南岭山地森林群落冠层结构与立木多度的关系
2014-12-29敬小丽杜伟静张璐苏志尧
敬小丽,杜伟静,张璐,苏志尧
(华南农业大学 林学院,广东 广州 510642)
南岭山地森林群落冠层结构与立木多度的关系
敬小丽,杜伟静,张璐,苏志尧
(华南农业大学 林学院,广东 广州 510642)
林冠是森林与外部环境相互作用最直接与最活跃的界面层,影响着森林的物理环境和生物环境。采用半球面摄影技术(Hemispherical Photography)、典范对应分析(Canonical Correspondence Analysis, CCA)和Kruskal-Wallis分析定量研究了森林群落冠层结构与立木分布及多度的关系。结果表明:(1)基于10000m2调查样地,南岭山地森林群落共有立木47科81属143种,枯立木多来源于林冠上层大树;(2)CCA排序结果显示,冠层结构各参数与立木分布均有一定相关性,叶面积指数、林冠开度与立木分布的关系尤为密切;(3)立木多度与活立木多度在林下散射光分组变量间差异显著(p< 0.05),枯立木多度在林冠开度分组变量间的差异性显著(p< 0.05),立木多度随林下直射光增强而减少。叶面积指数和林冠开度通过影响林下光照条件进而影响立木,尤其是枯立木的形成和多度。
山地森林群落;冠层结构;立木多度;活立木;枯立木;叶面积指数;林冠开度
林冠(Canopy)是由森林上部郁闭的枝叶和层内空气所组成的,是某一范围内所有树冠(包括枝叶)的集合体[1]。林冠通过截留降水影响森林水文循环[2-3],通过对光的吸收、反射和折射影响林下光照条件[4-5],还为某些节肢动物提供了栖息场所[6],从而有利于维持生物多样性。物理环境和生物环境的异质性影响森林环境及其生物进程[7]。在全球变化的大背景下,人们越来越深刻地认识到森林是气候变化的良好指示器[8]。林冠开度(Canopy Openness, CO)与叶面积指数(Leaf Area Index,LAI)是冠层结构的基本参数。同一树冠内的光照强度存在上下和内外的差异[9],森林冠层结构也强烈影响着林下光照条件[10],通过控制太阳能的截获量来调节林下直射光(Transmitted Direct Light,TDir)、散射光(Transmitted Diffused Light, TDif)和总光照(Transmitted Total Light, TTot)等[4]。运用半球面影像技术(hemispherical photography)可以快速、方便地测定冠层结构参数,该技术现已广泛应用于生态学领域的研究,如特定树种在不同区域的冠幅差异[11]、估算林冠属性[12-13]、估算冠层孔隙度[14]以及干扰对冠层结构的影响[4]。为了探讨森林群落冠层结构与立木多度的关系,以及冠层结构对枯立木形成的影响,本研究以南岭山地森林群落的立木(活立木与枯立木)为研究对象,定量研究了冠层结构参数与立木多度之间的相关性及其大小,为种间与种内竞争分析提供研究思路和参考依据。
1 研究地概况
研究样地位于广东省南岭国家级自然保护区(24º37′~ 24º57′ N,112º30′~ 113º04′ E),东邻乳源县,南接阳山岭背镇,西靠连州市,北部与湖南省宜章县莽山森林公园接壤,总面积58 368.4 hm2,是目前广东省陆地面积最大的保护区。该区属于亚热带季风气候区,年平均温度17.4 ℃,最冷月(1月)平均气温7.1 ℃,最热月(7月)平均气温26.2 ℃,年均降水量1 705 mm,多集中于8月。年相对湿度84%,冬季偶有降雪,日照率40%。水平地带性土壤多为红壤,分布的土壤类型随海拔高度的不同而不同。该区树种组成丰富,乔木层主要以金缕梅科Hamamelidaceae、松科Pinaceae、山茶科Theaceae、杜鹃花科Ericaceae及壳斗科Fagaceae占优势,优势种主要有檵木Loropetalum chinensis、华南五针松Pinus kwangtungensis、五列木Pentaphylax euryoides、猴头杜鹃Rhododendron simiarum、疏齿木荷Schima remotiserrata[15]。
2 研究方法
2.1 林分调查
在线路勘察的基础上,采用样方法设置样地,以10m×10m为样方单元,共计调查了100个样方(10000m2)。在每个样方单元内进行每木调查,测定胸径(DBH)≥ 3 cm的所有活立木的种名、胸径、树高和冠幅。同时根据与枯立木丛生的个体、根部萌枝、树形或木质等特征,记录DBH ≥ 5 cm且高度不低于2 m的枯立木种名、胸径和树高。
2.2 半球面林冠影像拍摄与分析
选择阴天或无风的天气,在日出或日落的时间[16],用三脚架将Nikon CoolPix 4500数码相机外接Nikkor FC-E8鱼眼镜头转换器水平放置于离地面1. 65 m 处,用指南针确定方向使记录的照片顶部与磁北方向重合,在每个10m×10m样方单元中心和对角线四分位处镜头朝上拍摄半球面林冠影像。采用Gap Light Analyzer 2.0(GLA)图像处理软件分析林冠影像,以输出的林冠开度(CO)、叶面积指数(LAI)、林下直射光(Transmitted Direct Light, TDir)、散射光(Transmitted Diffused Light, TDif) 和 总 光 照(Transmitted Total Light, TTot)等5个指标作为冠层结构参数。
2.3 数据分析
在CANOCO 4.5 中采用典范对应分析(Canonical Correspondence Analysis, CCA)方法分析活立木和枯立木在样地中的分布状况及其与林冠开度、叶面积指数、林下直射光、散射光和总光照5个冠层结构参数之间的关系。对数据进行蒙特卡罗检验(Monte Carlo Test),并计算冠层结构参数之间的相关系数及其与立木排序轴之间的相关系数。
采用Statistica 8.0统计软件对活力木和枯立木的多度与5个冠层结构参数分别进行Kruskal-Wallis(非参数ANOVA)分析,检验立木多度在各分组变量(见表1)间是否存在差异,并对其中差异性显著的进行多重比较。

表1 冠层结构参数分组†Table 1Groups of canopy structure parameters
3 结果与分析
3.1 立木组成与数量特征
基于10000m2的样方调查数据,南岭山地森林共计有立木47科81属143种,其中活立木47科81属143种,枯立木12科17属17种。在143种活立木中,以五列木科Pentaphylacaceae的五列木Pentaphylax euryoides和甜茶椆Lithocarpus polystachyus、壳斗科的青冈Cyclobalanopsis glauca以及杜鹃花科的猴头杜鹃和羊角杜鹃Rhododendron cavaleriei为优势种(见表2),此外还有金缕梅科的大果马蹄荷Exbucklandia tonkinensis和檵木、茶科的疏齿木荷和杨桐Adinandra millettii以及杜鹃花科的石壁杜鹃Rhododendron bachii。

表2 南岭山地森林群落优势立木数量特征†Table 2Quantitative characteristics of dominant standing trees in Nanling mountain forests
在17种枯立木中,以安息香科的赤杨叶Alniphyllum fortunei、八角科Illiciaceae的毒八角Illicium toxicum、壳斗科的罗浮锥Castanopsis fabri、青冈以及冬青科Aquifoliaceae的小果冬青Ilex micrococca为优势种,其他枯立木种类还有檫木Sassafras tsumu、长苞铁杉Tsuga longibracteata、檵木、金叶含笑Michelia foveolata、柃木Euyra japonica等。赤杨叶、毒八角、檵木、罗浮锥、南亚新木姜Neolitsea zeylanica、青冈和五列木是活立木和枯立木的共同优势木。从胸径上看,赤杨叶、檵木、罗浮锥的枯立木胸径比活立木小,而毒八角和青冈的枯立木胸径比活立木大,南亚新木姜和五列木枯立木的平均胸径与活立木相近;从树高上看,仅赤杨叶和五列木枯立木的树高明显比活立木小,其它树种的树高均比活立木大或与之相近。表明林冠上层大树枯死较多,成为枯立木的重要来源。
3.2 冠层结构与立木分布的关系
排除多度≤5的物种,对84种常见立木进行多度与冠层结构的典范对应(CCA)分析,结果显示,四个排序轴的特征值分别为0.381、0.207、0.115和0.110,特征值与第一轴和全部轴均通过蒙特卡罗检验(P< 0.05),说明排序结果可信(见表3)。
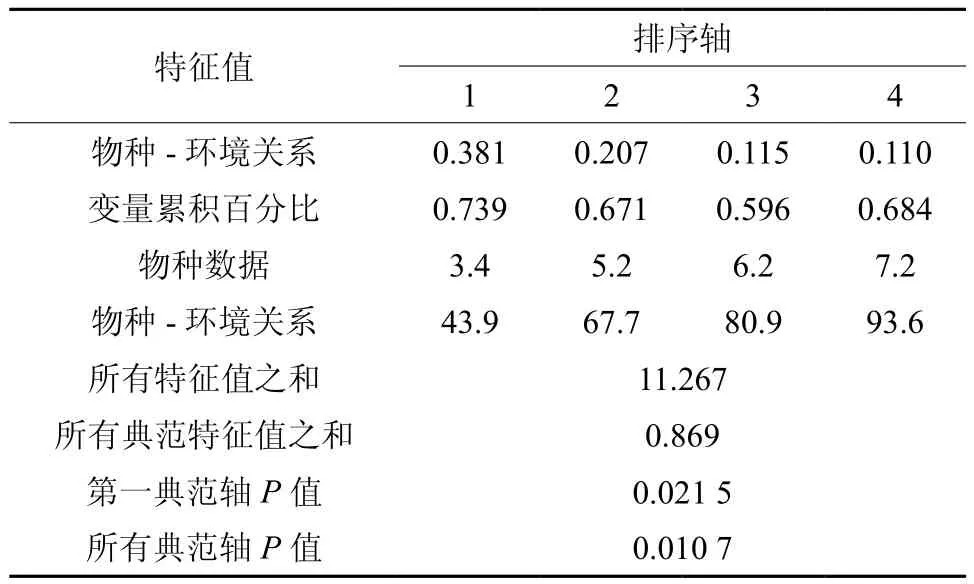
表3 各排序轴的加权相关矩阵Table 3Weighted correlation matrix of canonical correspondence analysis
立木冠层结构参数与排序轴之间的相关分析揭示,冠层结构与环境轴1和环境轴2的相关性较强(见表4),其中林冠开度与环境轴1的相关性最大(r= 0.685 1),叶面积指数与环境轴2的相关性最大(r= 0.891 8)。

表4 立木冠层结构参数与环境轴的相关系数Table 4Correlation coefficients of canopy structure parameters and environmental axis
冠层结构与立木的CCA二维排序图进一步表明立木分布与冠层结构各参数均有一定相关性(见图1)。图1中实心圆点代表立木种类,不同字母组成代表不同树种,带有箭头的连线代表冠层结构参数,箭头所指的方向代表该冠层结构参数的变化趋势,箭头长度代表相关性的大小,箭头连线与排序轴的夹角表示冠层结构参数与排序轴的相关性的大小,夹角越大说明相关性越大,物种点与冠层结构参数的箭头共同反映立木种类沿每一参数梯度方向的变化特征。从图1还可看出,立木在二、三象限分布相对较多且密集,对应叶面积指数较小、林冠开度较大的冠层结构以及林下光照较强的光照环境,而在一、四象限的分布较少且较稀疏,尤其在一象限分布最少最稀疏,对应叶面积指数较大、林冠开度较小的冠层结构以及林下光照较弱的光照环境。结合表4分析可知,叶面积指数和林冠开度与立木分布的关系较为密切。

图1 冠层结构与立木的典范对应分析Fig.1Canonical correspondence analysis between canopy structure factors and common standing trees
3.3 冠层结构与立木多度的关系
3.3.1 林冠开度与立木多度的关系
总的立木多度与活立木多度在林冠开度分组变量间的差异不显著(P > 0.05),但枯立木多度在林冠开度分组变量间的差异显著(P < 0.05)(见表5)。活立木多度在林冠开度10.1~15.5的范围内达到最大,此后逐渐减少,说明该范围内的光照、水分、温度等条件最适合该样地林木生长。枯立木多度在不同分组间的变化趋势跟活立木相似,在林冠开度10.1~15.5时多度最大,此后逐渐减少(见图2 A)。
3.3.2 叶面积指数与立木多度的关系
立木多度在叶面积指数分组变量间的差异不显著(P> 0.05)(见表5),在叶面积指数为2.01~3.00的范围内达到峰值(见图2 B),在两端的范围内明显减少,说明此范围内的叶面积指数最适合林木生长,可能是该范围内的叶面积指数有最适合林木生长的光照强度和水分条件。在叶面积指数再增大的过程中,林下光照减弱,影响植物的光合作用,进而影响植物生长。

表5 冠层结构与立木的Kruskal-Wallis分析Table 5Kruskal-Wallis analysis of standing trees with canopy structure parameters
3.3.3 林下光照与立木多度的关系
从Kruskal-Wallis分析的结果看,总的立木多度与活立木多度均在林下散射光的分组变量间差异显著(P< 0.05),枯立木多度在林下光照分组变量间差异均不显著(P> 0.05)(见表5)。由图2(D)可知,立木多度随着林下直射光的增强呈现明显的减少趋势。活立木与枯立木多度均在林下散射光2.01~3.00的范围内和在林下直射光≤3的范围内达到最大,枯立木多度变化趋势与活立木和总的立木多度变化趋势相同。活立木多度在林下总光照≤5和5.01~10.00的范围内均很大,在≥10.01的范围内明显减少(见图2 C, D,E),枯立木则在林下总光照5.01~10.00的范围内达到峰值,两端减少。

图2 立木多度对冠层结构分组变量的响应Fig.2Responses of standing trees to canopy grouping variables
4 结论与讨论
4.1 叶面积指数通过改变林下光照条件进而影响立木分布
叶片是植物重要的光合作用器官,冠层叶面积指数对群落的生物量有显著影响[17],而且与光合有效辐射(Photosynthetically Active Radiation,PAR)的关系密切[18],光合有效辐射影响着阳性植物和耐阴植物的分布。杨慧等[19]的研究结果表明,生物因素和环境因素是影响白桦Betula platyphylla种群空间分布格局的两个主要原因,在种间竞争不强的地区呈集群分布,在种间竞争激烈的地区呈随机分布。本研究表明,叶面积指数与典范对应分析环境轴2呈正相关,且相关性最大,说明叶面积指数对立木的分布影响较大。同时,叶面积指数与林下直射光、林下散射光呈显著负相关,叶面积指数越大,林下光照条件越差[19],说明叶面积指数主要通过影响林下光照条件影响植物分布。
4.2 林冠开度影响枯立木形成及多度
枯立木形成的原因包括对环境因子的竞争[21]、自然灾害[22]、地形因子[23]等。Sprintsin等[24]的研究表明,林冠开度和立地密度(单位样方内的林木数量)呈显著的线性正相关(R2= 0.96)。在林冠开度为53%时,立木密度达到最大值。营建母树林时,也需要伐除病虫危害株等扩大林冠开度[25],促进植株生长,说明林冠开度对林木的生长有一定影响。本研究结果也揭示,枯立木多度在林冠开度分组变量间的差异显著(P< 0.05),说明林冠开度对枯立木形成及多度有影响。林冠开度与林下光照条件直接相关,可能通过影响林下光照进而影响林木生长。
4.3 林下光照条件对立木多度影响显著
光照是影响植物生长的重要生态因子,本研究结果表明林下直射光和林下散射光对立木多度影响显著。立木多度随林下直射光的增强而减少,总的立木多度与活立木多度在林下散射光的分组间差异显著。有研究表明,提高散射光的比率能增加植物光能利用率[26],而植物通过调整光能利用率比改变生理结构更能促进光合生产[27],进一步说明散射光可以影响立木多度。林下直射光是通过林冠空隙直接入射到森林内部的太阳光线,是植物进行光合作用的重要光照来源,对植物的生长影响最大。本研究结果中,立木多度随着林下直射光增强而减少也证明了这一点。而且林下直射光强度与林冠开度直接相关,林冠开度越大,林下直射光越强,对植物生长越有利,本研究结果也表明立木多度随林冠开度和林下直射光的变化趋势大致相同。
综上所述,森林群落冠层结构与立木关系密切,叶面积指数和林冠开度通过影响林下光照条件进而影响立木,尤其是枯立木的形成和多度。林下直射光与林下散射光均对立木多度有影响,其中立木多度随林下直射光的变化趋势最明显,随着林下直射光增强而减少。不同的林冠开度有不同的林下光照条件和水分条件,从而影响不同种植物间或同种植物内部对光照、水分等生态因子的竞争作用,在研究森林群落种内种间竞争关系的过程中可将冠层结构作为影响因子纳入研究范围,作为一个研究竞争关系的新方向,探讨不同冠层结构条件下种内种间的竞争作用及强度。
[1]陈 厦,桑卫国. 暖温带地区3种森林群落叶面积指数和林冠开阔度的季节动态[J]. 植物生态学报, 2007, 31(3): 431-436.
[2]李汉强,邱治军,张宁南,等. 马占相思人工林的林冠截留效应[J]. 中南林业科技大学学报, 2013, 33(2): 86-90.
[3]Siles P, Vaast P, Dreyer E, et al. Rainfall partitioning into throughfall, stemflow and interception loss in a coffee (Coffea arabica L.) monoculture compared to an agroforestry system with Inga densif l ora[J]. Journal of Hydrology,2010,395(1-2):39-48.
[4]区余端,苏志尧. 粤北山地常绿阔叶林自然干扰后冠层结构与林下光照动态[J]. 生态学报, 2012, 32(18): 5637-5645.
[5]Beaudet M, Harvey B D, Messier C, et al. Managing understory light conditions in boreal mixedwoods through variation in the intensity and spatial pattern of harvest: A modelling approach[J].Forest Ecology and Management, 2011, 261(1): 84-94.
[6]郑国,李枢强. 森林冠层节肢动物多样性研究进展[J]. 动物分类学报, 2013, 38(1): 178-182.
[7]Ediriweera S, Singhakumara B M P, Ashton M S. Variation in canopy structure, light and soil nutrition across elevation of a Sri Lankan tropical rain forest[J]. Forest Ecology and Management,2008, 256: 1339-1349.
[8]Iglesias M R , Barchuk A, Grilli M P. Carbon storage, community structure and canopy cover: A comparison along a precipitation gradient[J]. Forest Ecology and Management, 2012, 265: 218-229.
[9]刘 娟,廖 康,安晓芹,等. 不同主枝开张角度杏树冠层微气候特征的差异[J]. 经济林研究,2012, 30(4): 82-86.
[10]Lhotka J M, Loewenstein E F. Indirect measures for characterizing light along a gradient of mixed-hardwood riparian forest canopy structures[J]. Forest Ecology and Management,2006, 226(1-3): 310-318.
[11]Astrup R, Larson B C. Regional variability of species-specific crown openness for aspen and spruce in western boreal Canada [J].Forest Ecology and Management, 2006, 228(1-3): 241-250.
[12]Ohsawa M, Shimokawa T. Extending the rotation period in larch plantations increases canopy heterogeneity and promotes species richness and abundance of native beetles: Implications for the conservation of biodiversity [J]. Biological Conservation, 2011,144(12): 3106-3116.
[13]Chianucci F, Cutini A. Estimation of canopy properties in deciduous forests with digital hemispherical and cover photography [J]. Agricultural and Forest Meteorology, 2013, 168:130-139.
[14]Lang M, Kuusk A, Mõttus M, et al. Canopy gap fraction estimation from digital hemispherical images using sky radiance models and a linear conversion method[J]. Agricultural and Forest Meteorology, 2010, 150(1): 20-29.
[15]张 璐,苏志尧,陈北光,等. 广东石坑崆森林群落优势种群生态位宽度沿海拔梯度的变化[J]. 林业科学研究, 2007,20(5): 598-603.
[16]Beaudet M, Messier C. Variation in canopy openness and light transmission following selection cutting in northern hardwood stands: an assessment based on hemispherical photographs[J].Agricultural and Forest Meteorology, 2002, 110(3): 217-228.
[17]莫丽芬,唐建维,刀祥生,等. 林分密度对小桐子生物量及其产量的影响[J]. 中南林业科技大学学报,2010, 30(4): 61-68.
[18]蚁伟民,张祝平,丁明懋,等. 鼎湖山格木群落的生物量和光能利用效率[J]. 生态学报,2000, 20(2): 397-403.
[19]杨 慧,娄安如,高益军,等. 北京东灵山地区白桦种群生活史特征与空间分布格局[J].植物生态学报,2007,31(2):272-282.[20]贾小容,苏志尧,区余端,等. 三种人工林分的冠层结构参数与林下光照条件[J]. 广西植物,2011, 31 (4): 473-478.
[21]张笑菁,赵秀海,康峰峰,等. 太岳山油松天然林林木的空间格局[J]. 生态学报, 2010, 30(18): 4821-4827.
[22]沈泽昊,王功芳,李道兴. 三峡大老岭山地常绿落叶阔叶混交林林隙干扰研究Ⅱ.林隙干扰的地形格局[J]. 植物生态学报,2001, 26(2): 149-156.
[23]刘妍妍,金光泽. 地形对小兴安岭阔叶红松(Pinus koraiensis)林粗木质残体分布的影响[J].生态学报,2009,29(3):1398-1407.
[24]Sprintsin M, Karnieli A, Sprintsin S, et al. Relationships between stand density and canopy structure in a dryland forest as estimated by ground-based measurements and multi-spectral spaceborne images[J]. Journal of Arid Environments, 2009,73(10): 955-962.
[25]佘远国,谈建文,李万德. 阳新县吴茱萸母树林营建技术[J].经济林研究, 2005, 23 (3): 62-64.
[26]Jenkins J P, Richardson A D, Braswell B H, et al. Refining light use efficiency calculations for a deciduous forest canopy using simultaneous tower-based carbon flux and radio-metric measurements[J]. Agricultural and Forest Meteorology, 2007,143: 64-79.
[27]Niinemets B. Photosynthesis and resource distribution through plant canopies[J]. Plant, Cell and Environment, 2007, 30: 1052-1071.
Relationship between canopy structure factors and abundance of standing trees in Nanling mountain forest communities
JING Xiao-li, DU Wei-jing, ZHANG Lu, SU Zhi-yao
(College of Forestry, South China Agricultural University, Guangzhou 510642, Guangdong, China)
Canopy is the most direct and active interface layer between forest and external environment, which affects physical and biotic environment of forest. By using Hemispherical Photography, Canonical Correspondence Analysis (CCA) and Kruskal-Wallis methods,the relations among the distribution and abundance of standing trees and forest canopy structure were quantitatively studied. The results indicated that (1) There were 47 families, 81 genera and 143 species based on the 10000m2sampling investigations and most of the dead standing trees derived from over-story big old-aged trees; (2) the results of CCA ordination indicated that all of the canopy structure parameters had a certain correlation with distribution of standing trees, and the leaf area index (LAI) and canopy openness (CO) had a signif i cant correlation with distribution of standing trees; (3) The abundance of standing trees and abundance of living standing trees had a signif i cant difference among Transmitted Diffused Light (TDif) groups (P < 0.05) and abundance of snags had a signif i cant difference among canopy openness groups (P < 0.05), besides, the abundance of standing trees decreased with the enhancement of Transmitted Direct Light (TDir). Moreover, LAI and CO affected under-story light conditions, thereby to the stumpage, especially the formation and abundance degrees of the withered tree.
mountain forest communities; canopy structure; stumpage abundance; standing tree; dead standing tree; leaf area index (LAI);canopy openness (CO)
S718.5
A
1673-923X(2014)05-0059-07
2013-08-10
广东省林业科技创新专项资金项目(2011KJCX031-02)
敬小丽(1989-),女,四川遂宁人,硕士研究生,研究方向:森林生态学;E-mail:jingxiaoli0930@163.com
张璐(1973-),女,贵州习水人,副教授,博士,研究方向:森林生态学;E-mail:zhanglu@scau.edu.cn
[本文编校:谢荣秀]
