甘蓝型油菜CRABS CLAW基因克隆及其RNA干扰载体的构建
2015-01-01周晓婴付三雄戚存扣
周晓婴, 付三雄, 陈 松, 张 超, 戚存扣
(1.国家油菜改良中心南京分中心,江苏 南京 210014;2.农业部长江下游棉花与油菜重点实验室,江苏 南京 210014;3.江苏省农业科学院经济作物研究所,江苏 南京 210014)
植物雌性不育通常由雌性器官发育异常引起,雌性不育表型具有多样性,不育机理复杂。了解其细胞分子机理对揭示植物花器官发育的分子机理以及在生产中克服不育性和利用雌性不育作为杂交育种手段等都具有重要的理论与实践意义[1]。
甘蓝型油菜雌性不育突变体FS-M1于1996年在甘蓝型油菜(Brassica napus L.)品种宁油10号的135个株系群体中偶然被发现。形态观察发现突变株FS-M1雌蕊明显短于雄蕊,无蜜腺,柱头乳突细胞较大但数量较少,粘附花粉能力弱,柱头授粉功能缺陷。切除柱头及部分花柱后再授粉,在FS-M1花柱中观察到伸长的花粉管,授精结角也接近正常。FS-M1雌性不育现象可能与柱头乳突细胞发育畸变和功能缺失有关,与自交不亲和无关[2-3]。遗传研究结果显示FS-M1的突变性状由2对隐性基因控制,遗传性状稳定,植株花粉育性正常[4]。
付三雄曾利用Agilent公司生产的油菜全基因组4×44K芯片比较了油菜FS-M1及其野生型花器官的转录组,发现突变体花器官组织1个CRABS CLAW(CRC)转录因子的基因表达量显著下调。本研究为了进一步验证CRC转录因子在油菜花器官发育中的作用,以及导致油菜FS-M1雌性不育性状的决定因素是否与CRC基因表达量显著下调有关,根据同源序列克隆了甘蓝型油菜CRC基因,并构建CRC基因干扰表达载体,拟通过转基因来验证CRC基因的功能,明确CRC基因下调表达与FS-M1雌性不育的关系。
1 材料与方法
1.1 材料
克隆载体pEASY-T1及感受态大肠杆菌DH5ɑ购自南京百斯凯公司,RNA提取试剂盒为Promega公司产品,琼脂糖凝胶回收试剂盒及细菌质粒小量提取试剂盒均购自北京天根生物有限公司,Ex Taq、DNA Marker、RNA反转录试剂盒、各种限制性内切酶(如 EcoR I、Xho I、Hin III、BamH I、Not I等)、T4连接酶、碱性磷酸酶(Alkaline Phosphatase)均购自TaKaRa公司,X-gal、IPTG、酵母提取物、蛋白胨、抗生素等购自南京基天生物公司。测序由上海英骏公司完成。
中间载体pHurricane、表达载体 pCAMBIA1390系本实验室保存。
1.2 CRC基因克隆
本试验依据已经公开的甘蓝型油菜CRC基因cDNA序列(登录号为XM00910 7216)设计RT-PCR引物。引物同时引进 NotⅠ和 XhoⅠ酶切位点,CRCXNFF:5'-CTCGAGCGGCCGCATGAACCTTGAA GAGAAACC-3',CRCXNR R:5'-CTCGAGCGGCCGCATTTCCTTGGGCTATCACCTT-3',以甘蓝型油菜品种宁油10号花蕾RNA反转录cDNA为模板,采用Ex Taq酶PCR扩增靶基因序列片段,将目标条带克隆到pEASY-T1载体上,挑选阳性克隆测序,并与目标序列进行比对分析,获得含CRC基因片段的克隆载体pT-CRC。
1.3 CRC基因反向重复框构建
根据中间载体pHurricane内含子序列设计特异性引物:INTRONFW:5'-CGTATTCTCAACGCAATC-3',和 INTRONRV:5'-CAAAGTGGGAAATCAGGT-3',用于鉴别靶序列插入方向。用 Not I酶切pT1-CRC,回收580bp目的片段。同时利用Not I酶切pHurricane载体,并进行末端去磷酸化处理后,再与目的片段连接,连接产物转化感受态大肠杆菌。采用INTRONRV和CRCXNF引物对鉴定正向插入的阳性克隆,此时获得的中间载体命名为pH-CRCF。再用Xho I酶切pT-CRC,回收580 bp目的片段,用Xho I酶切pH-CRCF载体,并去磷酸化处理、纯化后与目的片段连接,连接产物转化大肠杆菌。用INTRONFW和CRCXNF引物对进行菌落PCR,选择反向插入的阳性克隆。此时获得的中间载体命名为pH-CRCir,包含目的基因反向重复框。
1.4 CRC干扰表达载体的组装及酶切鉴定
本试验选择CaMV35S启动子作为CRC反向重复序列表达框的启动子。以pCAMBIA1390质粒DNA为模板,设计特异性扩增引物,在35SHF 5'端加Hind III的酶切位点,在35SBR 5'端增加BamH I的酶切位点。35SHF:5'-AAGCTTATGGTGGAGCACGACACTCT-3',35SBR:5'-GGATCCAGAGATAGATTTGTA GAGAG-3',扩增35S序列,先将扩增产物克隆到pEASY-T载体上,再用Hind III和Bam H I双酶切获得780bp大小的35S启动子序列,再组装到pCAMBIA1390载体相应的酶切位点上,得中间载体pCAMBIA1390-35S。
用BamHⅠ和EcoRⅠ分别双酶切pCAMBIA1390-35S和pH-CRCir,回收纯化线性化的pCAMBIA1390-35S载体片段和大小约2.3 kb的CRC正反向重复框片段,将回收产物用T4连接酶连接后转化大肠杆菌。用CRCXNF和CRCXNR引物进行菌落PCR鉴定阳性克隆,并提取阳性克隆质粒DNA,分别用单酶切(HindⅢ)和双酶切(BamHⅠ和EcoRⅠ)进行鉴定并测序,获得的载体定名为pA6-CRCi。
2 结果
2.1 油菜CRC基因片段克隆与序列分析
本研究对甘蓝型油菜宁油10号花蕾RNA进行RT-PCR,扩增到1个约580 bp大小的产物(图1)。经克隆测序,并用BioXM软件对其核酸序列进行分析,结果显示该cDNA全长580 bp,具有1个编码193个氨基酸的开放式阅读框。
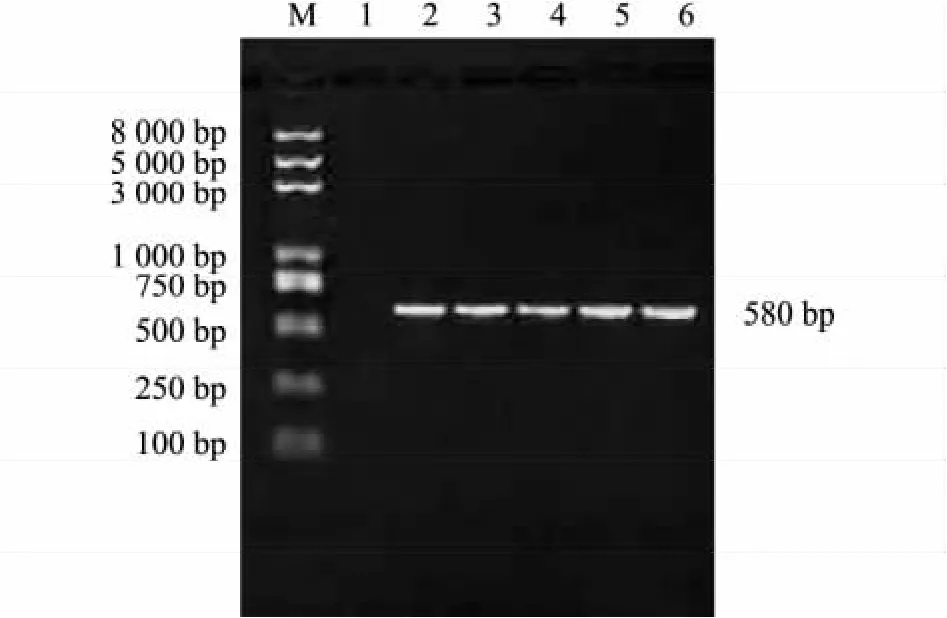
图1 CRC基因克隆菌落PCR鉴定结果Fig.1 The PCR amplification of CRC gene clones
用BLAST在线分析软件进一步将所克隆的cDNA序列与GenBank中登录的油菜、拟南芥等多种植物的CRC基因cDNA序列进行同源性比较,发现该核苷酸序列与油菜、拟南芥、非洲独行菜、芸薹、甘蓝等植物中的CRC基因序列高度同源,相似性大于90%,其中与甘蓝型油菜CRC基因的cDNA序列(XM 009107216)同源性达99%,表明所克隆序列即为油菜CRC基因序列,此克隆定名为pT-CRC。
2.2 油菜CRC基因反向重复表达框构建及酶切鉴定
将2.1克隆到的CRC基因片段分2步,依次克隆到中间载体pHurricane内含子两侧的Not I和Xho I限制性内切酶位点。分别利用内含子上的引物INTRONRV和INTRONFW与干扰片段的上游引物CRCNX F组合进行菌液PCR鉴定,可以筛选到干扰片段正向插入Not I位点的阳性克隆和反向插入Xho I位点的阳性克隆(图2)。

图2 PCR鉴定CRC基因片段插入正反方向克隆Fig.2 PCR identification of CRC positive clones
培养经PCR鉴定后的阳性克隆菌株,提取质粒,分别用Not I和Xho I限制性内切酶酶切,均可以切出大小约580 bp的条带(图3)。进一步用BamH I和EcoR I限制性酶进行双酶切,可以获得大小2.29 kb左右的片段(图3),该片段包含有正向插入、反向插入的干扰片段和内含子片段,表明CRC基因片段已成功连接到中间载体的相应酶切位点上。
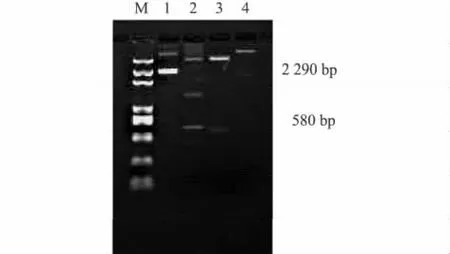
图3 酶切鉴定pH-CRCirFig.3 Enzymatic identification of pH-CRCir
2.3 CRC基因ihpRNAi植物表达载体构建
为构建组成型表达的CRC基因的干扰载体,本研究选择CaMV35S启动子作为CRC反向重复表达框的启动子。首先通过PCR从pCAMBIA1390上扩增1个780 bp的增强型CaMV35S启动子,再经过酶切与连接等步骤,将 CaMV35S启动子连接到pCAMBIA1390双元表达载体的Hind III和BamH I位点上,再用BamH I和EcoR I双酶切,获得2.29 kb的CRC基因反向重复框片段,与同样双酶切的pCAMBIA1390-35S载体片段连接后最终形成CRC基因的干扰表达载体 pA6-CRCi。pA6-CRCi经BamH I和EcoR I双酶切验证,酶切图谱中含有2.29 kb大小的CRC反向重复表达框片段(图4)。表明CRC基因的干扰重复框已经成功连接到带pCAMBIA1390-35S双元表达载体上。最终形成的pA6-CRCi载体含有1个由CaMV35S启动子驱动的CRC反向重复序列表达框,终止子是原载体自带的nos终止子。
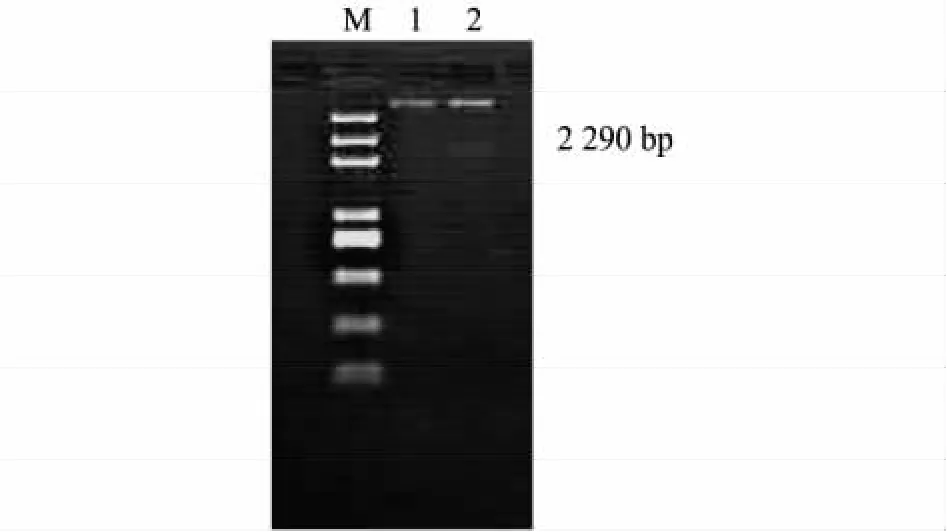
图4 BamHⅠ和EcoRⅠ酶切鉴定pA6-CRCiFig.4 Enzymatic identification of pA6-CRCi with BamHⅠand EcoRⅠ
3 讨论
植物的雌性不育现象普遍存在,在水稻[5]、油松[6]、小麦[7]、甘蓝型油菜[2]、大豆[8]和苜蓿[9]等十余种植物中均报道过雌性不育现象。导致植物雌性不育的原因比较复杂,而远缘杂交后代发现雌性不育的报道比较多。Stort[10]将兰花的属间或种间F1代与母本回交,发现其回交后代结实率下降,且雌性不育率远远大于雄性不育率。刘永胜等[11]观察到水稻籼粳F1杂种的结实率仅为40% ~60%,其主要原因是由雌性不育引起。离体培养易产生无性系变异,也是产生雌性不育突变体的原因之一[5]。环境的变化也会影响植物的育性,Lillecrapp等[12]在Trevatt Blue的研究中发现,温度可以影响到胚珠的数量,进而形成多胚珠现象而导致雌性不育。
植物雌性不育的原因相当复杂,一些表现为大孢子母细胞减数分裂不正常,不能形成功能性大孢子。Rim[13]等研究发现豆科百脉根属2个种的体细胞杂交后代由于大孢子母细胞减数分裂后期发生异常,不能形成4个大孢子而导致雌性不育;也有的是因为不能形成成熟的胚囊[14]或胚珠发育异常而导致不结实[15]。此外,雌蕊发育异常,如多雌和雌蕊短小等突变现象,也会引起育性降低或不育[16]。近几年来人们应用遗传学方法与分子生物学技术研究植物雌性不育的分子机理,发现了一些与雌性不育相关的基因。Lang等[17]研究证实,拟南芥sinl基因的突变会导致胚珠畸形发育与雌性不育的发生。Tian等[18]发现拟南芥中组蛋白脱乙酰化基因(AtHD1)表达下调会导致许多异常现象产生,包括花器官缺陷及雌性和雄性不育。Capomaccio等[19]对紫花苜蓿雌性不育突变株和可育对照株杂交分离后代进行了cDNA-AFLP扩增分析,筛选并鉴定了3个与雌性不育性状相关的基因MsBGluc、MsMAPKKK和MselF4G。
大多数植物的雌性不育在花的外观特征上都有所表现。如Carapetian[20]等在红花的雌性不育研究中观察到2个红花品种杂交产生的不育植株开花时会发生小花的干枯和花柄的延长。一些学者试图从花器官发育调控方面去寻找雌性不育的内在原因,其中花发育调控相关的一些转录因子受到人们的普遍关注,如YABBY家族的INNER NO OUTER(INO)参与了拟南芥胚珠的发育[21];TONGARI-BOUSHI1(TOB1)参与了水稻小穗的发育[22]。而拟南芥CRABS CLAW(CRC)转录因子也属于YABBY基因家族成员,是控制心皮发育的关键性基因,Alvarez等[23]报道CRC可抑制正在发育的雌蕊的横向生长,促进其轴向生长,CRC基因突变将引起雌蕊呈现较宽而短的结构,且使两心皮在顶点处呈非融合状态,并导致蜜腺的缺失[24-25]。心皮发育还有SPT(SPATULA)基因[26-27]及花器官确定基因AGA(AGAMOUS)的参与[28]。从一些心皮突变体的研究中,首先认为CRC基因很有可能是AGA调节的下游基因[25]。对分离到的2个突变基因CRC、SPT的研究发现,二者除了影响雌蕊的发育之外还导致其他不同的生理缺陷,通过对CRC、SPT和AGA以及其他的相关基因的详细分析,发现CRC和SPT 2个基因都与心皮的发育有关,在心皮发育过程中CRC和SPT是与AGA平行起作用,而不是位于AGA的下游。
FS-M1突变体是研究甘蓝型油菜柱头发育分子机理的理想材料。李春宏等[4]基于蛋白质组学水平比较甘蓝型油菜野生型和FS-M1的柱头差异蛋白质,获得了甘蓝型油菜野生型柱头显著上调且可鉴定的蛋白质有33个,包括抗胁迫/防御、氧化还原反应平衡调节、碳水化合物代谢、蛋白质水解、蛋白折叠、氨基酸和氮代谢、核苷酸代谢等7类功能蛋白。李春宏等[29]以FS-M1突变体和其野生型为材料利用油菜基因表达谱芯片筛选甘蓝型油菜柱头特异表达基因,发现野生型较FS-M1显著上调的基因多达198个,主要是水解酶基因、转移酶基因、氧化还原酶基因和转录因子中。但目前尚没有详实的证据阐明油菜突变体FS-M1雌性不育的原因。
我们对油菜雌性不育突变体FS-M1初步研究发现,其CRC基因表达水平较野生型对照极显著下调,结合先前的研究报道FS-M1突变体柱头短小且异常,因此我们推测油菜CRC基因下调可能是油菜雌性不育突变体FS-M1的内在原因。为此,我们构建了CRC基因的RNAi干扰载体,以期通过下调CRC表达,验证其在FS-M1突变体中所起的作用。本研究利用同源序列法通过PCR扩增克隆了油菜CRC基因序列,经过与已知序列进行比对分析,证明本试验所克隆的基因序列正是甘蓝型油菜的CRC基因序列。我们利用 pHurricane载体构建CRC反向重复序列表达框,该表达框有1个拟南芥的FAD2基因的可剪切的内含子序列,两边有多个限制性内切酶位点可供选择。将CRC基因片段分别插入该载体的Not I和Xho I位点,结合内含子序列设计的引物,可以比较方便地选择片段插入的方向。应用该载体我们成功构建了多个基因的干扰载体[30-31]。在本研究中我们选择了增强型的CaMV35S启动子,该启动子全长序列780 bp左右,5'端有一段重复序列,可以起到增强表达的作用,用该启动子驱动CRC的反向重复序列表达框,目的是提高组成型表达的强度,达到彻底抑制CRC表达的效果。本试验成功构建组成型表达的CRC基因干扰载体,经过酶切验证和序列分析,表明所构建的载体正确,这为进一步开展农杆菌遗传转化油菜奠定了基础。
[1] 胡 青,高述民,李凤兰.植物雌性不育的研究进展[J].北京林业大学学报,2004,26(1):87-91.
[2] 陈新军,戚存扣,张洁夫,等.油菜(Brassica napus L.)雌性不育突变体FS-M1的生物学特性研究[J].中国油料作物学报,2003,25(3):12-15.
[3] 陈新军,戚存扣,张洁夫,等.甘蓝型油菜雌性不育突变体FSM1花器官形态结构研究[J].作物学报,2005,31(9):1239-1240.
[4] 李春宏,付三雄,陈新军,等.甘蓝型油菜雌性不育突变体M7乳突细胞的细胞学观察[J].植物学报,2012,47(1):36-43.
[5] 凌定厚,马镇荣,陈梅芳,等.起源于体细胞培养的籼稻雌性不育突变[J].遗传学报,1991,18(5):446-451.
[6] 丁坤善,郑彩霞,包仁艳,等.油松雌性不育系球果蛋白质双向电泳技术的建立[J].植物学通报,2005,22(2):190-197.
[7] 窦秉德,张新玲,马 林,等.普通小麦中一种雌性不育现象的观察[J].作物学报,2001,27(6):1013-1016.
[8] 赵团结,盖钧镒.大豆叶与花形态异常、雌性不育突变体NJS-10H 的发现[J].大豆科学,2005,24(1):1-4.
[9] 张静文,金 樑,冯光辉,等.苜蓿雌性不育发生机制及其分子调控研究进展[J].云南农业大学学报,2011,26(4):560-566.
[10] STORT.Sterility barriers in some artificial F1orchid hybrids:female sterility[J].Rev Bras Genet,1987,10(1):109-118.
[11]刘永胜,周开达,阴国大,等.水稻籼梗杂种雌性不育的细胞学初步观察[J].实验生物学报,1993,26(1):95-99.
[12] LILLECRAPP A M,WALLWORK M A,SEDGLEY M.Female and male sterility cause low fruit set in a clone of the“Trevatt”variety of apricot(Prunus armeniaca) [J].Scientia Horticulturae Amsterdam,1999,82(3-4):255-263.
[13] RIM Y W,BEUSELINCK P R,MCGRAW R L,et a1.Megagametophyte development in Lotus corniculatus L.coninrbricensis and their protoplast fusion hybrid[J].Lotus Newsletter,1998,19:45-47.
[14] OUYANG X Z.Ultrastructure and AcPase ultracytochemical localization of the abortive functional megaspores in intersubspecific F1hybrid rice(Oryza sativa)[J].Chinese Rice Research Newsletter,1996,10(4):2-3.
[15]张宪省,马小杰.胚珠发育的分子基础[J].植物学通报,1998,15(3):1-5.
[16] PENMESTA R V,COOK D R.Production and characterization of diverse developmental mutants of Medicago truncatula[J].Plant Physiology,2000,123(4):1387-1397.
[17] LANG J D,RAY S,RAY A.Sinl,a mutation afecting female fertility in Arabidopsis,interacts with mod1,its recessive modifier[J].Genetics,1994,137(4):1101-1110.
[18] TIAN L,CHEN Z J.Blocking histone deacetylation in Arabidopsis induces pleiotuopic effects on plant gene regulation and development[J].Proceedings of the National Academy of Sciences of the United States of America,2001,98(1):200-205.
[19] CAPOMACCIO S,BARONE P,REALE L,et a1.Isolation of genes from female sterile flowers in M edicago satiu[J].Sexual Plant Reproduction,2009,22:97-107.
[20] CARAPETIAN J,KNOWLES P F.Genetic linkage between the tri-genic male-female sterility and oil quality alleles in saflower[J].Crop Science,1993,33(2):239-242.
[21] GALLAGHER T L,GASSER C S.Independence and interaction of regions of the INNER NO OUTER protein in growth control during ovule development[J].Plant Physiology,2008,147:306-315.
[22] TANAKAA W,TORIBAA T,OHMORIA Y,et a1.The YABBY gene TONGARI-BOUSHI1 is involved in lateral organ development and maintenance of meristem organization in the rice spikelet[J].The Plant Cell,2012,24:80-95.
[23] ALVAREZ J,SMYTH D R.CRABS CLAW and SPATULA,two Arabidopsis genes that control carpel development in parallel with AGAMOUS[J].Development,1999,26:2377-2388.
[24] LEE J Y,BAUM S F,ALVAREZ J,et al.Activation of CRABS CLAW in the nectaries and carpels of Arabidopsis[J].The Plant Cell,2005,17:25-36.
[25] FOURQUIN C,VINAUGER-DOUARD M,FOGLIANI B,et a1.Evidence that CRABS CLAW and TOUSLED have conserved their roles in carpel development since the ancestor of the extant angiosperms[J].PNAS USA,2005,102:4649-4654.
[26] GROSZMANN M,BYLSTRA Y,LAMPUGNANI E R,et a1.Regulation of tissue-specific expression of SPATULA,a bHLH gene involved in carpel development,seedling germination,and lateral organ growth in Arabidopsi[J].Journal of Experimental Botany,2010,61:1495-1508.
[27] GROSZMANN M,PAICU T,SMYTH D R.Functional domains of SPATULA,a bHLH transcription factor involved in carpel and fruit development in Arabidopsis[J].Plant J,2008,55:40-52.
[28] GOIZ,JOHN F,HUDSON,et al.Signaling in plant lateral organ development[J].The Plant Cell,2002(Suppl):277-288.
[29]李春宏,付三雄,戚存扣.应用基因芯片分析甘蓝型油菜柱头特异表达基因[J].植物学报,2014,49(3):246-253.
[30]陈 松,张洁夫,陈 锋,等.甘蓝型油菜种子特异性表达fad2基因的ihpRNA载体构建[J].中国油料作物学报,2006,28(3):25l-256.
[31]付三雄,张 超,陈 松,等.甘蓝型油菜丙酮酸激酶基因RNA干扰载体的构建[J].江苏农业学报,2013,29(1):20-27.
