持续高温应激对肥育猪生产性能、胴体性状、背最长肌营养物质含量及肌纤维特性的影响
2014-12-20杨培歌冯跃进顾宪红杨春合曹正辉
杨培歌 冯跃进 郝 月 顾宪红 杨春合 曹正辉
(中国农业科学院北京畜牧兽医研究所,动物营养学国家重点实验室,北京 100193)
高温环境近年来已被认为是制约畜牧业发展的主要环境因子之一[1-2]。随着全球性气候变暖,我国夏季的平均气温越来越高。高温这一应激因子严重地影响着畜禽的生长发育、生产、繁殖及肉品质等,给全球畜牧业造成很大的经济损失。肌肉是畜禽胴体中最主要的组成部分,更是人类肉制品的主要来源。随着生活水平的提高和消费观念的改变,我国人民对肉制品的消费数量逐年上升。同时,食品安全问题日益引起消费者的极大关注,人们对畜产品品质也提出了更高的要求。热应激导致畜禽肉品质下降已成为现代养殖业面临的一个重要问题。高温应激严重危害肉鸡的生产性能,导致肉鸡采食量、日增重下降,死亡率增高[3-4]。据报道,急性热应激或运输应激能够加快肉鸡宰后糖代谢速率,导致低pH、高亮度的PSE肉的发生[5-7],从而影响肉品质。也有关于持续高温应激影响畜禽肌肉品质方面的报道。肉鸡在持续热应激条件下,宰后肌肉pH和红度值降低,剪切力和亮度值提高[8]。持续高温能使生长猪肌纤维类型由慢肌纤维(Ⅰ型)向快肌纤维(Ⅱ型)转化,增加肌肉滴水损失,降低宰后24 h的 pH[9]。许多研究都集中在急性或短期应激对肉鸡[4,10]、羊[11]或者猪[12]的生产性能、血液理化指标及肉品质的影响方面,而对长期慢性高温应激影响猪只尤其是肥育后期猪只胴体性状、背最长肌质构分析、营养物质含量及肌纤维特性等相关指标的研究极少。不容忽视的是,在生产实际中,慢性高温应激在夏季普遍存在,研究持续慢性高温应激对猪肉品质的影响具有一定的现实意义。本研究以生产上最常见的“杜长大”三元杂交阉公猪为试验动物,模拟夏季高温环境,研究持续30℃的高温应激对肥育猪生产性能、胴体性状、背最长肌质构分析、营养物质含量及肌纤维特性的影响,为今后通过营养和环境途径调控肥育猪肉品质提供理论依据。
1 材料与方法
1.1 试验设施
本试验在中国农业科学院北京畜牧兽医研究所昌平试验基地的国家重点实验室环境控制舱内进行。24头猪分别饲养于6间人工气候舱内(舱大小为 4.00 m×3.05 m×2.00 m),每舱 4 头,每头单独放入长宽高为120 cm×80 cm×80 cm的猪笼内,猪笼前部装有鸭嘴式饮水器和料槽,所有肥育猪自由饮水。舱内湿度控制在(55±5)%,照明时间为08:00—22:00,黑暗时间为22:00—08:00。
1.2 试验动物与试验设计
选用的 24头试猪为(79.0±1.5)kg 的“杜长大”三元阉公猪,来自8窝母猪,每窝3头,分别随机分配到3组,每组8个重复,每个重复1头。3组分别为对照(control,CR)组、高温应激(heat stress,HS)组和采食量配对(pair-fed,PF)组,其中对照组昼夜维持恒温22℃,自由采食;高温应激组在试验第1天舱内温度由22℃逐渐升至28℃,每小时升高1℃,第2天由28℃升至29℃,第3天升至30℃,随后维持舱内恒定温度为30℃,直至试验结束,自由采食;采食量配对组昼夜维持恒温22℃,按前1天高温应激组8头猪的平均日采食量进行饲喂。适应期为7 d,适应期内自由采食饮水,舱内温度恒定维持在22℃。试验期为21 d。
1.3 饲养管理
试验猪饲喂典型的玉米-豆粕型饲粮,其组成及营养水平见表1。试验猪每天08:00、12:00和18:00各饲喂1次,饲养管理和免疫按《肥育猪饲养管理手册》进行。

表1 试验饲粮组成及营养水平(饲喂基础)Table 1 Composition and nutrient levels of the experimental diet(as-fed basis) %
1.4 样品采集及制备
试验期结束后,对肥育猪电击晕后倒挂屠宰劈半。快速分离左侧背最长肌,1块在4℃放置1和24 h后对背最长肌横断面大理石花纹评分;3块在4℃放置48 h后分别测定剪切力,进行质构分析;2块约20 g的肉块-20℃保存用于测定肌肉营养物质含量;另取 2块大小为 0.2 cm×0.2 cm×0.2 cm的背最长肌样品置于10%中性福尔马林溶液中固定,待测肌纤维直径和密度。同时将右侧胴体第10肋骨处背最长肌取样后液氮速冻,随后-80℃保存,用于肌纤维类型测定。
1.5 测定指标及方法
1.5.1 生产性能
试验开始前记录肥育猪初始体重,试验期内每天以重复为单位记录猪采食量,计算平均日采食量(ADFI),平均日增重(ADG)和料重比(F/G)。
1.5.2 胴体性状
胴体性状测定参考GB 8467—1987《瘦肉型种猪性能测定技术规程》。胴体重:在猪放血后,去掉毛、头、蹄、尾和内脏(保留板油和肾脏),用磅秤称取胴体的重量。胴体长:将右侧胴体倒挂,用皮尺测量从趾骨联合前缘中点至第一颈椎前缘中点的直线长度。平均背膘厚:将右侧胴体倒挂,用游标卡尺测量胴体背中线肩部最厚处、最后肋、腰荐结合处3点的脂肪厚度,取平均值。皮厚:将右侧胴体倒挂,用游标卡尺测量胴体背中线第6~7肋处皮肤的厚度。
1.5.3 大理石花纹评分
将第12肋骨到最末肋骨处背最长肌取下,4℃放置1、24 h后根据美国猪肉生产商委员会(National Pork Producers Council,NPPC)大理石花纹等级图谱进行横断面大理石花纹评分。
1.5.4 剪切力及质构分析(texture profile analysis,TPA)
用Food Technology Corporation公司的TMSPro质构仪进行剪切力测定和TPA;根据NY/T 1180—2006《肉嫩度的测定 剪切力测定法》测定肉嫩度。取左侧胴体第6~9肋骨处背最长肌,除去肌膜外附着的脂肪,放入封口袋中,4℃放置48 h后,于恒温水浴锅中持续加热至肌肉中心温度达70℃时为止,取出肌肉,室温冷却。用直径为1.27 cm的圆柱状取样器沿肌纤维平行的方向钻切肉样,孔样长度不少于2.5 cm,每个样品4个重复,测定剪切力。将煮后冷却肉样根据肌纤维走向修成2 cm×2 cm×2 cm的小块,每个样品4个重复,进行TPA。
1.5.5 背最长肌营养物质含量
取第9~10肋骨处背最长肌,-20℃保存。根据GB/T 9695.15—2008《肉与肉制品水分含量测定》,以二甲苯蒸馏法检测背最长肌样品中的水分含量;采用凯氏定氮法测定肉样中的粗蛋白质含量;采用 GB/T 5009.6—2003《食品中脂肪的测定》的方法,酸水解后乙醚浸提测定背最长肌样品中的肌内脂肪(IMF)含量。以上3个指标在同一实验室由同一操作者在短暂的时间间隔内、用同一设备对同一试样进行2次独立测试,其测定结果的绝对差值不得超过1%。
1.5.6 背最长肌肌纤维类型组成
1.5.6.1 总 RNA 的提取及反转录
按照超纯RNA提取试剂盒(康为世纪生物科技有限公司,Cat#CW0581)的操作说明书提取肌肉样品总RNA。RNA的纯度和浓度用Nanodrop 2000超微量分光光度计测定(Thermo公司),经1%琼脂糖电泳检测鉴定其完整性。用脱氧核糖核酸酶(DNase)Ⅰ试剂盒(康为世纪生物科技有限公司,Cat#CW2090)对 RNA中残留的基因组DNA进行消化处理,按照HiFi-MMLV cDNA第一链合成试剂盒(康为世纪生物科技有限公司,Cat#CW0744)进行反转录。
1.5.6.2 引物的设计与合成
Ⅰ型肌纤维(MyHCⅠ)、MyHCⅡa、MyHCⅡx和MyHCⅡb基因的荧光定量聚合酶链反应(PCR)引物序列见表2。
1.5.6.3 建立筛选引物标准曲线
以cDNA为模板进行5倍梯度稀释,稀释后样品各取2μL作模板,分别用目的基因引物和内参基因引物进行扩增,同时在60~95℃进行融解曲线分析,并绘制目的基因和内参基因的标准曲线。
1.5.6.4 样品实时荧光定量聚合酶链反应(realtime PCR)分析
本科院校图书馆阅读推广工作尚未形成长效机制,多以读书月形式展开,属于跨部门协同合作项目,一般不单独设岗,因此所调查的招聘信息中阅读推广岗所占比重较小。对行政管理岗位而言,主要要求文字书写能力、管理能力以及策划能力。行政管理类岗位对外招聘信息发布偏少。
将各样品cDNA 10倍稀释后取2μL作模板,分别用目的基因引物和内参基因引物进行扩增。同时在60~95℃进行溶解曲线分析。
1.5.6.5 real-time PCR 扩增效率及特异性检测
利用ABI 7500型荧光定量PCR仪比较3组肥育猪背最长肌MyHC基因表达差异。每次反应设置标准品未知样品(重复3次)和阴性对照。熔解曲线鉴定引物的特异性良好,通过标准曲线扩增效率计算公式,检测到各基因扩增效率在90%~110%。
1.5.6.6 统计分析
MyHCⅠ、MyHCⅡa、MyHCⅡx和 MyHCⅡb基因相对表达量用2-△△CT法计算后进行分析,以β-肌动蛋白(β-actin)作为内参基因,选择对照组猪背最长肌中的基因表达作为参照。

表2 实时荧光定量聚合酶链反应引物序列Table 2 Primer sequences of real-time PCR
1.5.7 背最长肌组织切片(肌纤维直径、密度)
1.5.7.1 制片
取出用10%中性福尔马林溶液固定的背最长肌样品,经过修块、浸洗后依次用50%乙醇、70%乙醇、80%乙醇、90%乙醇、95%乙醇、100%乙醇进行脱水处理;再依次浸入二甲苯与乙醇1∶1的混合溶液、二甲苯溶液中进行透明处理;透明后的背最长肌组织块在恒温培养箱中依次经过熔点分别为54~56℃,56~58℃、58~60℃的石蜡,再用60℃的石蜡进行包埋处理;经过冷却修整后用切片机进行连续切片,厚度为5μm,再经过展片、粘片后制成备用的石蜡切片。
1.5.7.2 苏木精-伊红(HE)染色
1.5.7.3 观察
用专业图像分析软件(Image-Pro Plus 6.0)分析切片,每个样品制作2张切片,每张切片至少选择3个视野,从而计算肌纤维直径和密度。
1.6 数据统计分析
试验数据以平均值±标准差表示,数据采用SAS 9.2软件进行ANOVA单因素方差分析(oneway ANOVA),且用Duncan氏法进行多重比较检验3组间的差异显著性。以P<0.05为差异显著,P<0.01 为差异极显著。
2 结果
2.1 持续高温应激对肥育猪生产性能的影响
由表3可知,高温应激组和采食量配对组肥育猪的平均日采食量极显著低于对照组(P<0.01)。高温应激组肥育猪的平均日增重显著低于对照组(P<0.05)。3组之间的料重比差异不显著(P>0.05)。
2.2 持续高温应激对肥育猪胴体性状的影响
试验结束后屠宰所有试验猪,测定肥育猪胴体性状。由表4可知,3组之间胴体重、胴体长、平均背膘厚和皮厚均差异不显著(P>0.05)。3组的大理石花纹评分在宰后1 h差异不显著(P>0.05),高温应激组肥育猪的背最长肌大理石花纹评分在宰后24 h有低于对照组的趋势(P<0.10)。
2.3 持续高温应激对肥育猪背最长肌剪切力及TPA的影响
由表5可知,高温应激组肥育猪背最长肌剪切力极显著高于对照组和采食量配对组(P<0.01)。在TPA中,高温应激组肥育猪背最长肌胶黏性和硬度有低于采食量配对组的趋势(P<0.10)。3组之间的背最长肌的弹性、咀嚼性及内聚性无显著差异(P>0.05)。

表3 持续高温应激对肥育猪生产性能的影响Table 3 Effects of constant heat stress on performance in finishing pigs(n=8)
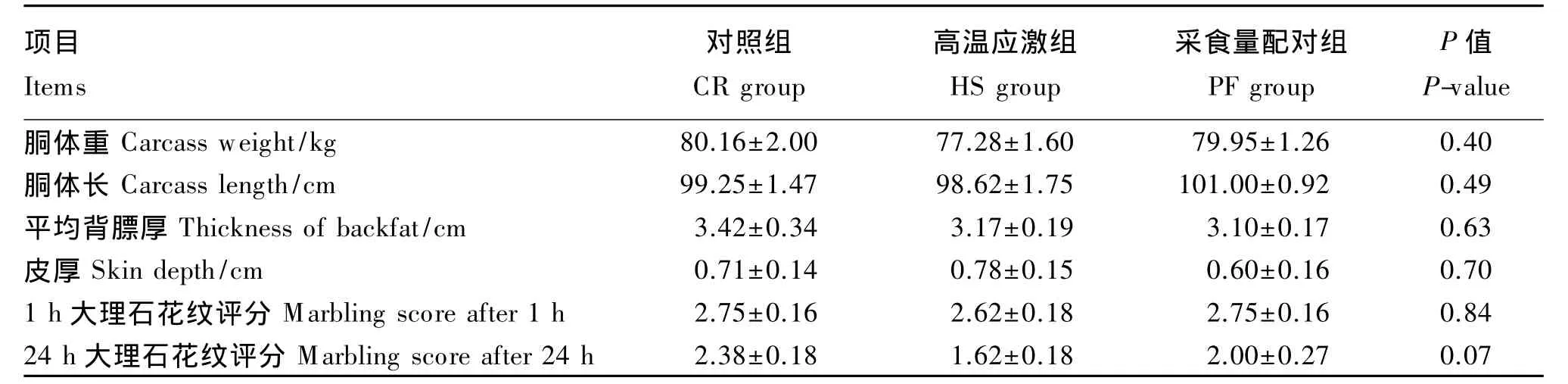
表4 持续高温应激对肥育猪胴体性状的影响Table 4 Effects of constant heat stress on carcass traits in finishing pigs(n=8)
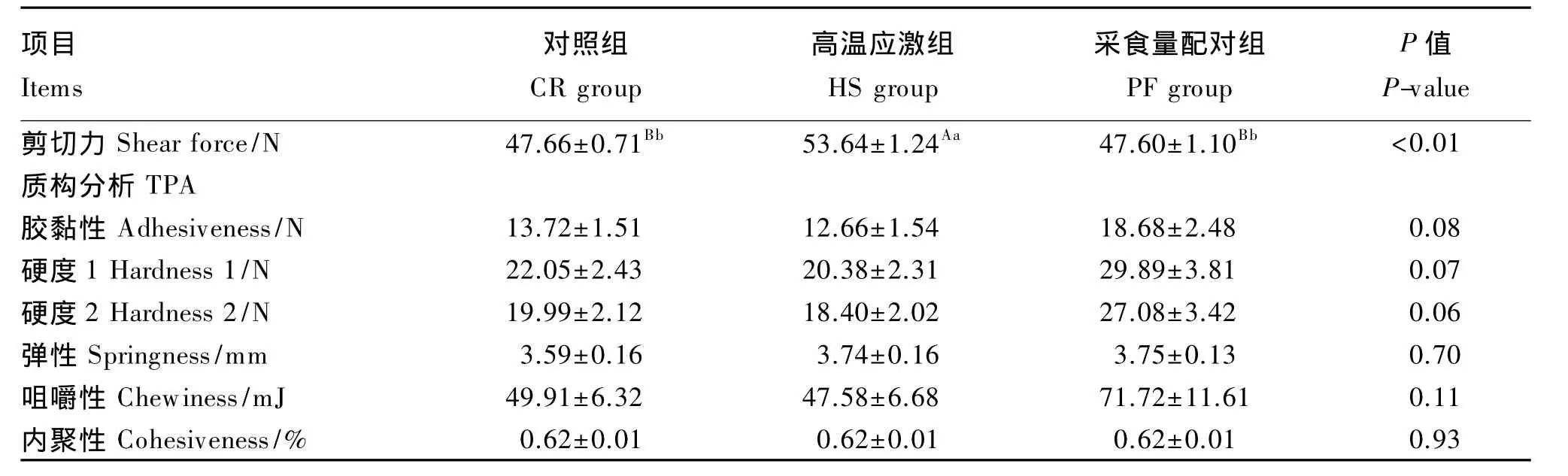
表5 持续高温对肥育猪背最长肌剪切力和TPA的影响Table 5 Effects of constant heat stress on shear force and texture profile analysis of longissimus dorsi in finishing pigs(n=8)
2.4 持续高温应激对猪背最长营养物质含量的影响
由表6可知,通过测定背最长肌营养物质含量发现,高温应激组和采食量配对组肥育猪背最长肌粗蛋白质含量显著高于对照组(P<0.05),且背最长肌肌内脂肪含量极显著低于对照组(P<0.01)。3组之间背最长肌干物质含量无显著差异(P>0.05)。

表6 持续高温应激对肥育猪背最长肌干物质、粗蛋白质和肌内脂肪含量的影响Table 6 Effects of constant heat stress on DM,CP and IMF contents of longissimus dorsi in finishing pigs(n=8)%
2.5 持续高温应激对猪背最长肌肌纤维类型、直径和密度的影响
由表7可知,通过real-time PCR测定背最长肌的肌纤维类型组成,结果显示高温应激组的MyHCⅠ表达量有低于采食量配对组的趋势(P<0.10),采食量配对组的MyHCⅡx表达量显著低于对照组(P<0.05)。由表8可知,3组之间肥育猪肌纤维直径和密度均无显著差异(P>0.05)。
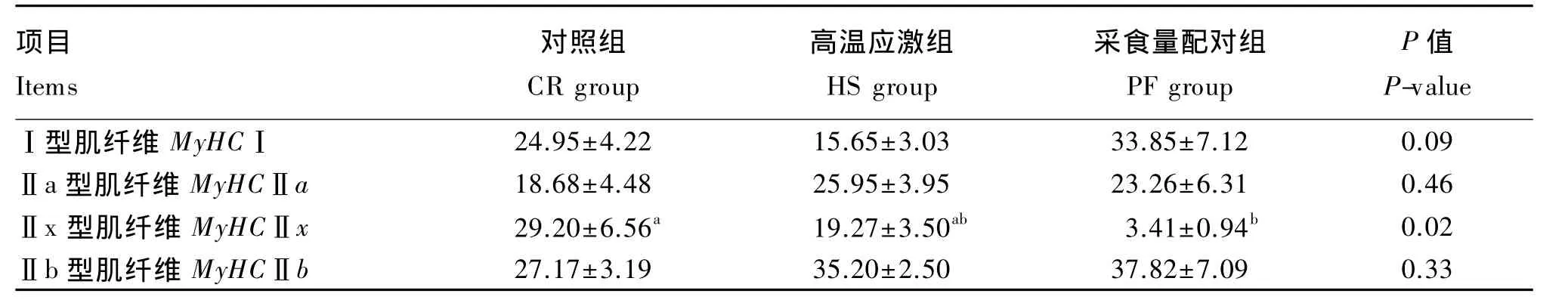
表7 持续高温应激对肥育猪背最长肌肌纤维类型组成的影响Table 7 Effects of constant heat stress on myofiber type of longissimus dorsi in finishing pigs(n=4) %

表8 持续高温应激对肥育猪背最长肌肌纤维直径和密度的影响Table 8 Effects of constant heat stress on muscle fiber density and diameter of longissimus dorsi in finishing pigs
3 讨论
3.1 持续高温应激对肥育猪生产性能的影响
本研究中,持续高温应激会显著降低肥育猪平均日采食量和平均日增重,该结果在前人的研究中已被证实[13]。当肥育猪长期饲养于高温环境中时,机体代谢发生变化,采食量自发下降、体增热减少,从而将体温维持在正常的水平。由于采食量配对组肥育猪的饲喂量是按前1天高温应激组肥育猪的平均日采食量供给的,所以采食量配对组肥育猪的平均日采食量也显著降低。在高温应激组和采食量配对组肥育猪的采食量一致的情况下,高温应激组肥育猪的平均日增重比采食量配对组低100 g左右,说明持续高温应激导致肥育猪平均日增重显著降低,而且这种变化不依赖于采食量的影响。因此,持续高温应激显著降低肥育猪平均日采食量和平均日增重,对畜牧生产有不利影响。
3.2 持续高温应激对肥育猪胴体性状的影响
本研究发现,在3组之间,肥育猪的胴体重、胴体长、平均背膘厚和皮厚均无显著差异,其可能原因是肥育猪为大动物,个体差异大,试验样本数量有限,同时热应激时间和强度尚未达到出现显著性差异的程度。肌内脂肪主要分布在肌束和肌纤维之间,其积累量和分布的变化使肌肉呈现出不同程度的大理石样花纹[14]。将肥育猪宰后1和24 h的背最长肌横断面与NPPC大理石花纹图谱对比,可以得到背最长肌大理石花纹评分。结果显示高温应激组肥育猪宰后24 h的背最长肌大理石花纹评分有低于对照组的趋势,说明从直观上看,高温应激组肥育猪背最长肌肌内脂肪含量较少,该结果与高温应激组背最长肌肌内脂肪含量的显著降低相一致。
3.3 持续高温应激对肥育猪背最长肌剪切力及TPA的影响
本试验中,持续高温应激组肥育猪背最长肌剪切力显著高于对照组和采食量配对组,说明持续高温应激导致肥育猪背最长肌剪切力提高,其肌肉嫩度较小、肉质较老。该结果与多位研究者的结果相同。卢庆萍等[9]报道,恒定33℃的持续高温应激会增加生长猪肌肉的剪切力。Zhang等[8]研究发现日循环高温(29~35℃)或持续高温应激组(34℃)的肉鸡胸肌剪切力均显著高于对照组,持续高温应激组(34℃)的肉鸡腿肌剪切力也显著高于对照组。方瑞[15]和冯京海[16]发现在日循环高温环境下,肉鸡胸肌剪切力显著提高。
质构仪(texture analyser)又叫物性测试仪,在质构仪的多种测试模式中,TPA是应用最广泛的一种测试模式。TPA又被称为2次咀嚼测试,主要是通过模拟人口腔的咀嚼运动,对样品进行2次压缩,通过界面输出质构测试曲线,从中分析质构特性参数:硬度、内聚性、弹性、胶黏性、咀嚼性等。本研究的TPA结果显示,高温应激组肥育猪背最长肌的胶黏性和硬度有低于采食量配对组的趋势,说明相对于采食量配对组,咀嚼前破碎高温应激组的背最长肌样品需要的能量较低,且牙齿挤压高温应激组样品需要的力量较小。在高温应激组和采食量配对组肥育猪采食量一致的情况下,单纯高温应激导致了肥育猪肉品质的改变,而不依赖于采食量的影响。
3.4 持续高温应激对肥育猪背最长肌营养物质含量的影响
营养物质含量对肌肉品质特性有重要影响,也可反映畜禽对饲粮营养物质的利用和沉积状况[17]。在本试验中,高温应激对肥育猪背最长肌干物质含量无影响,但会显著增加背最长肌粗蛋白质含量,极显著降低背最长肌肌内脂肪含量。与本试验结果不同的是,有学者研究发现热应激会使肉鸡和生长猪肌肉水分含量增加,干物质含量减少。Kerr等[18]发现,与生长于23℃适温环境的生长猪相比,饲养于33℃下的生长猪肌肉干物质含量较少。Zhang等[8]研究发现热应激会增加肉鸡胸肌水分含量。本研究中背最长肌干物质含量的结果与前人不同,可能是由于本试验应激强度较低、持续时间较长、动物机体代谢趋于适应且试验对象为肥育猪而造成的。
关于热应激对畜禽肌肉粗蛋白质和肌内脂肪含量的影响,学者们的研究结果不完全一致。刘圈炜等[17]的结论与本试验类似,33℃的持续热应激能提高生长猪肌肉粗蛋白质含量,降低生长猪肌内脂肪含量。同样有2位研究者发现肥育猪在热循环高温环境下肌肉粗蛋白质含量会显著升高[19-20]。与之相反地,Zhang 等[8]研究发现热应激会使肉鸡胸肌粗蛋白质含量显著降低,脂肪沉积显著增加,早年也有另外2位学者报道热应激使鸡的脂肪沉积增加[21-22]。对于一些与本研究结果相反的报道,可以解释为由于猪和鸡的脂肪代谢机理不同,导致了热应激情况下2个物种肌内脂肪沉积效果相反。肥育期猪只的采食量对肥育效果有重要的影响,在持续高温应激情况下,肥育猪采食量减少,所获能量优先用于生命维持和机体骨架蛋白质构成,其次用于脂肪沉积,从而使得肥育猪肌肉中肌内脂肪的沉积相对减少,肌肉中粗蛋白质含量则会相对提高。
另外我们发现,高温应激组肥育猪背最长肌的营养物质变化规律与采食量配对组相一致,说明肥育后期的人为限制饲喂也造成了背最长肌中肌内脂肪含量的减少,与前人[23-24]报道的限饲使猪背最长肌肌内脂肪含量显著下降的结果类似。本结果提示,高温应激造成的肥育猪背最长肌肌内脂肪含量的减少、粗蛋白质含量的增加,可能是由于持续高温应激导致肥育猪采食量的减少引起的。
3.5 持续高温应激对肥育猪背最长肌肌纤维类型、直径和密度的影响
肌纤维按照氧化代谢能力由高到低可分为Ⅰ型、Ⅱa型、Ⅱx型、Ⅱb型,其中Ⅰ型代表慢速氧化型,Ⅱa型代表快速氧化型,Ⅱx型代表混合型,Ⅱb型代表快速酵解型。慢速氧化型肌纤维又被称为红肌或慢肌纤维。高温应激组的MyHCⅠ表达量低于采食量配对组,说明在2组肥育猪采食量相同的情况下,高温应激导致MyHCⅠ表达量减少,肌纤维类型发生变化。本结果提示,高温应激导致了红肌纤维相对含量的减少,且这种变化不依赖于采食量的影响。卢庆萍等[9]报道,持续热应激使生长猪MyHCⅡ含量增加、MyHCⅠ含量减少,与本试验的结果类似。另外也有关于热应激导致骨骼肌细胞肌纤维类型转化的研究,且2者的结果正相反:Yamaguchi等[25]研究发现大鼠C2C12细胞在39℃下培养会使其由快肌纤维(MyHCⅡx)向慢肌纤维(MyHCⅠ)转变;而Frier等[26]的结果则是热应激会抑制大鼠MyHCⅠ含量的增加。但由于细胞与活体肥育猪对外界高温应激的调控机制有很大不同,局部细胞试验仅能作为参考。
采食量配对组肥育猪的混合型肌纤维(MyHCⅡx)表达量显著低于对照组,说明限饲对肌纤维类型转化有一定影响。李玥[27]报道称14日龄早期限饲的营养缺乏会抑制肉仔鸡肌纤维类型的正常转化,慢肌纤维比例较多。虽然由于试验对象及限饲时期的不同,李玥的结果与本试验并不一致,但却提示限饲可能会导致畜禽肌纤维类型的转化。
本研究中,高温应激不会造成肥育猪背最长肌肌纤维直径和密度的显著变化。经国内外研究发现,影响猪肌纤维直径和密度的最主要因素为品种,国外瘦肉型猪如杜长大、大白、长白等肌纤维直径一般为 54~73μm[28],而我们地方品种猪如莱芜猪的肌纤维直径为44~57μm[29]。另外,肥育猪性别、年龄和饲粮营养水平的不同也会对肌纤维直径和密度产生一定影响。本研究使用的肥育猪品种、性别和年龄相同,说明在肥育猪后期供给采食量配对组较少的饲粮不会对肌纤维直径和密度产生影响,肥育猪在30℃的持续高温应激环境下饲喂21 d也不会造成背最长肌肌纤维直径和密度的显著变化。
4 结论
持续高温应激会使肥育猪平均日采食量和平均日增重显著下降,背最长肌剪切力显著提高,肌肉粗蛋白质含量显著升高,肌内脂肪含量极显著降低,并使肌纤维类型发生转化,从而影响肉品质。
[1] LIN H,BUYSE J,DECUYPERE E.Acute heat stress induces oxidative stress in broiler chickens[J].Comparative Biochemistry and Physiology:Part A,2006,144(1):11-17.
[2] LU Q P,WEN J,ZHANG H F.Effect of chronic heat exposure on fat deposition and meat quality in two genetic types of chicken[J].Poultry Science,2007,86(6):1059-1064.
[3] COOPER M A,WASHBURN K.The relationships of body temperature to weight gain,feed consumption,and feed utilization in broilers under heat stress[J].Poultry Science,1998,77(2):237-242.
[4] QUINTEIRO-FILHO W M,RIBEIRO A,FERRAZDE-PAULA V,et al.Heat stress impairs performance parameters,induces intestinal injury,and decreases macrophage activity in broiler chickens[J].Poultry Science,2010,89(9):1905-1914.
[5] NORTHCUTT J K,FOEGEDING E A,EDENS F W.Water-holding properties of thermally preconditioned chicken breast and leg meat[J].Poultry Science,1994,73(2):308-316.
[6] MCKEE S R,SAMS A R.The effect of seasonal heat stress on rigor development and the incidence of pale,exudative turkey meat[J].Poultry Science,1997,76(11):1616-1620.
[7] SANDERCOCK D A,HUNTER R R,NUTE G R,et al.Acute heat stress-induced alterations in blood acidbase status and skeletal muscle membrane integrity in broiler chickens at two ages:implications for meat quality[J].Poultry Science,2001,80(4):418-425.
[8] ZHANG Z Y,JIA G Q,ZUO JJ,et al.Effects of constant and cyclic heat stress on muscle metabolism and meat quality of broiler breast fillet and thigh meat[J].Poultry Science,2012,91(11):2031-2937.
[9] 卢庆萍,张宏福,刘圈炜,等.持续高温对生长猪肌纤维特性及肉质性状的影响[C]//第六次全国饲料营养学术研讨会论文集.北京:中国畜牧兽医学会,2010:542-543.
[10] PETRACCI M,BETTI M,BIANCHI M,et al.Color variation and characterization of broiler breast meat during processing in Italy[J].Poultry Science,2004,83(12):2086-2092.
[11] CHULAYO A Y,MUCHENJE V.The effects of preslaughter stress and season on the activity of plasma creatine kinase and mutton quality from different sheep breeds slaughtered at a smallholder abattoir[J].Asian-Australasian Journal of Animal Sciences,2013,26(12):1762-1772.
[12] WESCHENFELDER A V,TORREY S,DEVILLERS N,et al.Effects of vehicle design on blood stress indicators and meat quality in pigs of three genotypes for two different travel distances[J].Journal of Animal Science,2010,88(Suppl.2):464-465.
[13] BELLEGO L L,VANMIGEN J,NOBLET J.Effect of high temperature on protein and lipid deposition and energy utilization in growing pigs[J].Animal Science,2002,75:85-96
[14] 张艳芳.CPT1基因对猪脂肪沉积的影响及其调控机制的研究[D].博士学位论文.杭州:浙江大学,2010.
[15] 方瑞.持续外界高温对肉鸡生长、福利状况、肉品质的影响及其调控措施研究[D].硕士学位论文.呼和浩特:内蒙古农业大学,2009.
[16] 冯京海.环境高温对肉鸡线粒体功能及胸肌品质的影响[D].博士学位论文.北京:中国农业科学院,2006.
[17] 刘圈炜,卢庆萍,张宏福,等.持续高温对生长猪血清生化指标及肌肉营养物质含量的影响[J].动物营养学报,2010,22(5):1207-1213.
[18] KERR B J,YEN J T,NIENABER J A,et al.Influences of dietary protein level,amino acid supplementation and environmental temperature on performance,body composition,organ weights and total heat production of growing pigs[J].Journal of Animal Science,2003,81(8):1998-2007.
[19] LOPEZ J,GOODBANDRD R D,ALLEN G L,et al.The effects of diets formulated on an ideal protein basis on growth performance,carcass characteristics,and thermal balance of finishing gilts housed in a hot,diurnal environment[J].Journal of Animal Science,1994,72(2):367-379.
[20] LEFUCHED L,LE DIXIDICH J,MOUROT J,et al.Influence of environmental temperature on growth,muscle and adipose tissue metabolism and meat quality in swine[J].Journal of Animal Science,1991,69(7):2844-2854.
[21] AIN BAZIZ H,GERAERT P A,GUILLAUMIN S,et al.Chronic heat exposure enhances fat deposition and modifies muscle and fat partition in broiler carcasses[J].Poultry Science,1996,75(4):505-513.
[22] GERAERT P A,PADILHA J C F,GUILLAUMIN S.Metabolic and endocrine changes induced by chronic heat exposure in broiler chickens:growth performance,body composition and energy retention[J].British Journal of Nutrition,1996,75(2):195-204.
[23] LEYMASTER K A,MERSMANN H J.Effect of limited feed intake on growth of subcutaneous adipose tissue layers and on carcass composition in swine[J].Journal of Animal Science,1991,69(7):2837-2843.
[24] WIECEK J,REKIEL A,BATORSKA J,et al.Effect of restricted feeding and realimentation periods on pork quality and fatty acid profile of M.longissimus thoracis[J].Meat Science,2011,87(3):244-249.
[25] YAMAGUCHIT,SUZUKI T,ARAIH,et al.Continuous mild heat stress induces differentiation of mammalian myoblasts,shifting fiber type from fast to slow[J].American Journal of Physiology:Cell Physiology,2010,298(1):C140-C148.
[26] FRIER B C,LOCKE M.Heat stress inhibits skeletal muscle hypertrophy[J].Cell Stress & Chaperones,2007,12(2):132-141.
[27] 李玥.限饲对肉鸡腓肠肌肌纤维与为卫星细胞形态与功能的影响及机理研究[D].博士学位论文.南京:南京农业大学,2008.
[28] STAUN H.The mutritional and genetic influence on number and size of muscle fibers and their response to carcass quality in pigs[J].World Review of Animal Production,1972,8(3):18-26.
[29] ZANG Y Q,WEI S D,WANG H,et al.Studies on meat quality properties of Shandong native pig breed[C]//Proceedings of international conference on pig production.Beijing:InterAcad publishers,1998:663-666.
