一株稻纵卷叶螟颗粒体病毒的系统发育分析和流行病学调查
2014-12-16贾茜雯孙士锋陈其津
张 珊,贾茜雯,孙士锋,庞 义,陈其津,杨 凯*
(1.中山大学有害生物控制与资源利用国家重点实验室,广州 510275;2.华南师范大学附属中学,广州 510630;3.广东海纳农业有限公司,广东惠州 516005)
稻纵卷叶螟Cnaphalocrocis medinalis Guenée 属于鳞翅目螟蛾科昆虫,俗名纵卷螟、苞叶虫、稻纵卷叶虫等,是一种典型的迁飞性害虫,寄主有水稻、大麦、小麦、玉米等农作物,以及茅草、狗尾草、芦苇等禾本科杂草(洪晓月等,2007)。稻纵卷叶螟在国内的分布地区为东自台湾,西迄西藏,南起海南岛,北到黑龙江,遍及28个省、市、自治区,南方以及长江中下游种植水稻的区域受到这种害虫的危害最为严重,目前为水稻的三大害虫之一(张孝羲等,1980)。
稻纵卷叶螟颗粒体病毒(Cnaphalocrocis medinalis granulovirus,CnmeGV)由中山大学庞义教授等(1981)首次报道。该病毒于1979年秋从广东省江门市恩平县晚造水稻田里的罹病稻纵卷叶螟幼虫中分离,田间宿主的死亡率高达30%-40%,虫口密度大的田块尤多(庞义等,1981)。因此,CnmeGV 是一种很有潜力的生物防治剂,可以用来控制稻纵卷叶螟的发生,减轻对水稻的危害。
杆状病毒是专一性感染昆虫的病毒,基因组为大型闭合环状双链DNA,衣壳呈杆状,具有嚢膜包被,病毒粒子包埋于包涵体中(Rohrmann,2013)。颗粒体病毒(granulovirus,GV)是杆状病毒的一个属,其包涵体为卵圆形,直径在200-400 nm 之间,一般仅包埋单个杆状的病毒粒子(Rohrmann,2013)。目前已分离的GV 共有148 株(Rohrmann,2013),而收录在GenBank 中的全基因组序列已测定的GV 仅有17 株(Rohrmann et al.,2013;Zhang et al.,2014;Zhu et al.,2014)。
CnmeGV 是根据首次分离到该病毒的宿主名称来命名的,这是普遍采取的一种命名病毒的方法。但是有些杆状病毒可以感染不同的宿主,因此在GenBank 中有些不同名称的病毒事实上是同一种病毒(Rohrmann,2013)。所以新分离病毒在开展全基因组序列测定之前,最好先进行分子水平上的鉴定。lef-8 和lef-9 基因编码杆状病毒RNA 聚合酶的两个亚基(Guarino et al.,1998;Herniou et al.,2003)。gran 基因编码GV 包涵体中主要的基质蛋白(de Moraes and Maruniak,1997;Zanotto et al.,1993)。以前的研究发现,由gran、lef-8和lef-9 基因部分融合序列推导的进化树与用杆状病毒核心基因融合序列推导的系统发育树在拓扑结构上完全一致(Lange et al.,2004;Jehle et al.,2006)。
本研究利用稻纵卷叶螟幼虫在实验室中成功增殖了CnmeGV,并扩增出CnmeGV 的gran、lef-8 和lef-9 片段,利用MEGA4 软件进行了系统发育分析。结果表明,GenBank 中没有CnmeGV 序列报道。此外,本研究还对几个地区的稻田的CnmeGV 流行病学进行了初步的调查。
1 材料与方法
1.1 材料
1.1.1 病毒
CnmeGV:于1979年采自江门市恩平县晚造水稻罹病稻纵卷叶螟幼虫,在栽有水稻的罩笼内饲养幼虫并扩增病毒后,4℃保存于本实验室(蒲蛰龙等,1986)。
1.1.2 昆虫
稻纵卷叶螟:2012年5月中旬-7月下旬,采自广州市岑村华南农业大学水稻试验田内的野生种群,室内继续培养,温度为(26±0.5)℃,相对湿度为90%-95%,光周期为光∶暗=16 h∶8 h。
1.2 方法
1.2.1 CnmeGV 的增殖
选取3-4 龄稻纵卷叶螟幼虫放入饲养盒中饥饿4-5 h,喂以混合CnmeGV 的水稻汁液。覆盖新鲜水稻叶片,滴加1 mL 灭菌ddH2O,有孔保鲜膜封闭,湿润棉球保湿。拍照记录感染病毒后稻纵卷叶螟的病症(如图1)。
1.2.2 病毒包涵体的纯化
向染病死亡的虫尸中加入适量的PBS(pH=7.4),研磨并用四层纱布过滤。经差速离心和蔗糖密度梯度离心,获得纯化的CnmeGV 包涵体(O'Reilly et al.,1992)。
1.2.3 PCR 扩增gran、lef-8、lef-9 基因片段及序列测定
参照(O'Reilly et al.,1992)从包涵体中提取病毒DNA。PCR 扩增gran、lef-8 及lef-9 基因片段,引物序列见(Jehle et al.,2006)。由上海Invitrogen 生物工程公司完成扩增片段的序列测定。Seqman 5.01 软件对双向测序结果进行拼接、校对、去除通用引物。
1.2.4 系统进化树的构建
用于构建系统发生树的杆状病毒包括CnmeGV、14 株已测序GV,以及作为外类群的苜蓿丫纹夜蛾核多角体病毒(Autographa californica multiple nucleopolyhedrovirus,AcMNPV)。除CnmeGV外,其余病毒的gran、lef-8 及lef-9 基因序列均下载自GenBank(Ayres et al.,1994;Escasa et al.,2006;Ferrelli et al.,2012;Harrison et al.,2008;Liang et al.,2011;Wormleaton et al.,2003;Zhang et al.,2012)。将下载序列导入MEGA4 软件,对序列进行多重比对和融合,选择UPGMA 法则,构建系统发生树,并对各分支节点进行Bootstrap 检验(检测1000 次,每次检测随机加样重复100次)。
1.2.5 稻纵卷叶螟样品的采集和体内CnmeGV 的分子生物学检测
采集地点为华南农业大学试验田(广州市岑村)、惠州市新岗村、惠州市万芦村、江门市恩平县、湖北省潜江市。采集幼虫虫苞,将虫苞分为两部分:一部分分装于保鲜袋中,在实验室内饲养4-5 d 后,观察是否有感染死亡的稻纵卷叶螟幼虫,并统计发病率;一部分在田间剥开,将单头幼虫放入95%乙醇中,每个地点取样20 头。在实验室提取稻纵卷叶螟DNA(Chai et al.,2011)。PCR 检测虫体内是否存在CnmeGV 的lef-8 基因片段。

图1 实验室增殖CnmeGVFig.1 Propagation of CnmeGV in laboratory
2 结果与分析
2.1 稻纵卷叶螟幼虫感染CnmeGV 的病症
感染CnmeGV 后,稻纵卷叶螟幼虫从正常的半透明状态逐渐变成灰白色,进而变成白色不透明体或者略带橙黄色,行动迟缓。罹病前期和中期仍能卷叶取食,但食欲减退。病虫多数不能化蛹,即使化蛹也会在蛹期死亡。幼虫一般在感染病毒后5-10 d 死亡,刚死亡时虫体为乳白色,体内液化(如图2-C)。用枪头反复吹打虫尸,可释放出含病毒的体液,而虫皮保持完整(如图2-E)。如虫尸在室温放置1-2 d,虫体将完全液化,且呈现紫酱色(如图2-D)。
2.2 基于gran、lef-8 及lef-9 基因融合序列的系统发生树的构建和分析
基于gran、lef-8 及lef-9 基因部分序列,我们构建了UPGMA 融合系统发生树。如图3 所示,系统发生树节点处数值表示该分支的相对置信度,该树的置信度较高,只有两个节点的数值在50%以下。而单独以gran、lef-8 以及lef-9 基因构建的系统发生树分支的置信度普遍低于50%(结果未显示)。

图2 稻纵卷叶螟幼虫感染CnmeGV 后的形态学病征观察Fig.2 The symptoms of C.medinalis larvae infected with CnmeGV
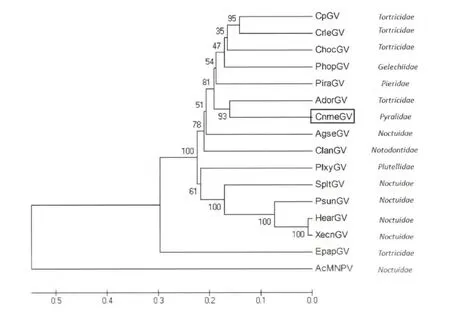
图3 基于gran、lef-8 和lef-9 基因部分融合序列的UPGMA 系统发生树Fig.3 UPGMA distance tree based on concatenated DNA sequences of the partial gran,lef-8 and lef-9 genes
由图3 可知,CnmeGV 和茶小卷叶蛾颗粒体病毒(Adoxophyes orana granulovirus,AdorGV)是一个单系类群的几率为93%(单系类群是指包含一个祖先类群的所有子裔的群组),即CnmeGV 和AdorGV 由同一祖先进化而来的概率为93%。在所有全基因组已测序的GV 中,与CnmeGV 亲缘关系最近的是AdorGV,进化距离为它们各自到最近节点的分支长度的和,约为0.35,且AdorGV 与CnmeGV 的gran、lef-8 和lef-9 融合基因在软件MegAlign 里比对后显示相似度只有59.8%。两种结果都表明与 CnmeGV 亲缘关系最近的为AdorGV,但相似度不高,可以确定CnmeGV 是一个未收录在GenBank 中的独立病毒株,为一个新种。
图3 中还显示宿主为卷叶蛾科幼虫的GV 除了大豆叶小卷蛾颗粒体病毒(Epinotia aporema granulovirus,EpapGV)之外,都集中分布在系统发生树的同一支内,且节点处有81%的置信度,而宿主为螟蛾科幼虫的CnmeGV 也被分在了这一支内,有可能是因为CnmeGV 是在进化的过程中从其他卷叶蛾幼虫体内转移到稻纵卷叶螟体内的。宿主为夜蛾科幼虫的GV 大部分也被分在了同一支内,如棉铃虫颗粒体病毒(Helicoverpa armigera granulovirus,HearGV)、八字地老虎颗粒体病毒(Xestia c-nigrum granulovirus,XecnGV)、美洲粘虫颗粒体病毒(Pseudelatia unipuncta granulovirus,PsunGV)和斜纹夜蛾颗粒体病毒(Spodoptera litura granulovirus,SpltGV)这一类群为单系类群的概率为100%,即它们是由共同祖先进化而来的概率为100%,其中HearGV 与XecnGV 亲缘关系极近,进化距离只有0.02 左右。系统发生树的分支情况显示GV 的分支与宿主是存在一定的联系的,病毒与宿主之间存在一定的协同进化关系。
2.3 CnmeGV 流行病学初步调查结果
分别从广州市岑村、惠州市万芦村、惠州市新岗村、湖北省潜江市和江门市恩平县采集稻纵卷叶螟幼虫,观察是否有CnmeGV 感染。在实验室饲养5-6 d 后,从江门市恩平县采集的幼虫出现了病毒感染死亡的症状,死亡率达36.7%(如表2)。抽提虫尸内的病毒基因组,利用上述lef-8基因引物,通过PCR 扩增可得到一DNA 片段(如图4)。序列比对表明,扩增片段的核苷酸序列与之前实验室保存的CnmeGV 扩增出的lef-8 基因序列完全一致,证明该病毒为CnmeGV。其他四个地区所采集的稻纵卷叶螟幼虫没有出现病毒感染的症状,PCR 未检测到虫体内存在CnmeGV 的lef-8片段(如图5),表明所检测的四个地区的稻纵卷叶螟种群中可能不存在CnmeGV。
3 结论与讨论
本实验室将从田间采集的稻纵卷叶螟成虫放置在栽有水稻的罩笼内,产卵,待孵化幼虫进入二龄后,喷以CnmeGV 致死虫尸的悬浮液,使幼虫感染并扩增CnmeGV(蒲蛰龙等,1986)。本研究建立了一种在实验室方便快捷地扩增CnmeGV的新方法。将采集的稻纵卷叶螟幼虫在饲养盒中饥饿处理4-5 h,喂以混合CnmeGV 的水稻汁液,水稻汁液具备清新水稻气味,能够吸引饥饿的幼虫取食。在覆盖新鲜水稻叶片后,加灭菌ddH2O、湿润棉球以及有孔保鲜膜,不仅可以保证水稻叶片的新鲜,还可以维持稻纵卷叶螟生长的湿润环境。
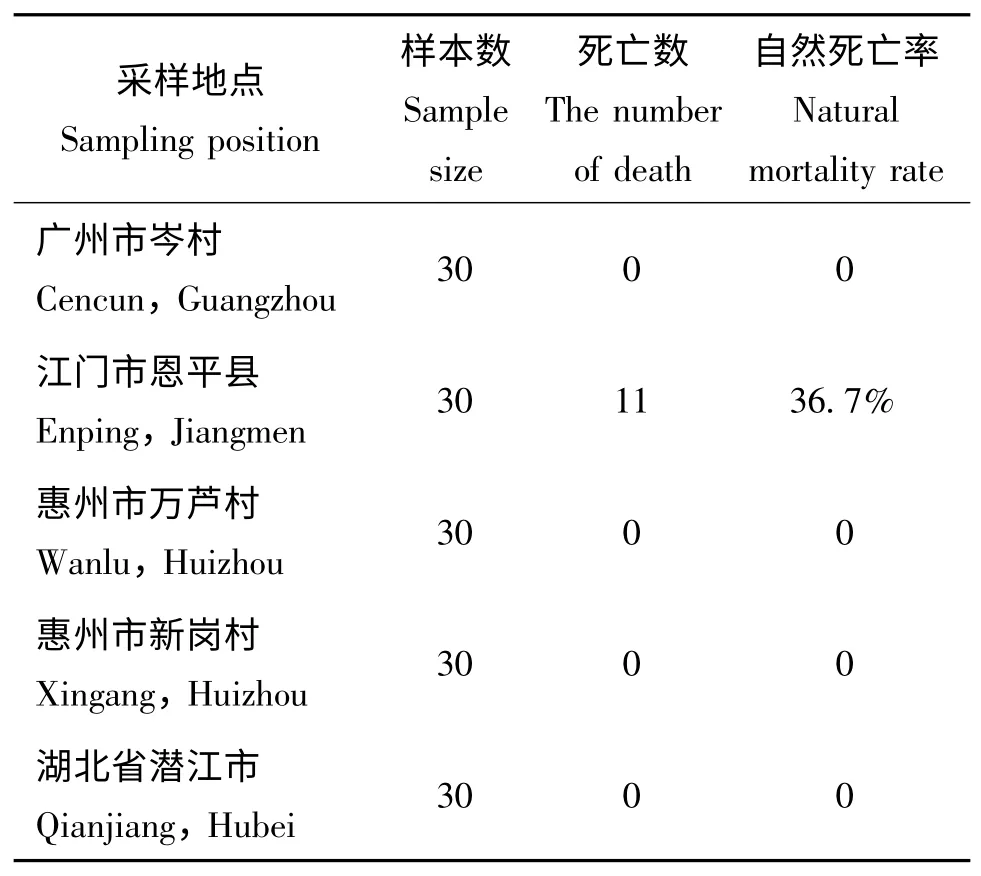
表1 不同地区稻纵卷叶螟自然染病死亡率统计Table 1 The natural mortality rate of wild C.medinalis larvae collected from different places
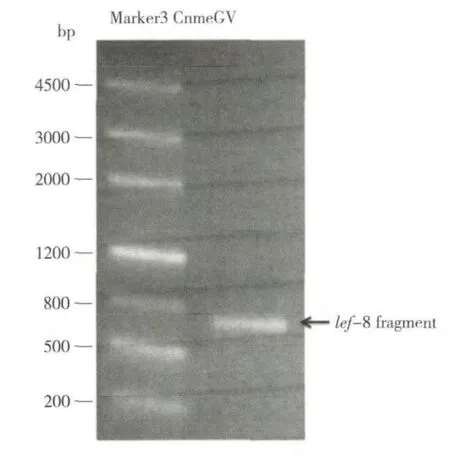
图4 江门市恩平县稻纵卷叶螟幼虫体内CnmeGV 的检测Fig.4 Electrophoretogram of PCR products amplified from the C.medinalis larvae collected from Jiangmen Enping
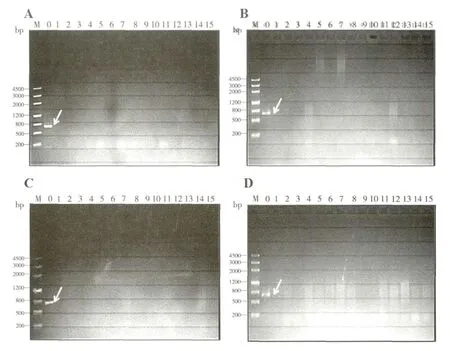
图5 不同地区稻纵卷叶螟幼虫体内CnmeGV 的检测Fig.5 Electrophoretogram of PCR products amplified from the C.medinalis larvae collected from different locations
根据感染昆虫宿主的组织趋向性不同,GV 可以被分为三种类型:Type 1 型GV 代表种为粉纹夜蛾颗粒体病毒(Trichoplusia ni granulovirus,TnGV),感染局限于宿主的中肠和脂肪体,不能侵染表皮和气管皮膜组织等重要的组织,幼虫死亡时虫体仅有一点或者是完全没有液化的现象;Type 2 型GV 代表种为苹果小蠹蛾颗粒体病毒(Cydia pomonella granulovirus,CpGV),这类病毒感染的途径和染病后虫体的病症与典型的鳞翅目核多角体病毒病类似,在侵染中肠后,能够侵染包括表皮、脂肪体和气管皮膜组织等虫体的主要组织,随着感染时间的增长,幼虫逐渐出现轻微的肿胀,并且在表皮下方有不规则的黄白色斑块,死后虫体会液化;Type 3 型GV 只有唯一的一株葡萄叶斑蛾颗粒体病毒(Harrisina brillians granulovirus,HbGV),这种病毒只感染宿主的中肠上皮组织,导致宿主因中肠上皮细胞的大量脱落影响排泄系统而快速死亡(Federici,1997)。本研究对稻纵卷叶螟幼虫感染CnmeGV 的病症进行了细致的观察,发现感染虫体的大部分组织均被CnmeGV 侵染,但没有完全液化,这与Type2 型的GV 所表现出来的特征相近。
昆虫流行病学是研究昆虫种群疾病的成因和形式的一门科学,主要内容包括:与疾病流行有关的寄主昆虫种群问题,病原微生物问题,疾病的感染、传播和流行因素等问题(蒲蛰龙,1992)。昆虫疾病的流行因素主要有生物因素和气象因素,其中生物因素包括病原微生物的致病力和传染力、寄主昆虫的生理状态、生活习性和密度以及鸟类等昆虫天敌;气象因素包括阳光、温度、湿度、风、雨等(蒲蛰龙,1992)。本研究通过对五个地区CnmeGV 流行病学的调查,发现仅江门市恩平县采集的稻纵卷叶螟幼虫出现了因感染CnmeGV 而死亡的现象,而此地恰好是1979年首次发现该病毒的地区。此外,本次调查的稻纵卷叶螟的死亡率与以前报道的一致,表明该病毒与区域生态环境能够和谐共存。其他四个地区未检测到有CnmeGV 的存在,说明CnmeGV 没有流行到这些地区,从某个角度反应了该病毒的流行传播能力不是很强,可以进一步通过大范围的采样来调查分析。
References)
Ayres MD,Howard SC,Kuzio J,et al.The complete DNA sequence of Autographa californica nuclear polyhedrosis virus[J].Virology,1994,202:586-605.
Chai HN,Du YZ,Qiu BL,et al.Detection and phylogenetic analysis of Wolbachia in the Asiatic rice leafroller,Cnaphalocrocis medinalis,in Chinese populations[J].Journal of Insect Science,2011,2:1-13.
de Moraes RR,Maruniak JE,Funderburk JE.Methods for detection of Anticarsia gemmatalis nucleopolyhedrovirus DNA in soil[J].Applied and Environmental Microbiology,1999,65:2307-2311.
Escasa SR,Lauzon HA,Mathur AC,et al.Sequence analysis of the Choristoneura occidentalis granulovirus genome[J].The Journal of General Virology,2006,87:1917-1933.
Federici BA.Baculovirus Pathogenesis,in the Baculoviruses[M].Miller LK,Eds.Plenum:New York,1997.
Ferrelli ML,Salvador R,Biedma ME,et al.Genome of Epinotia aporema granulovirus(EpapGV),a polyorganotropic fast killing betabaculovirus with a novel thymidylate kinase gene[J].BMC Genomics,2012,13:548.
Guarino LA,Xu B,Jin J,et al.A virus-encoded RNA polymerase purified from baculovirus-infected cells[J].Journal of Virology,1998,72:7985-7991.
Harrison RL,Popham HJ.Genomic sequence analysis of a granulovirus isolated from the old world bollworm,Helicoverpa armigera[J].Virus Genes,2008,36:565-581.
Herniou EA,Olszewski JA,Cory JS,et al.The genome sequence and evolution of baculoviruses[J].Annual Review of Entomology,2003,48:211-234.
Hong XY,Ding JH.Agricultural Entomology[M].Beijing:China Agriculture Press,2007.[洪晓月,丁锦华.农业昆虫学[M].北京:中国农业出版社,2007]
Hu YY,Liu NC.The morphogenesis of Cnaphalocrocis medinalis after infected with two mixed viruses[J].Journal of Wuhan University,1982,04:101-128.[胡远扬,刘年翠.两种病毒混合感染稻纵卷叶螟及其形态发生[J].武汉大学学报,1982,04:101-128]
Jehle JA,Lange M,Wang HL,et al.Molecular identification and phylogenetic analysis of baculoviruses from Lepidoptera[J].Virology,2006,346:180-193.
Lange M,Wang HL,Hu ZH,et al.Towards a molecular identificationand classification system of lepidopteran-specific baculoviruses[J].Virology,2004,325:36-47.
Liang Z,Zhang X,Yin X,et al.Genomic sequencing and analysis of Clostera anachoreta granulovirus[J].Archives of Virology,2011,156:1185-1198.
O'Reilly DR,Miller LK,Luckow VA.Baculovirus expression vector:A laboratory manual[M].Plenum:New York,1992.
Pang Y,Lai YL,Liu J,et al.A new granulovirus from naturally-infected Asiatic rice leafroller,Cnaphalocrocis medinalis(Guenée)[J].Microbiology,1981,02:103-104.[庞义,赖涌流,刘炬,等.稻纵卷叶螟幼虫颗粒体病毒[J].微生物学通报,1981,02:103-104]
Pu ZL.Insect Pathology[M].Guangzhou:Guangdong Science and Technology Press,1992.[蒲蛰龙.昆虫病理学[M].广州:广东科技出版社,1992]
Pu ZL,Ye YC,Pang Y,et al.The Research on Insect Baculovirus.In:Reports of Science and Technology[M].Beijing:Scientific and Technical Documentation Press,1986:34-37.[蒲蛰龙,叶育昌,庞义,等.昆虫杆状病毒的研究.见:科学技术成果报告[M].北京:科学技术文献出版社,1986:34-37]
Rohrmann GF.Baculovirus Molecular Biology[M].National Library of Medicine(US),NCBI,Bethesda(MD),2013.
Rohrmann GF,Erlandson MA,Theilmann DA.The genome of a baculovirus isolated from Hemileuca sp.encodes a serpin ortholog[J].Virus Genes,2013,47:357-364.
Wormleaton S,Kuzio J,Winstanley D.The complete sequence of the Adoxophyes orana granulovirus genome[J].Virology,2003,311:350-365.
Tamura K,Dudley J,Nei M,et al.MEGA4:Molecular Evolutionary Genetics Analysis(MEGA)software version 4.0[J].Molecular Biology and Evolution,2007,24:1596-1599.
Zanotto PM,Kessing BD,Maruniak JE.Phylogenetic interrelationships among baculoviruses:evolutionary rates and host associations[J].Journal of Invertebrate Pathology,1993,62:147-164.
Zhang BQ,Cheng RL,Wang XF,et al.The Genome of Pieris rapae granulovirus[J].Journal of Virology,2012,86:9544.
Zhang XX,Geng JG,Lu ZQ,et al.Studies on the biological and ecological characters of the rice leafroller[J].Chinese Bulletin of Entomology,1980,06(2):241-245.[张孝羲,耿济国,陆自强,等.稻纵卷叶螟生物学生态学特性研究初报.昆虫知识,1980,06(2):241-245]
Zhang XX,Liang ZP,Yin XM,et al.Complete genome sequence of Agrotis segetum granulovirus Shanghai strain[J].Archives of Virology,2014,159(7):1869-1872.
Zhu Z,Yin FF,Liu XP,et al.Genome sequence and analysis of Buzura suppressaria nucleopolyhedrovirus:a group II Alphabaculovirus[J].PLoS One,2014,9:1-8.
