS,BRIAN TS.Southern Pine Beetle,Dendroctonus frontalis,antennal and behavioral responses to nonhost leaf and bark volatiles[J].J.Chem.Ecol.,2014,39:481-493.
[15] PROGAR R A,GILLETTE N,FETTIG C J.Applied chemical ecology of the mountain pine beetle[J].FOREST SCIENCE.2014,60(3):414-433.
[16] WEI J N,KANG L.Roles of(Z)-3-hexenol in plant-insect interactions[J].Plant Signaling &Behavior,2011,6(3):369-371.
[17] 李建光,骆有庆.复叶槭挥发性物质对光肩星天牛的触角电位反应[J].北京林业大学学报,1999,21(4):1-5.LI J G,LUO Y Q.Electroantennogram activity of ash-leaf maple(Acer negundo)volatiles to Anoplophora glabripennis(Motsch.)[J].Journal of Beijing Forestry University,1999,21(4):1-5.(in Chinese)
[18] 李为争,杨雷,申小卫.金龟甲对蓖麻叶挥发物的触角电位和行为反应[J].生态学报,2013,33(21):6895-6903.LI W Z,YANG L,SHEN X W.Electroantennographic and behavioural responses of scarab beetles to Ricinus communis leaf volatiles[J].Acta Ecologica Sinica,2013,33(21):6895-6903.(in Chinese)
[19] 周琦.非寄主植物挥发物对云杉八齿小蠹危害行为的干扰[D].哈尔滨:东北林业大学,2011.
[20] ZHANG L W,NANCY E.Gillette.Electrophysiological and behavioral responses of Dendroctonus valens to non-host volatiles[J].Annals of Forest Science,2007,64(3):267-273.
[21] ZHANG L L,CHEN H,MA C.Electrophysiological responses of Dendroctonus armandi(Coleoptera:Curculionidae:Scolytinae)to volatiles of Chinese white pine as well as to pure enantiomers and racemates of some monoterpenes[J].Chemoecology,2010,20:265-275.
[22] 王茹琳,杨 伟,杨佐忠.华山松大小蠹对几种寄主挥发物组分的EAG 和行为反应[J].生态学杂志,2011,30(4):724-729.WANG R L,YANG W,YANG Z Z.Electroantennographic and behavioral responses of Dendroctonus armandi(Tsai et Li)to host plant volatiles[J].Chinese Journal of Ecology,2011,30(4):724-729.(in Chinese)
[23] REDDY G V P,GUERRERO A.Interactions of insect pheromones and plant semiochemicals[J].Trends in Plant Science,2004,9(5):253-261.
[24] 黄恩烱.8种昆虫对蓖麻毒素和相思豆毒素触角电位及行为反应的研究[D].北京:中国人民解放军军事医学科学院,2008.
[25] 吴绍平,陈辉,吴琼.华山松大小蠹成虫粪便挥发性物质分析[J].西北林学院学报,2012,27(2):111-116.WU S P,CHEN H,WU Q.Volatile compounds in the frass of adult Chinese white pine beetle[J].Journal of Northwest Forestry University,2012,27(2):111-116.(in Chinese)
[26] 赵明振,陈辉,马超.华山松大小蠹对华山松挥发性物质行为反应的研究[J].西北林学院学报,2014,30(5):115-119.ZHAO M Z,CHEN H,MA C.Behavioral responses of Dendroctonus armandi (Coleoptera:Curculionidae:Scolytinae)to volatiles of Chinese White Pine[J].Journal of Northwest Forestry University,2014,30(5):115-119.(in Chinese)
[27] XIE S A,LV S J.An improved lure for trapping the bark beetle Dendroctonus armandi (Coleoptera:Scolytinae)[J].Eur.J.Entomol,2012,109:569-577.
