萌芽更新对桉树根际土壤微生物群落功能多样性的影响
2014-12-14宋贤冲杨中宁曹继钊张照远项东云
宋贤冲,杨中宁,曹继钊,张照远,项东云*
(1. 广西林业科学研究院 国家林业局中南速生材繁育实验室 广西优良用材林资源培育重点实验室,广西 南宁 530002;2. 广西国有七坡林场,广西 南宁 530225)
近年来世界范围内对木材需求大幅增加,森林已成为各国重要的战略资源,我国作为木材需求大国,对森林资源的依赖更为显著。桉树(Eucalyptus)由于具有速生丰产的优良特性,已成为我国速生丰产人工林的主要造林树种。截至2013年8月,广西全区桉树人工林面积已达到202.7万hm2,居全国桉树人工林种植面积首位[1]。如此大规模地种植桉树人工林,会对森林生态系统产生何种影响,也一直是困扰人工林生态系统研究者的一大问题。
根系是植物吸收水分和养分的器官,而根际又为土壤与植物生态系统物质交换的活跃界面,也是微生物发育的一个特殊生境,对外界环境极为敏感[2-4]。展小云等[5]利用 Biolog技术对小叶锦鸡儿根际土壤微生物群落功能多样性特征,及其对大气CO2浓度、土壤氮水平和土壤水分等环境因子变化的响应进行了研究,发现根际土壤微生物群落的功能在很大程度上受到外界环境因子的影响。近些年,随着人们对森林生态系统的重视,关于林木根际土壤微生物的研究日趋增多[6-8],但关于桉树根际土壤微生物的研究仍以少量分离培养的方法为主[9-11]。植物根际环境随着土壤类型、植物类型和生育阶段会发生很大的时空变异。杨远彪[10]利用稀释涂布平板法对连栽桉树根际微生物进行分析,结果表明二代林(一代萌芽更新林)土壤微生物数量最多,三代林(二代萌芽更新林)次之,一代林(新造林)最少。陈礼清等[12]对1 ~ 10年生巨桉(E. grandis)人工林土壤微生物数量进行研究,结果表明各微生物类群呈现1 ~ 4 a降低,而后显著增加。
本研究采取空间换时间的方法,选取广西不同萌芽更新代数的桉树人工林,通过对根际土壤微生物对Biolog Eco板碳源利用情况的测定分析,从不同碳源利用能力的大小来研究萌芽更新对桉树人工林根际土壤微生物功能多样性的影响。研究结果为揭示桉树不同萌芽更新代数下土壤微生物群落结构及其功能奠定基础。
1 材料与方法
1.1 采样点概况
采样点位于广西国有七坡林场七坡分场,地理位置为 22°32′ N,107°86′ E,属南亚热带季风气候,阳光充足,雨量充沛;地貌属于低丘地形,土壤类型为砂页岩发育形成的赤红壤,成土母岩为砂页岩。采样点林分均为2年生,分别为新造林、一代萌芽更新林和二代萌芽更新林。
1.2 土样的采集
2013年8—10月,分批次在上述采样点采集土壤样品。土壤采集方法参照林业行业标准LY/T 1852—2011,样品采集采取“之”字形 5点取样法,采集桉树根际土,每个点取样量大体一致,5个点的土样混成一个混合样。在树木周围多点挖取0 ~ 20 cm土层内的根系,先抖落根系上大块不含根系的土壤,然后取近根系表面的细粒土壤,装入塑料袋内混匀,作为根际土壤。
1.3 研究方法
微生物群落多样性测定应用Biolog方法,测定时间为2013年11月。10 g新鲜土壤加入100 mL无菌的0.145 mol·L-1NaCl溶液在摇床上振荡15 min,然后将土壤样品稀释至10-3,再从该悬浮液中取150 μL接种到生态板的每一个孔中,最后将接种好的板置于28℃的恒温培养箱中培养,每隔24 h在Biolog读数仪进行读数,培养时间共为144 h。
每孔的平均颜色变化率(Average well colour development, AWCD)计算方法如下[13]:
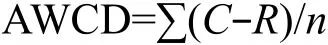
式中,C为每个有培养基孔的光密度值,R为对照孔的光密度值,n为培养基种类,Eco板n值为 31,C-R≤0的孔在计算中记为 0[14]。丰富度指数指被利用碳源的总数目,微孔的光密度值≥0.2,则认为是阳性值并计入微生物群落的丰富度S[15];多样性指数采用Shannon-Weinner指数(H')[16]:
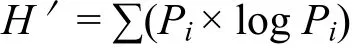
式中,Pi为有培养基的孔与对照孔的光密度值差与整板总差的比值,即:
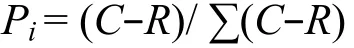
采取固定 AWCD值进行微生物代谢多样性类型分析,可以消除接种密度带来的差异[17]。因此,本研究采用培养 96 h的光密度值来进行主成分分析、单种培养基与主成分得分系数的相关分析、丰富度和多样性指数的分析。统计分析采用Excel和SPSS 17.0进行。
2 结果与分析
2.1 土壤微生物碳源平均颜色变化率(AWCD法)
Biolog Eco平板每个孔中的颜色变化率在一定程度上反映该碳源被微生物利用的情况,整个平板的 AWCD是反映微生物群落利用某一碳源能力的重要指标[16]。由图1可知,随着培养时间的延长,各样点土壤微生物利用碳源的能力呈上升趋势。各样点土壤微生物碳源 AWCD排序为一代萌芽林>二代萌芽林>新造林。
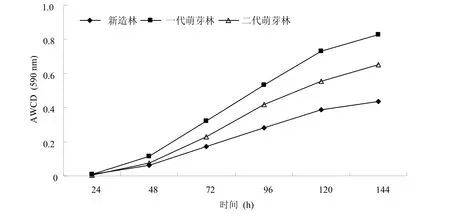
图1 不同萌芽更新代数桉树根际土壤平均颜色变化率
2.2 碳源利用类型的主成分分析
不同萌芽更新代数桉树根际土壤微生物群落代谢多样性类型存在显著差异。主成分分析结果表明,一代、二代萌芽林和新造林根际土壤中微生物群落具有明显的分异(图2),在主成分1(PC 1)方向有较大的分异(位于坐标轴右侧),而一代萌芽林和二代萌芽林之间分异不明显。新造林根际土壤微生物群落数据点较为集中,数据重现性好。
通过对根际土壤中与主成分1相关显著的碳源进行分析,结果表明对根际土主成分1起分异作用的主要碳源分别为羧酸类物质(4-羟基苯甲酸、α-酮丁酸、丙酮酸甲酯和衣康酸)、多聚物类(吐温40和α-环糊精)、氨基酸类(L-苯丙氨酸、L-天门冬酰胺、L-苏氨酸和L-丝氨酸)。
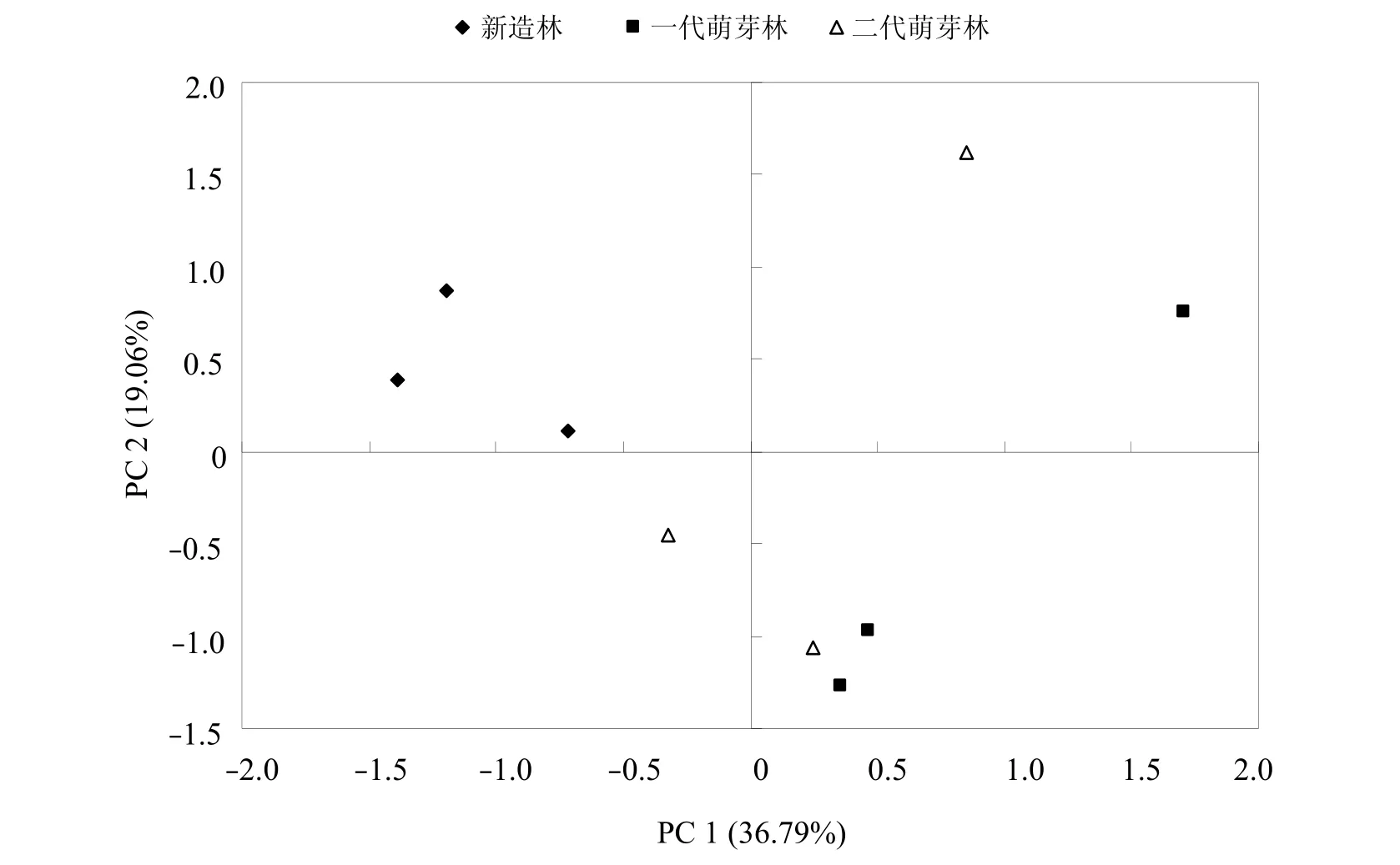
图2 不同萌芽代数桉树根际土壤微生物碳源利用类型的主成分分析
2.3 被利用碳源的丰富度和多样性指数
被利用碳源的丰富度和多样性指数,表示在平均颜色变化率一致的情况下土壤微生物群落利用碳源类型的多少,即功能多样性[17]。不同萌芽代数根际土壤中被利用碳源的丰富度和多样性指数之间的差异不显著(表1)。
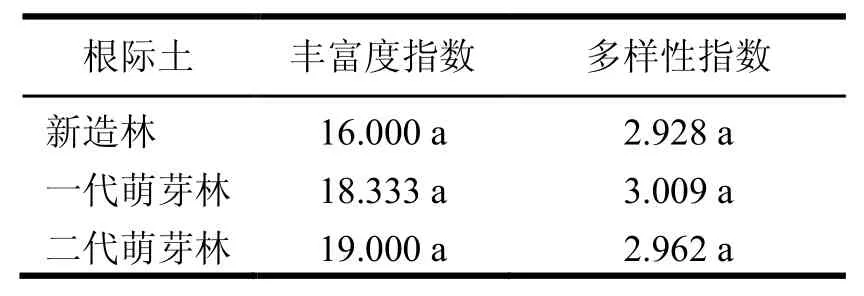
表1 根际土被利用碳源的丰富度和多样性指数
3 讨论
土壤微生物在生态系统的物质转化和能量传递过程中起着重要的作用,可作为衡量土壤健康程度的指标[18]。植物在生长过程中,根系不断延伸和凋亡,同时还分泌一些化合物,这些都为根际微生物的生长繁殖提供了天然的营养物质。本研究桉树根际土壤微生物利用碳源的总量,随着培养时间的延长呈逐渐增加的趋势,这与其他研究[5,16]结果一致。
植物根际环境随着土壤类型、植物类型和生育阶段会发生很大的时空变异。本研究结果与杨远彪[10]利用稀释涂布平板法对连栽桉树根际微生物的分析结果一致。造成上述差异的原因可能是萌芽林根系较新造林发达,微生物已经适应了根际微环境。
土壤微生物碳源利用类型的主成分分析可揭示不同样点利用碳源能力的分异程度。主成分分析结果表明,一代、二代萌芽林和新造林根际土壤中微生物群落在主成分1方向具有明显的分异,而一代萌芽林和二代萌芽林之间分异不明显,各样点在主成分2方向的分异不明显。由于各样点在主成分2方向分异不明显,故后续选取主成分1来比较相关显著的碳源。
不同萌芽代数桉树根际环境存在差异,其土壤微生物群落利用碳源类型也不同。张海涵等[19]研究了不同生态条件下油松(Pinus tabulaeformis)菌根根际土壤微生物群落特征,发现对安塞油松和商南油松菌根根际土壤微生物群落起分异作用的主要碳源分别为糖类、羧酸类和氨基酸;展小云等[5]对CO2浓度、氮素和水分处理下小叶锦鸡儿根际土壤微生物碳源利用情况进行主成分分析,结果表明与主成分相关显著的碳源类型主要是聚合物、糖类、氨基酸及少量的羧酸。本研究结果表明对根际土主成分1起分异作用的主要碳源中的4-羟基苯甲酸和氨基酸类是作物根系分泌物的主要成分[20]。由此推断桉树根际土壤微生物可能具有降解酚酸和自身代谢产物,避免自毒作用的能力;同时,氨基酸是土壤有机氮的重要组成部分,微生物在生长代谢过程中可利用氨基酸作为其氮源合成植物生长调节剂促进植物的生长[21]。综上,不同萌芽代数桉树根际土壤微生物群落利用碳源能力的差异与其利用根系分泌物和氨基酸等物质的差异有关。
被利用碳源的丰富度和多样性指数反映土壤微生物群落的组成结构和多样性。本研究结果表明不同萌芽代数根际土壤中被利用碳源的丰富度和多样性指数之间的差异不显著,这可能是因为不同萌芽代数间根际土壤微生物活力不同,但其微生物种类之间没有太大的差异。
致谢:感谢广西吴圩森林生态系统定位观测研究站、广西国有七坡林场提供林地支持;感谢广西大学生命科技学院冯家勋教授课题组在Biolog分析给予的指导和帮助!
[1] 陈少雄,陈小菲.我国桉树经营的技术问题与思考[J].桉树科技,2013,30(3):52-59.
[2] 林先贵.土壤微生物研究原理与方法[M].北京:高等教育出版社,2010.
[3] 曾曙才,苏志尧,陈北光,等.植物根际营养研究进展[J].南京林业大学学报(自然科学版),2003,27(6):79-83.
[4] Lynch J P.Root architecture and plant productivity[J].Plant Physiology,1995,109(1):7-13.
[5] 展小云,吴冬秀,张琳,等.小叶锦鸡儿根际微生物群落功能多样性对环境变化的响应[J].生态学报,2010,30(12):3087-3097.
[6] 李娇,蒋先敏,尹华军,等.不同林龄云杉人工林的根系分泌物与土壤微生物[J].应用生态学报,2014,25(2):325-332.
[7] 李海峰,刘岩,康颖,等.转基因小黑杨对土壤微生物群落结构的影响[J].南京林业大学学报(自然科学版),2014,38(2):75-80.
[8] 杨琼,谭凤仪,吴苑玲,等.不同林龄海桑林和无瓣海桑林根际微生物特征[J].生态学杂志,2014,33(2):296-302.
[9] 章家恩,刘文高,王伟胜.南亚热带不同植被根际微生物数量与根际土壤养分状况[J].土壤与环境,2002,11(3):279-282.
[10] 杨远彪.连栽桉树根际微生物分析及固氮菌的研究[D].南宁:广西大学,2008.
[11] 牛芳华,李志辉,周德明,等.尾巨桉幼苗根际土壤微生物分布特点及酶活性[J].中南林业科技大学学报,2011,31(3):151-155.
[12] 陈礼清,张丹桔,张健.巨桉人工林不同林龄阶段土壤微生物多样性特征[J].四川农业大学学报,2011,29(4):472-476.
[13] 陈晓倩,殷浩文.微生物群落多样性分析方法的进展[J].上海环境科学,2003,22(3):213-217.
[14] Garland J L.Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization[J].Soil Biology and Biochemistry,1996,28(2):213-221.
[15] Ratcliff A W,Busse M D,Shestak C J.Changes in microbial community structure following herbicide(glyphosate) additions to forest soils[J].Applied Soil Ecology,2006,34(2-3):114-124.
[16] 郑华,欧阳志云,王效科,等.不同森林恢复类型对土壤微生物群落的影响[J].应用生态学报,2004,15(11):2019-2024.
[17] 郑华,欧阳志云,方治国,等.BIOLOG 在土壤微生物群落功能多样性研究中的应用[J].土壤学报,2004,41(3):456-461.
[18] 任丽娜,王海燕,丁国栋,等.森林生态系统土壤健康评价研究进展[J].世界林业研究,2011,24(5):1-6.
[19] 张海涵,唐明,陈辉,等.不同生态条件下油松(Pinus tabulaeformis)菌根根际土壤微生物群落[J].生态学报,2007,27(12):5463-5470.
[20] 孔滨,杨秀娟.Biolog生态板的应用原理及碳源构成[J].绿色科技,2011(7):231-234.
[21] 张强,陈明昌,程滨,等.植物与土壤的氨基酸营养研究进展[J].山西农业科学,2001,29(1):42-44.
