粳稻穗角、着粒密度及每穗颖花数的遗传
2014-12-23刘金波王宝祥刘晓丽方兆伟卢百关周振玲陈庭木秦德荣徐大勇
刘金波, 王宝祥, 刘晓丽, 李 健, 杨 波, 方兆伟, 卢百关, 刘 艳,迟 铭, 周振玲, 陈庭木, 秦德荣, 徐大勇
(1.江苏徐淮地区连云港农业科学研究所,江苏 连云港222000;2.南京农业大学作物遗传与种质创新国家重点实验室,江苏南京210095)
粳稻直立穗型是继矮化和理想株型后水稻高产株型的又一重要形态[1]。直立穗粳稻品种的选育及推广,促进了粳稻产量的进一步提高[2]。粳稻直立穗品种着粒密度大,弯曲穗品种着粒密度小。张文忠等研究发现产量和着粒密度高低顺序是直立穗品种>半直立穗品种>弯曲穗品种[2]。徐大勇等认为直立穗高产水稻品种表现出一次和二次枝梗数增加、每穗颖花数和实粒数上升、着粒密度增大的特点[3]。董丹等研究认为直立穗品种库容量大,干物质生产能力强、流畅,茎秆的干物质输出率和转换率高,而且穗长变短,叶型直立,进而使群体的环境条件优越,产量较高[4]。目前,生产上粳稻品种以穗型半直立为主。因此,研究穗型和着粒密度及每穗颖花数的遗传,对于水稻杂种优势利用和提高粳稻产量具有重要的意义。洪德林等提出水稻“生态配组”、“水涨船高”的育种策略[5]。李峰[6]、张书标等[7]通过水稻直立密穗型等位突变体(Dense and erect panicle2,dep2)的研究,发现dep2 突变体株叶形态变好,抗倒伏能力增强,千粒重下降,但突变体产量并未降低。因此,可利用直立穗品种高产、抗倒、株叶形态好等优良特性改良品种,提高粳稻产量。
王伯伦等以穗弯曲度小于30°为标准,分析认为直立穗受1 对隐性基因控制[8]。刘金波、陈献功、牛付安等也曾对粳稻的穗型做过研究[9-11],但亲本间差异不大。本研究利用株高和穗部性状差异较大的亲本杂交产生的丰富分离群体,采用主基因+多基因模型,研究直立穗型及相关性状的遗传,为粳稻品种的选育提供理论依据。
1 材料与方法
1.1 供试材料
试验材料有直立穗品系3012(P1)、弯曲穗品种万特大粒(P2)以及P1×P2产生的F1和F2。直立穗品种3012(P1)为连云港市农业科学院选育的籼粳交中间材料,株高74.5 cm,平均穗角为17.1°,主茎穗着粒密度(简称着粒密度,下同)为1 cm 30.36粒,主茎穗每穗颖花数(简称每穗颖花数,下同)为447.8 粒,千粒质量17.4 g;弯曲穗品种万特大粒(P2),由连云港市农业科学院从连云港市黄淮作物育种所引入,株高135.9 cm,平均穗角135.3°,着粒密度为1 cm 7.86 粒,每穗颖花数为157.9 粒,千粒质量42.3 g。双亲差异显著。两者杂交产生的F1代穗大粒多,米质好,综合了两亲本的高产优质特性。
1.2 试验方法
1.2.1 田间种植与考种方法 2010 年正季将试验材料P1、P2、F1和F2种植于连云港市农业科学院玉带河试验基地。5 月4 日浸种,5 月7 日播种,6 月23 日移栽。亲本和F1,每材料种植5 行,每行10株。F2种植150 株。株行距13.3 cm×16.7 cm。单本种植,常规栽培管理。成熟时,测量穗角(穗尖到穗颈节间的连线与茎秆的延长线所形成的夹角)[8],同时每株取主茎穗考察每穗颖花数和着粒密度性状。着粒密度是指1 cm 稻穗上颖花数。亲本和F1均调查10 株,F2调查120 株。
1.2.2 数据分析方法 利用南京农业大学章元明教授提供的植物数量性状主基因+多基因混合遗传模型软件,分析性状的遗传模型,并根据模型估计主基因和多基因效应值及其方差等遗传参数[12]。利用SPSS Statistics 对穗角和着粒密度及每穗颖花数3个性状进行相关性分析。
2 结果与分析
2.1 3012 ×万特大粒组合的P1、P2、F1和F2 4 个世代的穗角表现
从表1 可以看出,3012 平均穗角为(17.10±2.02)°,万特大粒平均穗角为(135.30±2.15)°,双亲差异显著。F1平均穗角为(114.40±1.71)°,介于双亲之间,偏向于高亲。F2平均穗角为(102.79±33.05)°,偏向于高亲,而且出现超高亲个体。F2群体呈连续分布,有3 个峰,显示有效应较大的主基因存在。
2.2 3012 ×万特大粒组合的P1、P2、F1和F2 4 个世代的着粒密度表现
从表2 可以看出,3012 平均着粒密度为1 cm(30.36 ±1.02)粒,万特大粒平均着粒密度为1 cm(7.86 ±0.56)粒,双亲差异显著。F1平均着粒密度为1 cm (21.65 ±0.77)粒,介于双亲之间,偏向于高亲。F2平均着粒密度为1 cm (12.64 ±4.39)粒,偏向于高亲。F2群体呈连续分布,有2 个峰,显示有效应较大的主基因存在。

表1 3012 ×万特大粒组合的P1、P2、F1和F2穗角次数分布Table 1 Frequency distributions of panicle angle in P1,P2,F1 and F2 generations derived from the cross of 3012 and Wantedali
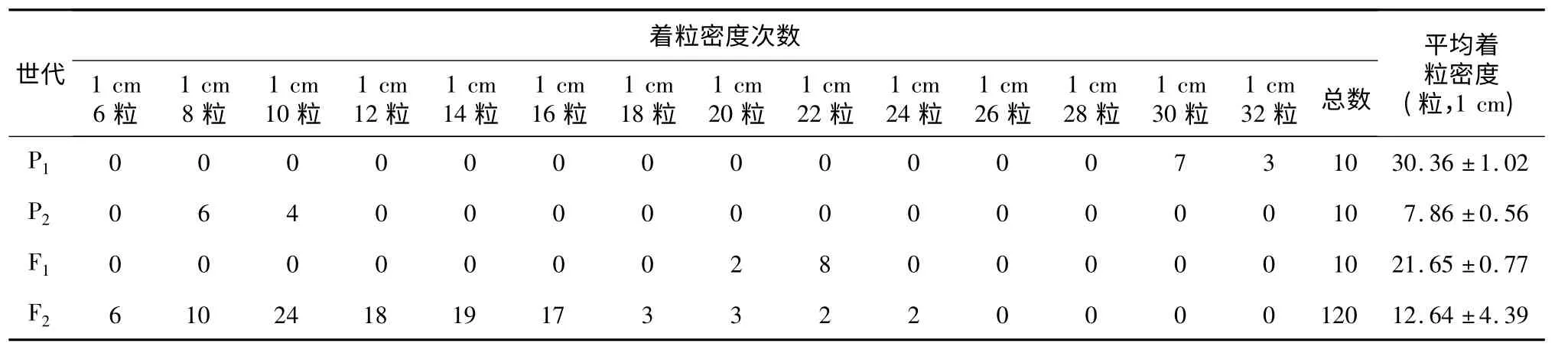
表2 3012 ×万特大粒组合的P1、P2、F1和F2着粒密度次数分布Table 2 Frequency distributions of seed setting density in P1,P2,F1 and F2 generations derived from the cross of 3012 and Wantedali
2.3 3012 ×万特大粒组合的P1、P2、F1和F2 4 个世代的每穗颖花数表现
从表3 可以看出,3012 平均每穗颖花数为(447.80 ±8.11)粒,万特大粒平均每穗颖花数为(157.90 ±4.91)粒,双亲差异显著。F1平均每穗颖花数为(441.50 ±7.13)粒,介于双亲之间,偏向于高亲。F2平均每穗颖花数为(304.33 ±109.01)粒,偏向于高亲,而且出现超高亲个体。F2群体呈连续分布,有2 个峰,显示有效应较大的主基因存在。
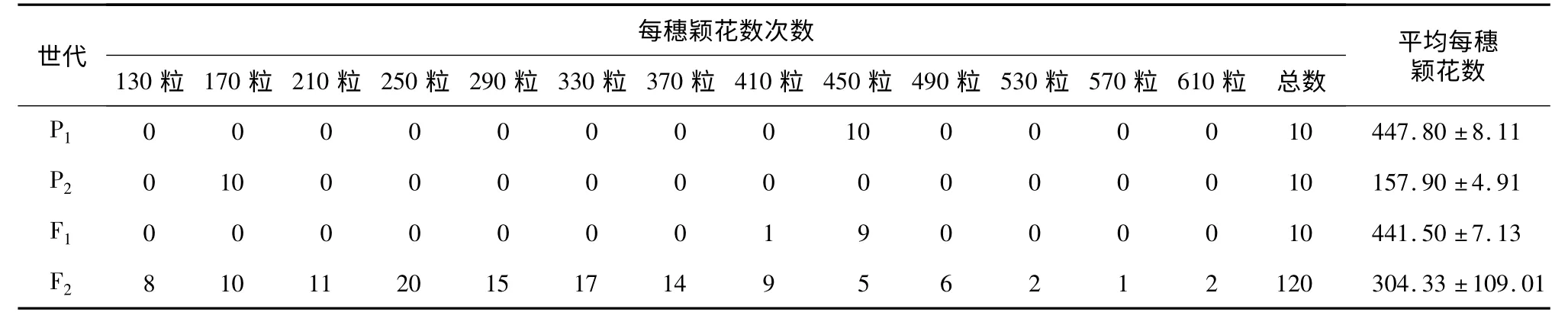
表3 3012 ×万特大粒组合的P1、P2、F1和F2每穗颖花数次数分布Table 3 Frequency distributions of spikelets per panicle in P1,P2,F1 and F2 generations derived from the cross of 3012 and Wantedali
2.4 遗传参数的估计
经遗传模型极大似然值和AIC值(赤池信息准则,Akaike’s information crilerion)比较,似然比检验(LRT)结果(表4)显示,3012 ×万特大粒组合穗角和每穗颖花数的最适遗传模型均为D-0 模型,即该性状表现为1 对加性-显性主基因+加性-显性-上位性多基因混合遗传。着粒密度的最适遗传模型为E-0 模型,即该性状表现为2 对加性-显性-上位性主基因+加性-显性-上位性多基因混合遗传。
根据遗传模型,可以看出穗角存在3 个成分分布,分布比例为1∶ 2∶ 1,显示3 个峰,拟合后显示为2 个峰;同样,着粒密度存在9 个成分分布,分布比例为1∶ 2∶ 1∶ 2∶ 5∶ 3∶ 1∶ 3∶ 1,显示9 个峰,拟合后显示3 个峰;每穗颖花数存在3 个成分分布,分布比例为2∶ 1∶ 1,显示3 个峰,拟合后显示1 个峰(图1)。

表4 穗角、着粒密度和每穗颖花数遗传模型的适合性检验Table 4 Tests for goodness-of-fit of genetic model for panicle angle,seed setting density and spikelets per panicle
根据遗传模型中成分分布及相应的均值,估计模型中的一阶参数和二阶参数(表5)。由表5 可知,在3012 ×万特大粒组合中控制穗角的1 对主基因的显性效应与加性效应基本上相当,基因显性度为-0.99,说明控制穗角的1 对主基因加性效应和显性效应都同等重要,穗型弯曲对直立呈部分显性。控制穗角的主基因的遗传率为56.62%,多基因的遗传率为43.02%。说明穗角以主基因控制为主,多基因效应也较大,环境效应小,占总表现型方差的0.36%。

图1 穗角、着粒密度和每穗颖花数次数分布、拟合分布和成分分布Fig.1 Frequency distribution,fitted mixed distribution and its component distribution of panicle angle,seed setting density and spikelets of per panicle
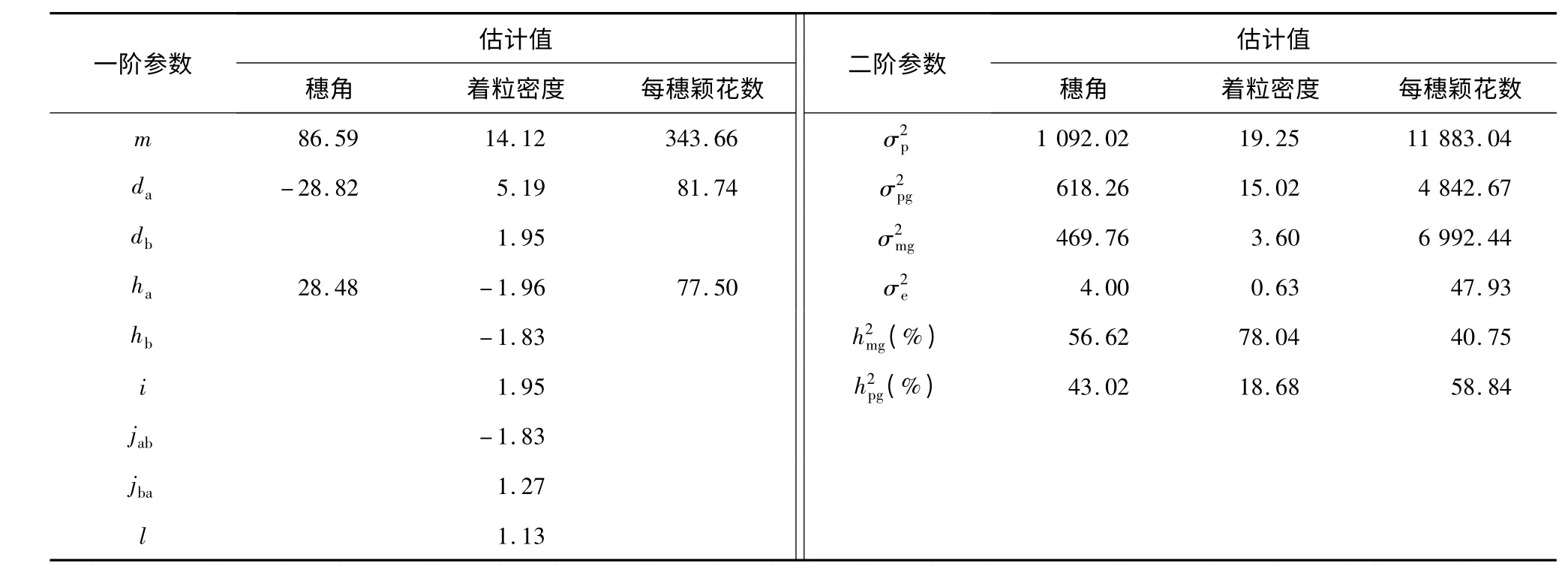
表5 3012 ×万特大粒组合的穗角和着粒密度及每穗颖花数的遗传参数估计值Table 5 The estimates of genetic parameters of panicle angle,seed setting density and spikelets per panicl e in the cross of 3012 ×Wantedali
同样,控制着粒密度的2 对主基因里,第2 对主基因的加性效应值相当于第1 对主基因加性效应值的37.58%。2 对主基因显性度分别为-0.38 和-0.94,说明控制着粒密度的2 对主基因都以加性效应为主,都呈负向显性;着粒密度密对疏呈部分显性。该组合中2 对主基因间互作效应基本相同。在基因效应中,加性效应最大,占41.73%;互作效应其次,为36.12%;显性效应较小,占22.15%。加性效应与加性×加性互作效应在总效应中比重很大,为53.13%,显性效应以及显性与其他效应的互作效应次之,为46.87%。控制着粒密度的主基因遗传率为78.04%,多基因遗传率为18.68%。说明着粒密度以主基因控制为主。环境效应小,占总表现型方差的3.28%。
控制每穗颖花数的1 对主基因的显性度为0.95,说明控制每穗颖花数的1 对主基因的加性效应和显性效应都同等重要;每穗颖花数多对少呈部分显性。控制每穗颖花数的主基因遗传率为40.75%,多基因遗传率为58.84%,说明每穗颖花数以多基因控制为主,主基因作用也较大,环境效应小,占总表现型方差的0.41%。
3 讨论
本研究采用植物数量性状主基因+多基因混合遗传模型分析主基因和多基因效应值及其方差等遗传参数,能够清晰地认识性状的遗传规律,然后再进行图形分布的拟合,可以真实地再现群体的分布,避免人为划分标准造成分析的偏差[9]。
水稻的杂种优势主要表现在每穗颖花数的增加[13]。而稻穗的直立与否,影响水稻群体的通风透光,从而影响水稻的产量。了解穗型和每穗颖花数的遗传,可以有针对性地进行亲本改良和水稻高产育种。本研究发现穗角和每穗颖花数性状均受1 对主基因+多基因共同控制。而笔者、陈献功、牛付安、江建华等研究认为穗角和每穗颖花数性状均受2 对主基因+多基因共同控制[9-11,14]。这可能与亲本选择有关。后者研究的母本均为纯粳稻品种,平均每穗粒数少、粒质量较小。而本研究的母本为籼粳交偏粳类型,母本每穗粒数多、粒小、穗型直立,而父本粒大、穗长、穗型弯曲。穗型的遗传与张书标等[7]、王伯伦等[8]、朱立宏等[15]、朱克明等[16]的研究结果相近,穗型直立受1 个隐性单基因控制。刘金波等[9]和江建华等[14]研究发现每穗颖花数的遗传受环境因素影响较大,而本研究发现每穗颖花数的遗传受环境因素影响很小,这主要是由本研究亲本每穗颖花数差异较大而造成的。
孟维韧等根据水稻着粒密度将试材分为散穗型、半散穗型、半紧穗型和紧穗型4 种类型,各类型间稻谷产量差异显著,其中半散穗型和半紧穗型品种产量较高[17]。范桂枝等以粳稻Asominori 与籼稻IR24 所衍生的染色体片段置换系(CSSLs)为材料,在水稻第4 染色体上发现1 个控制水稻着粒密度的QTL,贡献率为21.17%,加性效应为-0.49[18]。黎凌等利用水稻密穗突变体纯合单株A98 与中花1号杂交,发现A989 突变体中密穗性状是由单一基因控制,该密穗突变体表现晚开花、穗二级枝梗和小花数增加以及包颈现象[19]。本研究结果表明,着粒密度性状受为2 对加性-显性-上位性主基因+加性-显性-上位性多基因控制,以主基因作用为主,主基因的遗传率为78.04%。
对穗角、着粒密度和每穗颖花数3 个性状间进行相关性分析,发现穗角和着粒密度间存在极显著负相关,相关系数为-0.24;穗角和每穗颖花数间没有显著性正相关,相关系数为0.01;着粒密度和每穗颖花数间存在极显著正相关性,相关系数为0.87。这表明,直立穗品种穗角小,着粒密度大,每穗颖花数较多;而弯曲穗品种穗角大,着粒密度小,每穗颖花数较少,这与刘金波等的研究结果[9]一致。因此,穗角相对小(半直立),着粒密度相对大,可能会产生每穗颖花数超亲的现象,这也与刘金波等的研究结果[9]一致。说明,直立穗品种和弯曲穗品种杂交产生的F1,穗型半直立,每穗颖花数却很多,表现出强杂种优势。从田间调查结果看,杂种F1在有效穗数性状上也出现超亲优势。
从育种策略看,对主基因遗传为主的性状,可采用杂交、回交的育种策略;多基因遗传为主的性状,应采用聚合杂交、集团选择的育种策略。从育种选择效果看,以加性效应控制为主的性状,可在分离早代进行选择;以显性效应控制为主的性状,在分离晚代进行选择。从本研究结果可以看出,从育种策略和选择效果上,穗型和着粒密度性状适用于前者,每穗颖花数性状适用于后者。
直立穗品种具有高产的潜力,株型紧凑,耐肥抗倒。直立穗基因在粳稻品种中已经得到广泛应用,能否在籼稻中得到利用?籼稻亲本叶片长、植株高、穗长、穗型弯曲,如果与直立穗品种杂交,能否促进杂交籼稻优势的利用?这都需要进一步研究。
[1] 陈温福,徐正进,张文忠,等.水稻超高产育种生理基础[M].辽阳:辽宁科学技术出版社,1995.
[2] 张文忠,徐正进,张步龙,等.直立穗型品种演进状况分析[J].沈阳农业大学学报,2000,33(3):161-166.
[3] 徐大勇,朱庆森.直立穗型粳稻品种农艺特性及育种研究进展[J].植物遗传资源学报,2003,4(3):350-354.
[4] 董 丹,陈书强,刘柏林,等.直立穗基因对水稻源、库、流有关性状影响的研究[J].辽宁农业科学,2009(1):1-6.
[5] 洪德林,杨开晴,潘恩飞.粳稻不同生态类型间F1的杂种优势及其亲本的配合力分析[J].中国水稻科学,2002,16(3):216-220.
[6] 李 峰.水稻直立密穗突变体dep2 的分离与DEP2基因功能的鉴定[D].北京:中国科学院研究生院,2010.
[7] 张书标,马洪丽,黄荣华,等.籼稻直立穗突变体的培育、鉴定及其突变性状的遗传分析[J].核农学报,2007,21(3):209-211,241.
[8] 王伯伦,董玉慧,王 术.水稻半矮生与穗直立性状遗传规律研究[J].沈阳农业大学,1997,28(2):83-87.
[9] 刘金波,洪德林.粳稻穗角和每穗颖花数的遗传分析[J].中围水稻科学,2005,19(3):223-230.
[10] 陈献功,刘金波,洪德林.粳稻直立穗与弯曲穗3 个杂交组合6个世代穗角和每穗颖花数的遗传分析[J].作物学报,2006,32(8):1143-1150.
[11] 牛付安,刘 健,郭 媛,等.4 个环境下稳定表达的控制粳稻穗角性状的新位点[J].中国水稻科学,2012,26(4):409-416.
[12] 盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003.
[13] VIRMANI S S ,AQUINO R C ,KHUSH G S.Heterosis breeding in rice(Oryza salivaL.)[J].Theor Appl Genet,1982,63(4):373-380.
[14] 江建华,张启武,洪德林.粳稻穗部性状遗传分析[J].植物学报,2010,45(2):182-188.
[15] 朱立宏,顾铭洪.水稻落粒性的遗传[J].遗传,1979,1(4):17-19.
[16] 朱克明.水稻直立穗基因EP2的克隆与功能分析[D].扬州:扬州大学,2009.
[17] 孟维韧,王伯伦,吕 军,等.水稻着粒密度对产量和品质形成的影响[J].华中农业大学学报,2009,28(3):262-267.
[18] 范桂枝,蔡庆生,王春明,等.高CO2浓度下水稻穗部性状的QTL 分析[J].中国农业科学,2008,41(8):2227-2234.
[19] 黎 凌,时振英,沈革志,等.水稻密穗突变体A989 突变基因克隆和转基因植株分析[J].作物学报,2010,36(6):887-894.
