伊犁农田两种狗尾草无性系种群的数量性状特征
2014-12-24贾风勤努尔买买提纳森巴特赖卓彦
贾风勤,张 娜,努尔买买提,纳森巴特,赖卓彦
(1.新疆自治区普通高校天然产物化学与应用重点实验室 资源与生态研究所 伊犁师范学院化学与生物科学学院,新疆 伊宁835000;2.台北市立动物园,台湾 台北11656)
研究植物无性系种群构件数量特征变化,可以了解不同构件对无性系生长、繁殖和物质生产的贡献等。生物量结构是判断不同构件组分对无性系生产与发育贡献大小的依据。生物量配置是植物生理整合作用对生境异质环境或微生境的反应[1]。狗尾草(Setariaviridis)和金色狗尾草(S.glauca)为禾本科狗尾草属一年生无性系草本植物,均具有耐旱[2]、耐盐碱[2]等生物生态学特性,能在短时间内形成地表优势种,被用于退耕地[4]、矿区[5]、公路[6]等的生态恢复和防护,既是一种优良牧草,却也是较难防除的农田杂草[7-8]。在新疆伊犁,当年的休耕农田中常可形成两种狗尾草同时存在的群落和层片。目前,有关两种狗尾草种子萌发[9]、根叶生长[10-11]的生物学特性的研究已见报道,鲜见有关两种植物的无性系种群数量性状特征及其生长策略报道。本研究将通过对两种狗尾草种群构件诸数量性状内在规律的研究,深入了解其无性系的生态适应机制,为合理开发和利用狗尾草属植物资源提供科学理论依据。
1 材料与方法
1.1 研究区自然概况
研究区位于新疆伊犁霍城县四爪陆龟自然保护区,周边为灌溉农耕区。地理位置44°06′-44°18′N,80°48′-81°58′E。海拔650~1 100m。属于温带半干旱气候,无霜期165d,年均气温9.1℃,年均降水219mm。保护区与周边绿洲交错带主要以人工种植杨属(Populus)、榆属(Ulmus)树种为主,林下植被以角果毛茛(Ceratocephalustesticulatus)、苦豆子(Sophoraaolpecuroides)单优种群为主,常以斑块状镶嵌于林地中;保护区植被主要由角果毛茛、囊果苔草(Carexphysodes)、毛穗旱麦草(Eremopyrumdistans)、苦豆子、蒿(Artemisiasp.)植物组成[12-13]。灌溉农耕区农作物以玉米(Zea mays)、水稻(Oryzasativa)、小麦(Triticumaestivum)为主,休耕当年裸地则多以狗尾草和金色狗尾草形成的优势群落所占据,伴生种则以荠菜(Capsellaburas-pastoris)、圆叶锦葵(Malvarotundifolia)、田旋花(Convolvulusarvensis)、灰绿藜(Chenopodiumglaucum)为主。
1.2 材料
研究材料狗尾草和金色狗尾草在伊犁地区均在4-5月出苗,6―9月为花果期,圆锥花序,种子成熟后随风散布或自行脱落。
1.3 试验方法
研究样地选择弃耕一年的农田,2012年8月于狗尾草和金色狗尾草籽实成熟期,在两者形成优势群落的休耕农田中,分别选择丛径大小不等、籽粒尚未脱落或极少脱落的狗尾草和金色狗尾草分蘖丛各38个。为尽量减少籽实损失,将每个分蘖丛生殖株的穗头先齐穗颈处剪下,置于纸袋中,穗及对应生殖株为同一编号,并标示相应分蘖丛编号。其余地上部分连同地下根系一并完整挖出,编号装入塑料袋带回室内。将各分蘖丛冲洗干净后分置,将丛内所有分蘖株齐根剪下。计数每个分蘖丛的总分蘖株(总分蘖)和生殖分蘖株(生殖分蘖)数量,测定各分蘖株长度并将丛内分蘖株叶、鞘、茎、穗等构件分置,其中总株高为各分蘖株株高总和,在烘箱内75℃烘干72h至质量恒定不变后称量。

对分株构件数量性状与总分蘖数和总生物量间定量关系的确定,是在拟合的线性、指数和幂3种函数中,选用相关性最高的作为关系模型。
1.4 数据处理
用SPSS 17.0对总分蘖数、生殖分蘖数、总株高与总生物量以及茎、叶、鞘与总蘖数间关系进行相关性分析。用单因素方差分析(One-way ANOVA)比较种间茎、叶、鞘和穗生物量、生殖分蘖数、总分蘖数、总株高、总生物量的差异显著性。用Excel软件绘图。
2 结果
2.1 无性系种群数量特征
测定结果显示,两种狗尾草无性系种群内所有构件数量性状的变异系数均超过50%,最高达123.48%,最小为55.32%;茎、叶、鞘生物量在两个狗尾草种群内均呈现依次降低趋势,在种间无显著差异(P>0.05)。穗生物量、生殖分蘖数、总分蘖数和总株高均表现为狗尾草显著高于金色狗尾草(P<0.05)(表1)。
在单项指标(茎、叶和鞘生物量及生殖蘖数)上,狗尾草无性系在茎生物量和生殖分蘖数量上表现出较大变异,变异系数分别为107.17%和107.03%;金色狗尾草以叶生物量变异最大,为84.43%。总体
指标上(总分蘖数、总生物量、总株高),两种狗尾草均以总生物量变异最大。狗尾草无性系所有数量特征的变异系数均显著高于金色狗尾草(P<0.05)。

表1 狗尾草和金色狗尾草无性系数量性状Table 1 Quantitative characteristics of studied clone populations of Setaria viridis and S .glauca
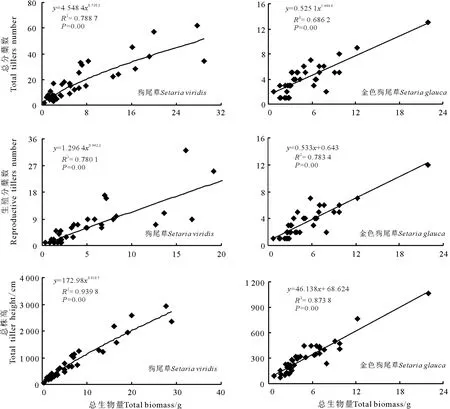
图1 两种狗尾草无性系分蘖数量、生殖蘖数量、总株高与总生物量关系拟合曲线Fig.1 Relationships between tiller number,reproductive tillers,total tiller height and total biomass in two Setaria species
2.2 无性系种群总分蘖数、生殖分蘖数和总株高与总生物量间关系
分析结果显示,两种狗尾草无性系总分蘖数、生殖分蘖数和总株高与总生物量间均呈显著正相关性(P<0.05)。其中,狗尾草无性系种群中总分蘖数、生殖分蘖数、总株高与总生物量间相关系数分别为0.888、0.883和0.969,且拟合度最好的关系模型为幂函数正相关关系;金色狗尾草中相关系数分别为0.828、0.885和0.935,拟合度最好的关系模型为线性正相关关系。狗尾草无性系种群中以生殖分蘖数增长速度为快,总分蘖数增长为慢;金色狗尾草则以总株高增长为快,而生殖分蘖数和总分蘖数则表现为平行增长(图1)。
2.3 两种狗尾草无性系构件生物量与总分蘖数间关系
统计分析表明,狗尾草属两物种无性系的茎、叶和鞘生物量与总分蘖数间均呈显著正相关关系(P<0.05),但在两物种间表现出不同增长关系。其中,狗尾草无性系茎、叶、鞘生物量与总蘖数间拟合程度最高的关系模型均为幂函数,相关系数分别为0.854、0.890和0.904;金色狗尾草表现为线性增长关系,相关系数r值0.811、0.795和0.839(图2)。
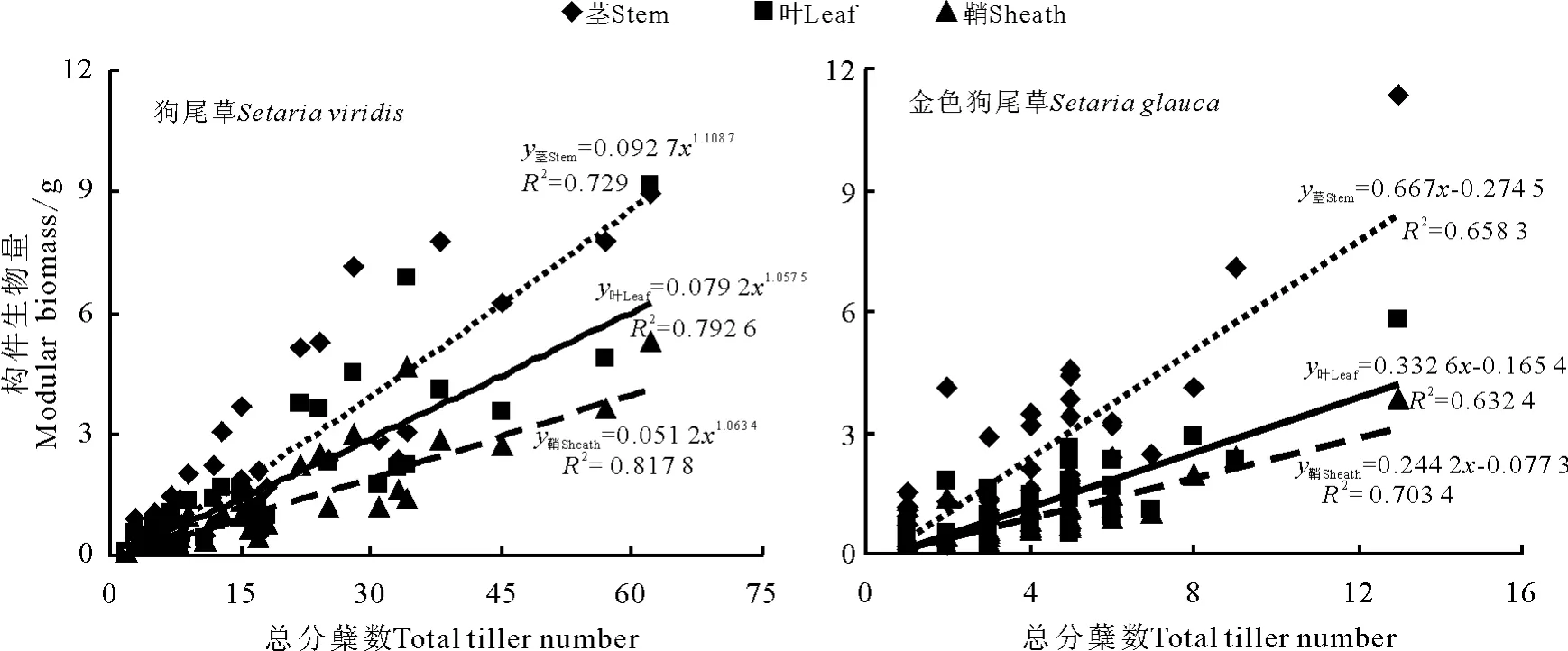
图2 两种狗尾草无性系构件生物量与总蘖数间拟合曲线Fig.2 The curve fitting of relationship between modular biomass and total tillers number of two Setaria species
2.4 两种狗尾草无性系种群生物量分配
研究结果显示,狗尾草和金色狗尾草种群构件生物量分配均呈营养构件生物量分配所占比例高于生殖构件,营养构件生物量分配为47.74%~18.18%,生殖分配为10.82%~15.04%。两个狗尾草生物量分配均呈现茎>叶>鞘>穗。两种狗尾草无性系种群间除鞘生物量分配外,茎、叶和穗生物量分配间均存在显著差异(P<0.05)(图3)。
3 讨论与结论
表型可塑性是指一个基因型受到环境影响时的表型表达,或在异质环境中所能产生的不同表型,它被认为是植物在可变环境中最大适合度的功能反映[14]。本研究中,如果用变异系数来衡量狗尾草和金色狗尾草无性系种群数量特征对随机环境因子的反映即表型可塑性[15-16],则狗尾草无性系种群较金色狗尾草呈现出更大的表型可塑性。狗尾草无性系除总分蘖数和总株高外,其他数量特征变异系数均超过100%;金色狗尾草无性系变异系数最大的为叶生物量,其值为84.43%,这表明环境因子的变化导致狗尾草无性系数量性状呈现较大变异,金色狗尾草则相对稳定,也表明两者均可通过种群表型可塑性的较大变异来适应不同微环境的策略。
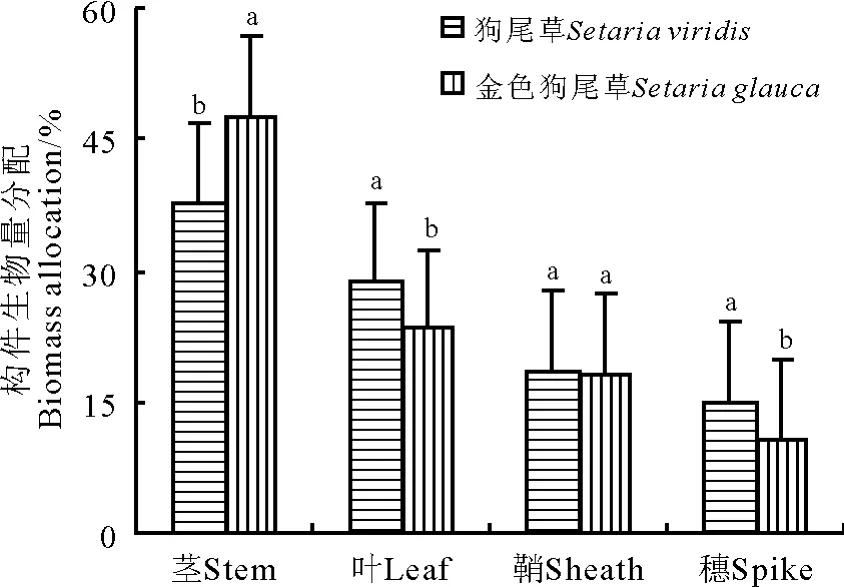
图3 狗尾草和金色狗尾草无性系种群构件生物量分配间关系Fig.3 Allocation relationship between different modulars of two Setaria species
如果把植物组分间生长规律看作是由遗传因子控制的,用拟合方程的确定系数R2来估计,随机环境影响用1-R2来估计[16],狗尾草无性系种群在总分蘖数、生殖分蘖数、总株高与总生物量间关系及组分生物量与总分蘖数生长关系中,分别有78.01%~93.98%和72.90%~81.78%是受遗传因子控制的;金色狗尾草则降低至68.62%~87.38%和63.24%~70.34%,表明狗尾草无性系种群数量特征受遗传因子作用更大。在受遗传因子控制的同时,两种狗尾草也通过各构件数量特征的改变来增加资源获得以占据多样生境。狗尾草无性系种群通过较大幅度的分蘖数量变化(2~62个,变异系数为84.97%)来最大程度获得可利用资源,并通过较小且稳定的平均株高(平均值为44.97,变异系数为23.88%)减少对可利用资源的消耗;金色狗尾草则通过变化幅度小的分蘖数量(1~13个,变异系数为59.38%)和稳定、高大的分蘖株(平均值为8.64,变异系数为29.62%)与狗尾草成为共同优势种。
狗尾草属两种无性系种群数量特征均具有较大的表型可塑性。狗尾草无性系种群的总蘖数、生殖蘖数和总株高与总生物量关系,茎、叶、鞘生物量与总蘖数的关系均表现为幂函数正相关,而金色狗尾草则呈线性增长。生物量分配在两物种间除鞘生物量分配外,茎、叶和穗生物量分配间均存在显著差异。
[1]Stuart Chapin III F,Bloom A J,Field Ch B,Waring R H.Plant responses to multiple environmental factors[J].BioScience,1987,37:49-57.
[2]张继义,赵哈林.短期极端干旱事件干扰后退化沙质草地群落恢复力稳定性的测度与比较[J].生态学报,2011,319(20),6060-6071.
[3]Guo R,Zhou J,Hao W P,Gong D Z,Zhong X L,Gu F X,Liu Q,Xia X,Tian J N,Li H R.Germination,growth,photosynthesis and ionic balance inSetariaviridisseedlings subjected to saline and alkaline stress[J].Canadian Journal of Plant Science,2011,91:1077-1088.
[4]王星,李占斌,李鹏.陕西省丹汉江流域退耕地恢复过程中的植被演替[J].应用生态学报,2012,23(2):347-356.
[5]闫德民,赵方莹,孙建新.铁矿采矿迹地不同恢复年限的植被特征[J].应用生态学杂志,2013,32(1):1-6.
[6]李妮,陈其兵,谭昌明.高速公路边坡乡土灌木建植模式水土保持效益[J].水土保持学报,2012,26(1):67-71.
[7]Friesen G,Shebeski L H.Economic losses caused by weed competition in Manitoba grain fields.Ⅰ.Weed species,their relative abundance and their effect on crop yields[J].Canadian Journal of Plant Science,1960,40:457-467.
[8]Wang R L,Dekker J.Weedy adaptation in Setaria spp.Ⅲ.Variation in herbicide resistance inSetariaspp.[J].Pesticide Biochemistry and Physiology,1995,51:99-116.
[9]陶敏,包达川,蒋明喜.三峡区9种植物种子萌发特性及其在植被恢复中的意义[J].生态学报,2011,31(4):0906-0913.
[10]毛伟,李玉霖,赵学勇,黄迎新,王少昆.科尔沁沙地灌丛内外草本植物狗尾草叶性状的比较研究[J].草地学报,2009,18(6):144-150.
[11]黄刚,赵学勇,苏延桂.科尔沁沙地3种草本植物根系生长动态[J].植物生态学报,2007,31(6):1161-1167.
[12]贾风勤,金希堂,李紫薇,张维.不同生境囊果苔草种群分蘖株的数量特征[J].西北农林科技大学学报(自然科学版),2011,39(4):205-210.
[13]贾风勤,李紫薇,金希堂,张维.新疆四爪陆龟保护区植物组成及其结构分析[J].干旱区研究,2011,28(6):1020-1024.
[14]朱志红,刘建秀,王孝安.克隆植物的表型可塑性与等级选择[J].植物生态学报,2007,31(4):588-598.
[15]李仲芳,王刚.种内竞争对一年生植物高生长与生物量关系的影响[J].兰州大学学报(自然科学版),2001,38(2):141-146.
[16]杨允菲,李建东.松嫩平原不同生境芦苇种群分株的生物量分配与生长分析[J].应用生物学报,2003,14(1):30-34.
