丝瓜三个果实性状及日开花时间性状遗传规律分析
2014-12-14杨寅桂周庆友庄泽彬汪国平
杨寅桂,周庆友,,张 娅,肖 遥,庄泽彬,汪国平*
(1.江西农业大学 农学院,江西 南昌 330045;2.华南农业大学 园艺学院,广东 广州 510642)
丝瓜为葫芦科(Cucurbitaceae)丝瓜属(Luffa spp.)一年生攀缘性草本植物,包括两个栽培种:普通丝瓜[Luffa cylindrical(L.)Rome.]和有棱丝瓜[Luffa acutangula(L.)Roxb]。丝瓜在我国南北均有栽培,我国长江流域及其以北各省区栽培普通丝瓜为主,而广西、广东及海南省则以栽培有棱丝瓜为主[1]。因丝瓜耐热、耐湿、适应性强,一直是我国重要的夏季蔬菜,其产品不但内销,而且大量出口港澳市场,是出口创汇的主要蔬菜品种之一[2]。
果实性状对丝瓜商品性有重要的影响。广东各地因消费习惯不同,对有棱丝瓜果色的要求也各有差异,如湛江、韶关、清远等地要求瓜果为赤麻色、大肉型,而珠江三角洲地区及港澳市场则要求深绿色、长条型[3]。两类瓜都要求首尾均匀,瓜把短或不明显。日开花时间则对产量有重要的影响,对于虫媒授粉植物来说,白天花朵开放,蜜蜂、蝴蝶等昆虫活动有利于及时授粉,提高座果率,而夜间昆虫不活动,花朵开放后得不到及时的授粉,从而致使座果率偏低。目前,丝瓜性状遗传研究与黄瓜、西瓜等重要葫芦科作物相比研究还不深入,果实性状遗传研究仅有少数报道[3-5],而日开花时间性状遗传分析暂时还无报道。本课题组近年对丝瓜主要性状的遗传规律开展了系统研究,本研究以有棱丝瓜和普通丝瓜为亲本构建种间群体,对3 个果实性状(果棱、果皮颜色、果把形状)及日开花时间性状的遗传规律进行了分析,以期为丝瓜遗传育种提供指导。
1 材料与方法
1.1 试验材料
本研究所使用的母本材料(P1)为广东省农科院蔬菜研究所品种“粤优大肉”后代高代选系,特点为有棱、果皮绿色、果把呈瓶颈形;父本(P2)为湖南株洲湘蔬种业有限公司“株杂二号白丝瓜”后代高代选系,特点为无棱、果皮白色、果把呈钝圆形。通过亲本材料的杂交、回交、自交,获得各世代种子。
1.2 试验方法
试验在华南农业大学跃进北蔬菜场进行,2010 年至2012 年春季获得各世代种子,包括:(P1×P2)F1、(P2×P1)F2、[(P1×P2)×P1]B1、[(P1×P2)×P2]B2,还有部分反交的(P1×P2)F’2、[(P2×P1)×P2]B’2。2012 年秋季栽植亲本及各世代材料,进行果实性状及日开花时间性状调查。亲本及F1在3 年间性状表现稳定,故2012 年秋茬未设置重复,各分离群体采取随机区组安排,双行种植,株距约40 cm,大行距约80 cm,小行距约60 cm。田间双行稀植并插直立人字架引蔓,其他栽培管理措施按照常规。

图1 丝瓜开花的鉴定标准Fig.1 The criterion of flowering in Luffa
盛瓜期对典型商品瓜鉴定果实性状。果棱、果皮颜色和果把形状的鉴定标准参照《植物新品种特异性、一致性和稳定性测试指南—丝瓜》(中华人民共和国农业部发布),并稍做修改。果棱性状分为3 个等级:无、中、强,分别赋值1、2、3;果皮颜色分为4 个等级:白色、黄白色、浅绿色、绿色,分别赋值为1、2、3、4;果把形状分为3 个等级:钝圆形、溜肩形、瓶颈形,分别赋值1、2、3。赋值方法参考程周超[6]。
丝瓜开花的衡量标准为花瓣张开,可以看到雄蕊(图1),于丝瓜开花盛期选择一天,从16:00(母本即将开花)起每隔2 h 调查田间开花植株直至第2 天06:00(父本已经开花),共调查7 次。根据鉴定结果,为了方便遗传分析,将日开花时间转换为数值,16:00—18:00 开花赋值为1,18:00—20:00 赋值为2,20:00—22:00 赋值为3,22:00—00:00 赋值为4,00:00—02:00 赋值为5,02:00—04:00 赋值为6,04:00—06:00 赋值为7。
采用SPSS 11.5 统计软件进行卡方测验分析。采用Excel 进行平均数、方差分析等。采用加性-显性遗传模型进行数量性状遗传分析,遗传模型适合性测验采用ABC 尺度法,选择最适遗传模型,并估算遗传参数[7]。
2 结果与分析
2.1 果棱性状的遗传分析
P1的果棱隆起0.5~1 cm,扎手感明显;P2果实无棱,表皮光滑,无扎手感;F1的果棱中等,F2果棱性状分离,从强到无棱各级都有出现,正交及反交的F2果棱性状分布趋势一致(表1),无棱∶弱棱∶强棱接近1∶2∶1,对正、反交F2群体进行卡方测验,F2群体χ2=0.567 4,F’2群体χ2=0.215 4,均小于=5.991;F1与有棱亲本回交,后代果棱性状偏向有棱亲本;F1及反交F’1与无棱亲本回交,后代果棱性状都偏向无棱亲本,不表现无棱∶弱棱为1∶1 的分离。此结果说明,果棱性状为质量性状,有棱对无棱为不完全显性。
2.2 果把形状、果皮颜色及日开花时间的遗传规律分析
果把形状F1为中间类型,F2明显偏离钝圆形,B1未出现钝圆形,说明钝圆形为隐性(表1);果皮颜色F1为浅绿,F2明显偏向绿色,B1未出现白色、黄白色,说明果皮绿色为显性(表1)。日开花时间P1为17:30—18:30,P2为5:30—6:30,F1为21:00—22:00,偏向于亲本P1,说明早开花为部分显性。3 个性状的正、反交F2间及正、反交F1作母本的回交群体间性状的表现呈一致趋势,说明不存在细胞质的影响。
3 个性状在F2群体中呈连续分布表现,因此将之判断为多基因控制的数量性状。
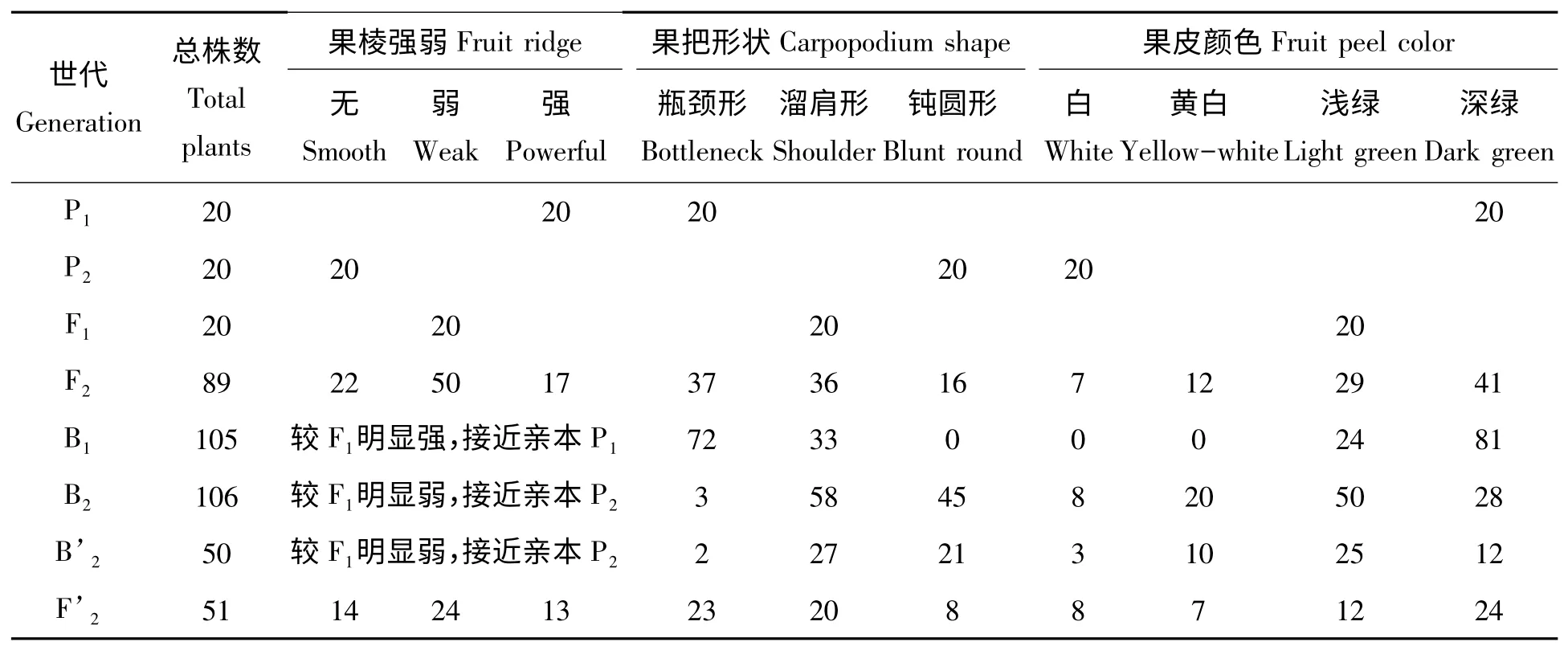
表1 丝瓜3 个果实性状调查结果Tab.1 The frequency distribution of fruit ridge,carpopodium shape and fruit color traits
2.2.1 遗传模型适合性检测 丝瓜各世代果皮颜色、果把形状、日开花时间的平均数和观察方差见表2。
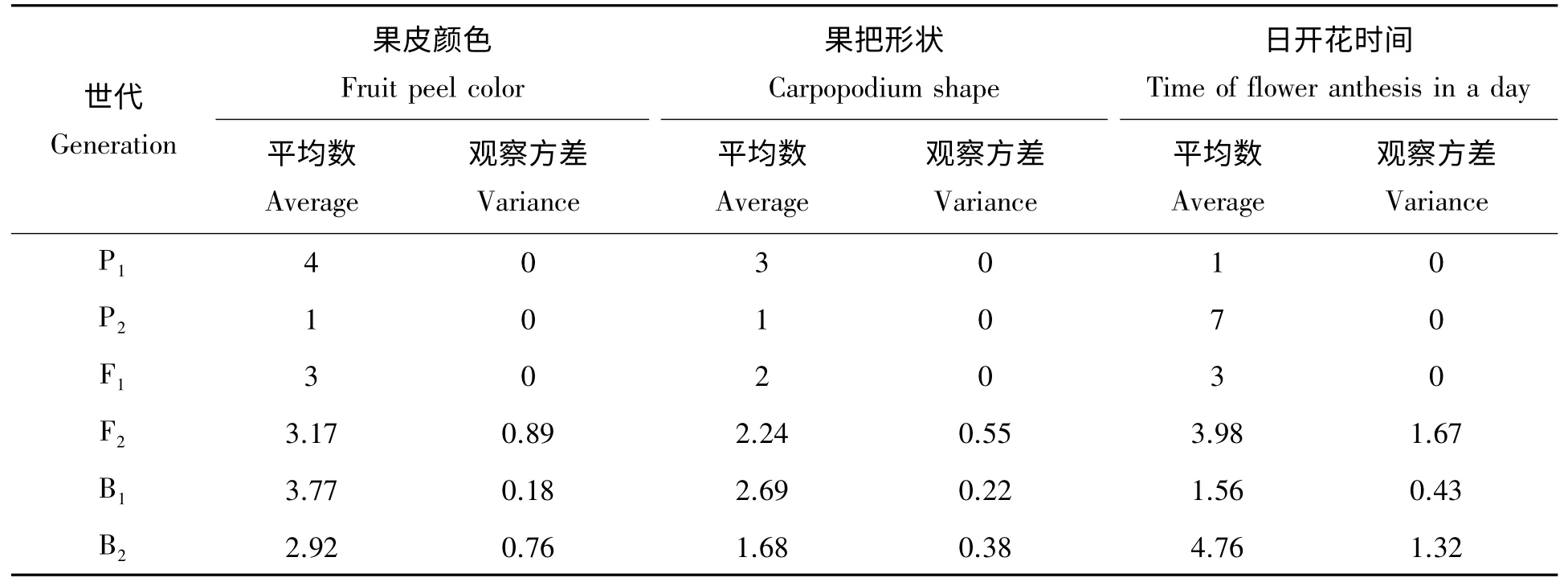
表2 丝瓜果皮颜色、果把形状、日开花时间平均数和方差分析Tab.2 The average and variance of fruit peel color,carpopodium shape and time of flower anthesis in a day traits of parents,F1,F2,B1and B2generations
从测量结果得到世代平均数和观察方差后,用加性-显性的遗传模型来估算各种遗传方差所占比重。针对所使用的遗传模型对丝瓜果皮颜色、果把性状和日开花时间性状的研究是否适合,采用ABC3种尺度测验。若符合加性-显性遗传模型,则A、B、C 值均应为0,若不符合该模型,存在上位性,则不等于0。ABC 尺度计算公式如下:
A 尺度检测:A=P1+F1-2B1,δA2=VP1+VF1+4VB1,t=A/δA;
B 尺度检测:B=P2+F1-2B2,δB2=VP2+VF1+4VB2,t=B/δB;
C 尺度检测:C=P1+P2+2F1-4F2,δC2=VP1+VP2+4VF1+16VF2,t=C/δC。
查t 值分布表得:当Df>120 时,t0.05=1.96,t0.01=2.57。由表3 可知,A、B、C 尺度t 值均小于1.96,即A、B、C 值与0 差异不显著,表明丝瓜果皮颜色、果把形状、日开花时间性状均符合加性-显性遗传模型,不存在上位性遗传。
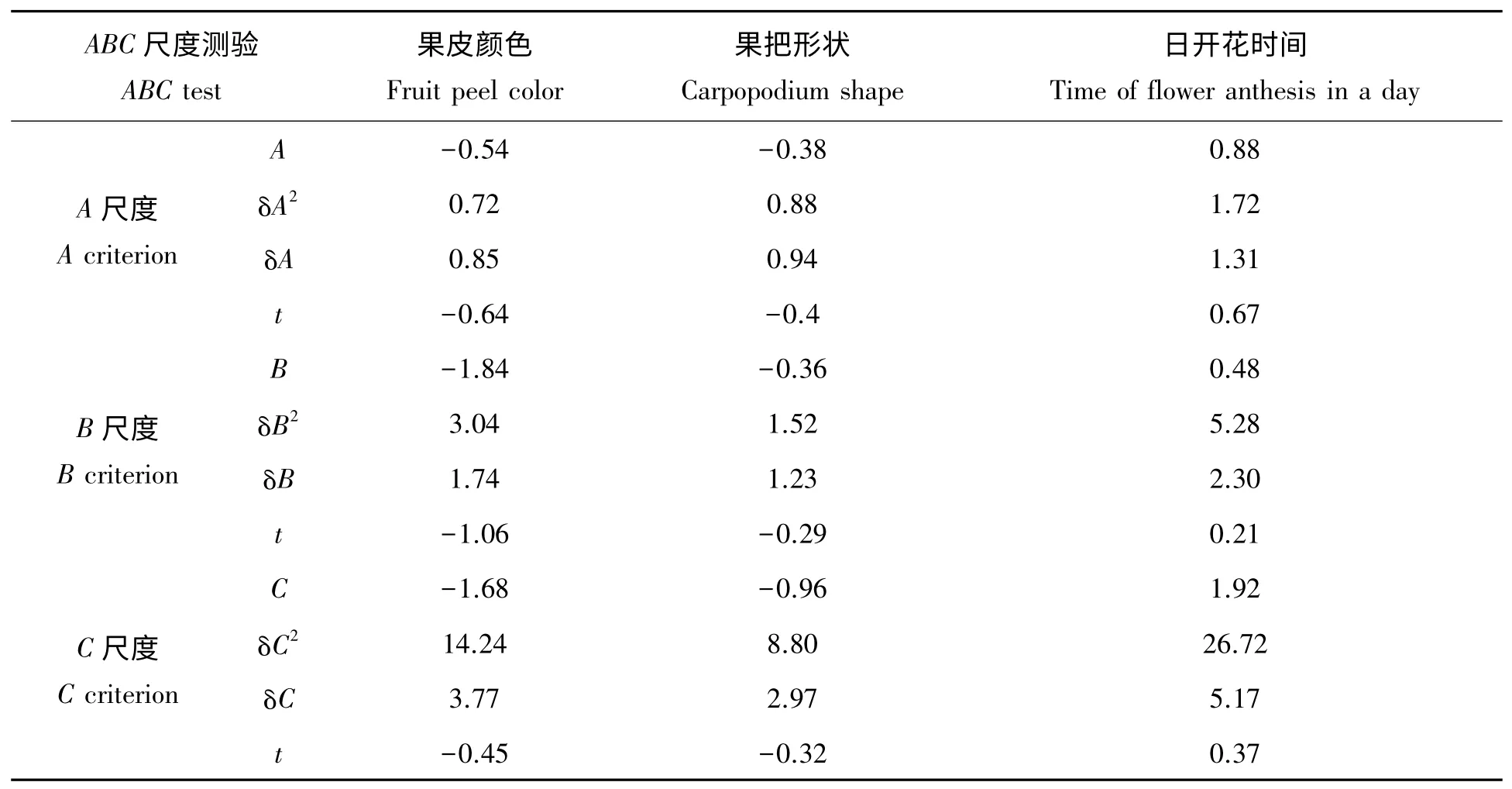
表3 丝瓜果皮颜色、果把形状、日开花时间ABC 尺度测验结果Tab.3 The ABC test of fruit peel color,carpopodium shape and time of flower anthesis in a day
2.2.2 遗传参数估计 根据加性-显性遗传模型,将总表型方差分解为加性方差Vd、显性方差Vh和环境方差Ve3 个组成部分,表型值分解为群体平均数μ、加性效应值d,显性效应值h 和环境差值e。由此可得VF2、VB、Ve、广义遗传力hB2、狭义遗传力hN2、显性度、显性遗传方差Vh、加性遗传方差Vd的计算公式(如下)。
VF2=0.5d+0.25h+Ve,VB=VB1+VB2=0.5d+0.5h+2Ve,Ve=(VP1+VP2+2VF1)/4,hB2(%)=(VF2-Ve)/VF2×100%,hN2(%)=(2VF2-VB)/VF2×100%,显性度==VF2+VB1+VB2+Ve,Vd=0.5d=2 VF2-(VB1+VB2)
已知VP1、VP2、VF1、VF2、VB1、VB2(表2),根据以上公式可计算出并列于表4 中。
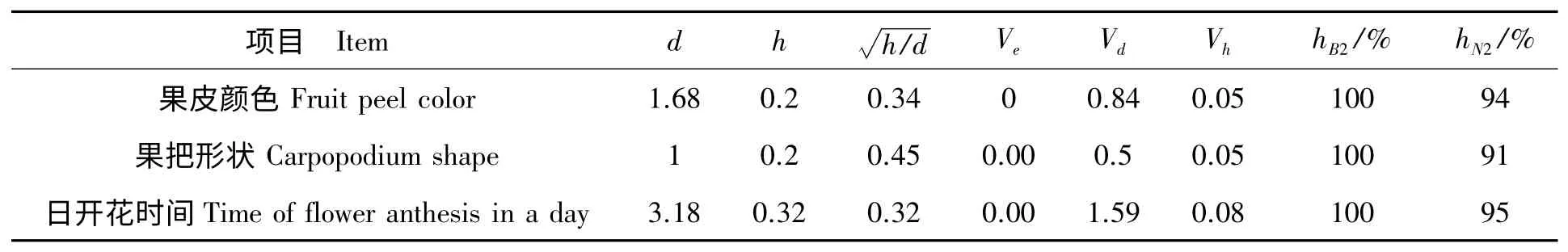
表4 丝瓜果皮颜色、果把形状、日开花时间遗传参数Tab.4 Genetic parameters of fruit peel color,carpopodium shape and time of flower anthesis in a day
由表4 可知,本试验果皮颜色、果把形状、日开花时间性状的环境方差都为0,广义遗传力hB2都为100%,即表明不考虑丝瓜单株所处环境的差异对表型值差异的影响,只考虑遗传原因决定这3 个性状的变异。狭义遗传力hN2果皮颜色性状为94%、果把形状性状为91%、日开花时间性状为95%,表明在遗传群体方差组成成分中,加性效应非常明显。显性度均小于1,表明该丝瓜组合果皮颜色、果把形状、日开花时间性状的遗传属于不完全显性遗传,基因加性效应明显。
2.2.3 加性基因数的估计 以公式K=(P1-P2)2/8Vd估计加性基因数,得出果皮颜色性状最少由2 个加性基因控制,果把形状性状最少由1 个加性基因控制,日开花时间最少由3 个加性基因控制。
3 讨论
3.1 丝瓜果棱性状的遗传
丝瓜果棱的强弱是有棱丝瓜育种中重要的选择性状,有棱丝瓜品种需要去皮棱食用,因而育种目标之一是浅棱,以减轻运输过程中因棱相互碰撞造成的果实损伤,而且方便去皮。本研究结果表明果棱由1 对主基因控制,有棱对无棱为部分显性,这一结果与Thakur 等[8]的结果一致,Thakur 将该基因命名为Ri;除主基因外,还存在微效调控基因,这一结论与生产上栽培的有棱丝瓜品种中棱的强弱存在一定变异相符合,推测不同有棱丝瓜品种中微效基因存在与否及多少调控主基因Ri 表达,从而表现出棱沟深浅的差异。
研究中发现B1群体和P1、B2群体和P2的果棱存在差异,但难于用手感及肉眼细分,F2的果棱出现各种级别的分离,感官分级时很难避免主观性,说明丝瓜果棱鉴定标准分为无、弱、强3 个等级比较粗糙。今后的研究应该想办法实现客观准确鉴定,以实现该性状的主基因+多基因的混合遗传模型分析。
3.2 丝瓜果皮颜色、果把及日开花时间性状的遗传
3.2.1 丝瓜果皮颜色、果把性状的遗传 丝瓜果皮颜色是外观品质的重要性状之一,不同地区的人因消费习惯的不同对丝瓜果皮颜色的要求也不同。林明宝[3]使用有棱丝瓜两个深绿果色的自交系UR1472 和CS2513 分别与两个赤麻果色自交系KN1318 和SZ1413 进行正交、反交、自交及回交后,对各世代果色的遗传表现进行分析,结果表明丝瓜赤麻果色的遗传受1 个显性核基因控制,为质量遗传;而王益奎等[5]认为有棱丝瓜中嫩瓜皮色深绿对灰绿为数量遗传。在生产实践中,丝瓜果皮颜色表现出由白到墨绿等多种变异,本实验中也观察到颜色深浅变异,因此判断丝瓜果皮颜色性状为多基因控制的数量性状,支持了王益奎等的观点。
丝瓜的果把即近瓜蒂端,瓜把的形状直接影响整个果实形状及果肉的食用率,育种目标是培育果把短或无、上下均粗的品种。在生产实践中,钝圆形(平肩)的品种少,有棱丝瓜还未见果把钝圆形的品种,这与本试验中观察到的钝圆形为隐性结果相符。
3.2.2 丝瓜日开花时间性状的遗传 自然条件下,有些植物的不同生态型或品种在同样的生长环境条件下的开花时间不一致。在日本同域分布的萱草属植物有日间开花型(Hemerocallis fulva,开花时间为04:30—07:30)和夜间开花型(Hemerocallis citrina,开花时间为16:30—20:30)之分,而它们的自然杂交种的开花则呈现不连续的双峰分布模式[9]。种植在同一条件下的蒲公英不同克隆植株之间也存在花期变异[10]。本试验对丝瓜杂交6 世代P1、P2、F1、F2、BC1、BC2的开花时间的次数分布进行分析,结果发现:P1的开花时间为17:30—18:30,P2的开花时间为05:30—06:30;F1的开花时间为夜间21:00—22:00,F2开花时间呈单峰分布,主要集中于20:00—22:00 开花;BC1集中开花时间为16:00—20:00,20:00以后有少量的花开放,BC2开花时间呈双峰分布,第一次集中开花的时间为22:00—00:00,第二次集中开花时间为02:00—04:00(图2)。这一发现与宋波[11]对丝瓜开花时间的观察结果基本保持一致,开花时间有所差异主要是因为调查的季节不同和调查的方式有差异。此外,还与萱草属植物的自然杂交种的开花分布结果保持一致。
植物开花受光、温度、水分、CO2、传粉媒介、植物激素及基因等因素影响[12]。目前,有棱丝瓜和普通丝瓜日开花时间差异大,出现这种现象的原因尚不明确,因此,后续工作可以开展寻找影响丝瓜日开花时间差异因素的研究。
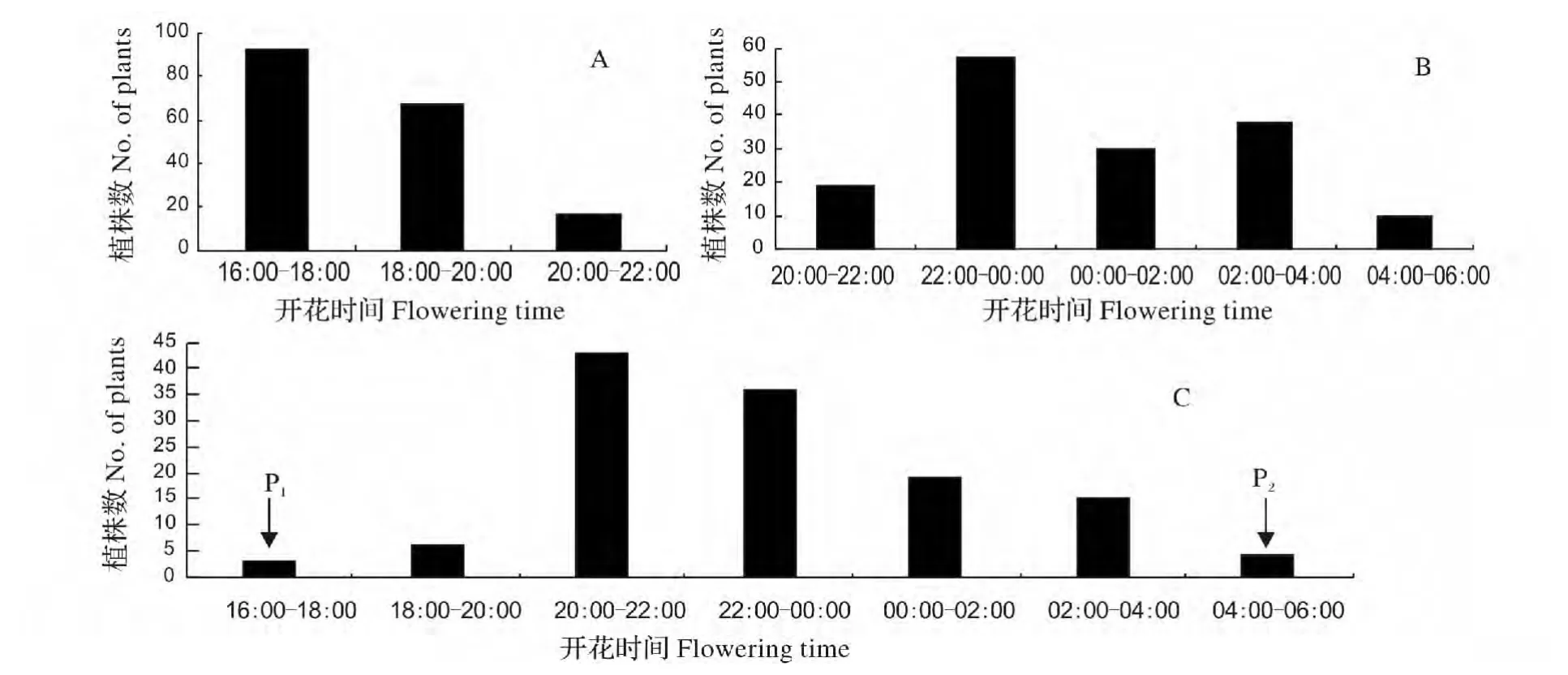
图2 丝瓜不同群体日开花时间的频数分布Fig.2 Frequency distribution of flowering time in a day of B1,B2 and F2 population in Luffa
丝瓜日开花时间鉴定比较辛苦,需要通宵连续观察,为方便夜间工作,防止邻近枝蔓的混淆,田间种植时需要注意稀植。为降低工作强度,试验曾采用将花枝采下浸泡于矿泉水中在实验室观察开花的方法,但发现部分花枝全天不能开放,表明该方法不稳定,最后本研究采用田间鉴定,每隔2 h 调查一次。丝瓜日开花时间呈连续分布,为数量性状,试验发现每隔2 h 调查一次的时间间隔偏大,后续工作中应该缩短,以获得更为准确详细的数据。
模型分析中亲本显性基因多少的信息,是选择亲本的重要参考之一[13]。本研究遗传分析表明,丝瓜果皮颜色、果把性状、丝瓜日开花时间均符合加性-显性遗传模型,遗传效应主要为加性效应,等位基因之间的交互作用所产生的显性效应甚微,无上位性效应,因此无法通过显性基因的多少来选择亲本;控制这3 种性状的最少基因数目分别为2 对、1 对、3 对。广义遗传力及狭义遗传力都非常高,环境因素引起的变异不作考虑,可固定的遗传成分占主导地位,因此在杂交育种中可在较早世代进行筛选,并且会有较好的选择效果[14-15]。
[1]王益奎,黎炎,李文嘉.我国丝瓜资源及遗传育种研究进展[J].北方园艺,2009(4):121-124.
[2]陈碧琳,邱汉林,叶晓青.岭南名优蔬菜栽培技术[M].广州:科学普及出版社,1989:162-163.
[3]林明宝,胡志群,林师森.有棱丝瓜果色遗传研究初报[J].广东农业科学,2000(2):16-17.
[4]苏小俊,徐海,高军,等.普通丝瓜果实性状的遗传分析[J].江苏农业学报,2009,25(5):1112-1118.
[5]王益奎,李文嘉,黎炎.有棱丝瓜相对性状遗传规律研究[J].长江蔬菜,2008(16):7-8.
[6]程周超.黄瓜SSR 遗传图谱的构建及黄瓜重要农艺性状的QTL 定位[D].北京:中国农业科学院,2010.
[7]马育华.植物育种的数量遗传学基础[M].南京:江苏科学技术出版社,1982:101-109.
[8]Thakur M R,Choudhury B.Inheritance of some qualitative characters in luffa species[J].Indian Journal of Genetics and Plant Breeding,1966,26(1):79-86.
[9]Hasegawa M,Yahara T,Yasumoto A.Bimodal distribution of flowering time in a natural hybrid population of daylily(Hemerocallis fulva)and nightlily(Hemerocallis citrina)[J].J Plant Res,2006,119:63-68.
[10]Collier M,Rogstad S.Clonal variation in floral stage timing in the common dandelion Taraxacum officinale(Asteraceae)[J].Am J Bot,2004,91:1828-1833.
[11]宋波.有棱丝瓜与普通丝瓜种间杂种后代的遗传分析[D].南京:南京农业大学,2008.
[12]罗睿,郭建军.植物开花时间:自然变异与遗传分化[J].植物学报,2010,45(1):109-118.
[13]侯广云,王文美,井立玲.小麦品种及突变体八个农艺性状的遗传模型分析[J].核农学报,1994,8(1):57-60.
[14]高国强,苏学合,曾昭海,等.陆地棉纤维品质和产量性状的遗传分析[J].核农学报,1998,12(4):193-199.
[15]高国强,孙永堂,苏学合,等.陆地棉优质新种质主要经济性状配合力分析[J].核农学报,2000,14(4):212-217.
