STIM1在肿瘤中的研究进展*
2014-12-05徐金眉李小安刘卫华
徐金眉,周 艳,李小安,刘卫华
成都医学院第一附属医院 消化内科(成都 610500)
钙离子作为细胞生命活动中重要的信号分子,通过钙信号通路调控多种细胞功能,例如细胞分泌、神经激活、细胞凋亡、基因转录等[1]。几乎所有类型的细胞都要靠调节胞内Ca2+浓度来维持正常细胞活动。大量研究证实,肿瘤细胞中的Ca2+浓度远远高于正常细胞,且细胞膜上钙离子通道表达量增高,钙离子依赖的蛋白激酶如蛋白激酶C(protein kinase C,PKC)等在肿瘤细胞中活性也增高。肿瘤细胞中钙离子浓度稳定影响着肿瘤细胞的增殖、迁移等生命活动[2]。
细胞内Ca2+浓度调节主要依靠内质网中钙离子的释放和细胞外钙离子的内流。当贮存在细胞内质网中的Ca2+损耗后,细胞膜上的通道蛋白激活开放,使细胞外的Ca2+流向细胞内,以维持细胞内稳定的钙离子浓度。长期以来,存在着这样一个谜团:钙库中Ca2+浓度变化是如何影响细胞膜上的钙离子通道的。2005年Roos等[3]使用RNAi扫描技术筛选果蝇S2细胞的170个基因,其中有一个基因编码的蛋白能显著影响细胞内Ca2+的变化,这个蛋白就是基质相互作用蛋白分子1(stromal interaction molecule 1,STIM1)。STIM1位于细胞内质网膜上,感受内质网膜内钙离子浓度,是激活细胞膜上钙离子通道的“钥匙”。STIM1广泛存在于非兴奋细胞中,包括肿瘤细胞,它感受肿瘤细胞内Ca2+浓度变化,在调节肿瘤细胞生命活动方面发挥重要作用。
1 STIM1的结构
STIM1基因定位于人第11号染色体短臂第1区第5带第5亚带上(11p15.5),碱基长度约4 380bp,编码一种重约77KD的单次跨膜蛋白。STIM1的N端在内质网膜内,包括一个EF-hand、TM跨膜结构域、一个SAM序列。EF-hand是一个带负电荷的螺旋-环-螺旋模体,因此能结合钙离子,感受流动的钙离子浓度[4-5]。STIM1的C端定位于胞浆,包括CC1/CC2/CC3结构域,用于维持静息状态时STIM1的结构[3]。Orai1相互作用结构域-SOAR(STIM1orai activating region)或 者 叫 做CAD(CRAC activation domain)结构域,是通道CRAC的激活区域[6]。其他结构域还包括IDSTIM区、SHD 区、S/P(serine/proline-rich)区、Poly-K 尾巴。STIM1C端的200个氨基酸残基定位在其通道蛋白Orai1附近,但不能激活Orai1。钙离子流通过预测SOAR/CAD、SHD结构域分子变化来保持STIM1的活化或非活化状态[5]。
基质相互作用蛋白分子2(stromal interaction molecule 2,STIM2)一种与STIM1结构同源的蛋白,目前对STIM2的作用研究尚不深入且分歧颇大。有研究[7]认为它与STIM1一样有调节钙离子通道的作用;但还有人认为它在钙库操纵钙通道的调节中不起作用[8],甚至对STIM1起拮抗作用[9-10]。
2 STIM1与SOCC通道
细胞膜上主要的钙离子通道有:电压门控钙通道(voltage dependent calcium channel,VDCC),受体操纵钙通道(receptor operated calcium channel,ROCC),钙库操纵钙通道(store-operated Ca2+channel,SOCC)或钙释放激活钙通道(Ca2+releaseactivated Ca2+channels,CRAC)。VDCC是可兴奋细胞胞外钙离子进入细胞内的主要途径。ROCC的开放受某些神经递质,如谷氨酸、肾上腺素调节。它的配体受体结合,受体构型改变,使ROCC通道开放,Ca2+进入细胞内。由细胞内内质网钙池排空信号所激活和细胞膜表面SOCC开放所形成的钙库操纵性钙内流(store-operated Ca2+entry,SOCE)是非兴奋细胞内调节Ca2+稳态平衡的重要途径[8]。SOCE也是肿瘤细胞调节Ca2+水平的重要手段。
非兴奋细胞内质网中充满Ca2+,EF-hand一端结合Ca2+。细胞外信号分子激活细胞膜表面受体,活化PLC,使PIP2裂解为IP3和DAG。IP3与ER上的IP3受体相结合,激活内质网释放钙离子,内质网Ca2+浓度降低,Ca2+从 EF-hand脱离,位于内质网膜的STIM1被激活,STIM1活化后移动到细胞膜上Orai1通道蛋白一侧,与此同时,质膜上的Orail1蛋白自发组装成四聚体CRAC通道,有活性的STIM1二聚体激活CRAC通道,细胞外的Ca2+进入细胞内。当内质网中再充满Ca2+,EF-hand重新结合Ca2+,STIM1失活,CRAC通道关闭,Ca2+内流停止(图1)。
研究证明,抑制SOCC通道,破坏肿瘤细胞内钙离子稳态,能提高肿瘤细胞对化疗药物的敏感性。SOCE通过影响肿瘤细胞与细胞外基质相互作用,影响肿瘤的黏附、迁移和侵袭能力。早在40年前人们就发现了SOCE,但直到最近10年,人们才真正重视构成此通道的蛋白分子Orai1与STIM1。STIM1作为SOCE通道中感受钙离子浓度最关键的分子,它与肿瘤的关系受到广泛关注[12]。
3 STIM1与肿瘤
3.1 STIM1与肿瘤细胞的增殖与凋亡
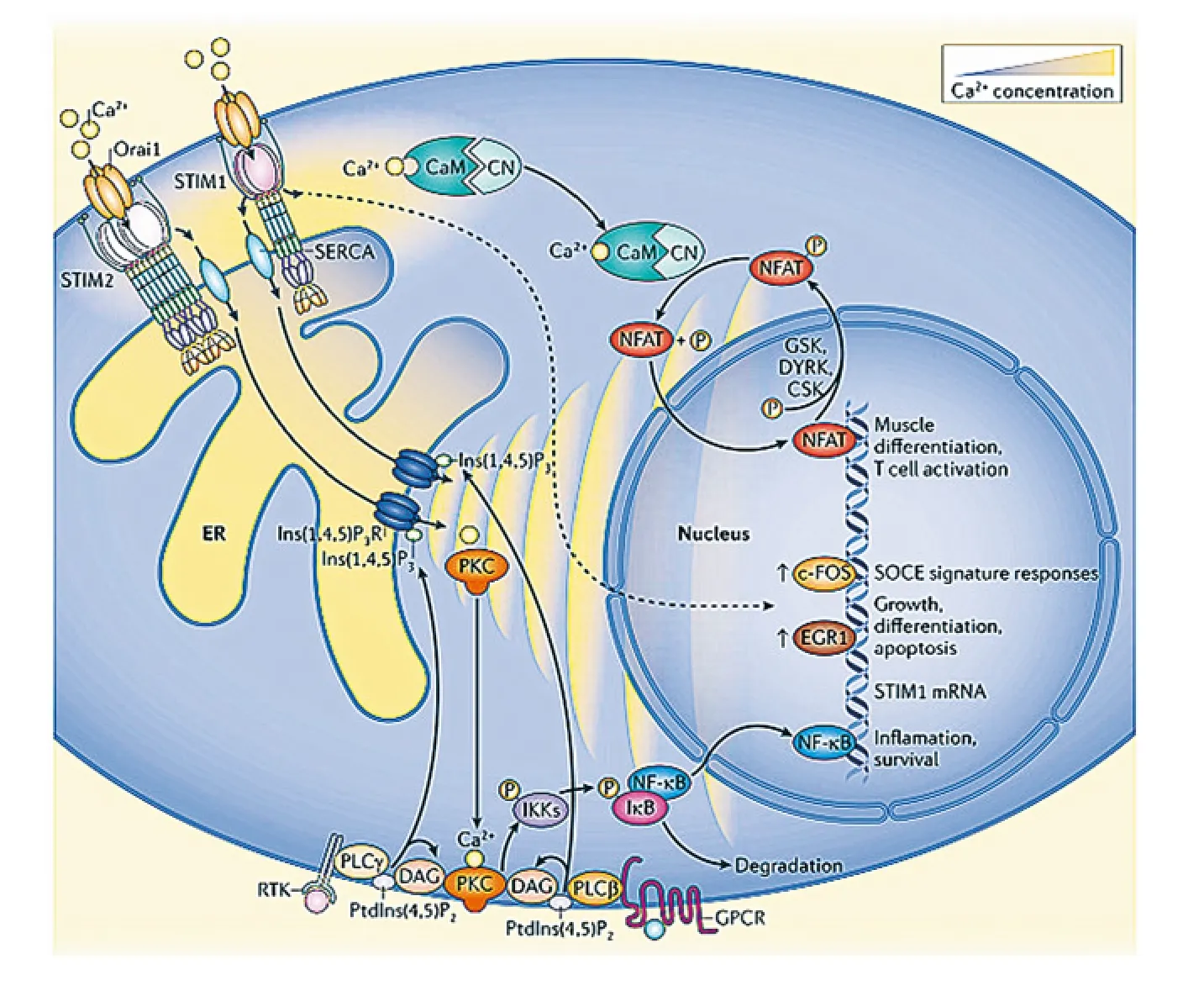
图1 STIM1及SOCE通道调节细胞内钙离子浓度[11]
科学家们对STIM1与肿瘤细胞相互作用的研究最早可以追溯到1997年,在对G401棒状肿瘤及横纹肌肉瘤细胞株的研究中发现,STIM1不仅能抑制瘤细胞生长,而且能引起细胞死亡[13]。这随即吸引了大量学者的目光,在接下来的十几年里,人们逐渐将研究重心从STIM1在Ca2+通道中的作用转移到它与肿瘤细胞的相互关系上来。Wenjun等[14]使用SOCE通道抑制剂2-APB和SKF96365实验发现,STIM1调控顺铂介导的非小细胞肺癌细胞凋亡;Sun等[15]在结肠癌细胞 HCT116中证明了,丁酸钠能促进细胞内钙池中的钙快速释放,使细胞处于预凋亡状态,使用SOCE通道抑制剂和STIM1干扰后,这种丁酸钠介导的结肠细胞凋亡减弱,而使用L型钙离子通道抑制剂处理后,不影响细胞凋亡。Chen等[16]在STIM1与宫颈癌关系的研究中,通过下调和上调STIM1表达,检测细胞增殖、凋亡以及细胞周期,发现抑制STIM1表达会抑制宫颈癌细胞增殖,使细胞周期阻滞在G1/S期或G2/M期。此时,检测到周期相关蛋白p21水平上升,而Cdc25c水平下降,但另一些周期蛋白,如cyclinA、cyclinB1、cdk1却没有受到STIM1的影响。研究者收集宫颈癌与癌旁临床标本,采用免疫印迹的方法检测出STIM1在癌中的表达高于癌旁。有趣的是,在分析标本资料时,他们发现STIM1的表达水平随肿瘤病灶增大而上升,而且STIM1在原发病灶的表达水平显著高于盆腔转移的淋巴结。在SCID小鼠的动物模型中STIM1也有促进肿瘤生长、促进血管生成和加深局部浸润的作用。虽然,这些研究都证明STIM1和其通道蛋白有促进肿瘤细胞增殖的作用,但是Yang等[17]在其文章中称STIM1和Orai1干扰乳腺癌细胞株 MDA-MB-231后并没有对细胞的增殖产生影响。STIM1对肿瘤细胞增殖产生差异性结果可能是因为STIM1在不同的细胞或组织类型中差异性表达所致。差异性表达使STIM1在宫颈癌等组织中表达高于乳腺癌,使其产生不同的细胞效应;或是因为组织和细胞类型不同,表达后修饰亦有差别。表达后修饰使STIM1在细胞类型中活性不同,进而影响其生理学作用;在不同组织和细胞中STIM1影响细胞增殖信号通路有所差别。肿瘤细胞内影响增殖的信号通路很多,它们相互交叉,相互影响。在不同细胞类型中STIM1可能是通过不同信号通路来影响细胞增殖的。目前,STIM1对肿瘤细胞增殖影响的机制尚不明确。STIM1是否有促进肿瘤增殖作用及其作用机制是一个很值得研究的问题。
3.2 STIM1与肿瘤细胞的黏附、侵袭与转移
肿瘤的浸润和转移是用于区分良恶性肿瘤的重要指标,直接影响临床治疗的效果和预后判断。临床上,肿瘤转移是治疗失败、患者死亡的主要原因。如何抑制肿瘤的浸润转移是肿瘤治疗研究的重点。在肿瘤细胞的迁移过程中,细胞的张力、形态与黏附性受细胞内钙离子浓度变化的影响,所以大量研究集中在STIM1对肿瘤细胞黏附、转移和侵蚀能力的影响上。
Yang等[18-19]用相同的方法分别在乳腺癌细胞系和肝细胞癌中证明了STIM1及其通道Orai1能减弱细胞间黏附,促进肿瘤细胞的侵袭和迁移。Chen等[19]在乳腺癌动物模型中证实,使用SOCE通道特异性阻滞剂SKF96365或STIM1shRNA干扰都能使肿瘤迁移能力减弱。Yang等[18]认为细胞迁移是一个需要从周围的细胞外基质分离再黏附的过程。他们使用细胞外基质(extracellular matrix,ECM)细胞黏附实验观察STIM1对肝癌细胞系HCC-LM3黏附力的影响。他们使用明胶、纤连蛋白和多聚赖氨酸包被培养板底,观察肝癌细胞黏附能力。对比发现STIM1基因干扰后,在涂有明胶和纤连蛋白的孔里细胞黏附能力增强,而在多聚赖氨酸包被的孔里黏附能力却没有受到影响。究其原因是纤连蛋白是一种高分子量的细胞外基质糖蛋白,通过结合整联蛋白和激动骨架蛋白,对细胞黏附、生长、迁移、分化产生重要作用;明胶是胶原的水解产物,它常用来涂于细胞培养板底,以改善细胞附着力;而多聚赖氨酸是靠物理作用吸附于孔底,利用带正电荷的亲水表面吸附细胞。纤连蛋白与明胶都是ECM成分,而多聚赖氨酸不是。这些都直接证明STIM1影响肿瘤的迁移可能是通过影响细胞从细胞外基质脱离实现的。
黏着斑是一种细胞的肌动蛋白细胞骨架与细胞外基质的连接方式[20]。Yang等[17-18]采用细胞免疫荧光方法,染色黏着斑的主要组成部分钮蛋白,观察钮蛋白变化情况。在对照组中钮蛋白呈点状网格状,干扰组或通道抑制组中钮蛋白体积变大,并环绕在细胞周围。转入GFP标记的桩蛋白载体动态观察结果发现,STIM1影响黏着斑组装和拆卸从而影响肿瘤细胞迁移。
表皮生长因子(epidermal growth factor,EGF)已被证实有促进肿瘤侵袭转移的作用。研究者为探究STIM1是否与EGF的促进肿瘤迁移能力有关,Chen等[16]下调STIM1的表达水平,transwell实验发现宫颈癌细胞迁移能力下降了40%~50%,剩下的细胞迁移能力对EGF刺激的敏感性也有所下降。相反,STIM1过表达能增强宫颈癌细胞的迁移与侵袭能力。EGF能提高钙蛋白酶活性,STIM1干扰后,EGF介导的钙蛋白酶激活能力却下降了70%~80%。这些都证明EGF的促进肿瘤迁移能力可能与STIM1有关。
Zhuang等[21]在研究中发现STIM1的富集受其 mRNA 的3′端非编码区(3'-untranslated region,3'-UTR)影响[22-24]。miR-195 影 响 3'-UTR,使STIM1mRNA很不稳定,然而HuR却使STIM1mRNA趋于稳定。有趣的是,异位miR-195过表达增强了STIM1mRNA与ardonaute蛋白复合物的相互作用,增强了STIM1mRNA与p-bodies的共区域化,使其RNA更易降解。HuR作用却恰好相反。随后,Ran通过观察受伤后皮肤裸露面积的实验发现,miR-195过表达抑制了STIM1的表达,从而抑制了皮肤细胞的迁移,进而影响皮肤愈合。
3.3 STIM1与肿瘤血管内皮细胞
肿瘤之所以可以迅速生长并出现转移取决于肿瘤新生血管的形成。在原发肿瘤的诱发下,促进血管生成因素与抑制血管生成因素间的平衡被打破,新生血管持续生长,大量异常血管生成。与正常的血管相比,肿瘤血管内皮不完整、与周细胞相互作用减弱,导致血管屏障功能受损,通透性改变。肿瘤血管呈螺旋形生长,生长迅速且形态异常,使其血供更为丰富。这些特点都有利于肿瘤的生长和远处转移。所以,抗血管生成可以有效抑制肿瘤生长和远处转移。
STIM1在调节各种肌细胞功能和心血管疾病的发生中发挥作用[25-26]。在大鼠颈动脉球囊拉伤模型中研究发现:新生血管内膜中的STIM1和Orail表达增多;沉默STIM1或抑制通道的激活,可以减少血管平滑肌细胞的增殖和新生血管内膜的形成。因此,研究者们猜测STIM1与血管内皮细胞和血管生成有关。大量研究结果证实,不同的内皮细胞如人脐静脉内皮细胞(HUVEC)和肺动脉内皮细胞(HPAEC)上均有STIM1和 Orai1的表达。Lodola等[27]从肾母细胞癌患者外周血中分离的内皮祖细胞(RCC-EPCs)与正常的EPC对比研究发现,SOCE有重塑和调控血管生成的作用。Shinde等[28]进一步研究发现,STIM1能耦合凝血酶受体,激活肌球蛋白轻链磷酸化,促进肌动蛋白纤维形成,使细胞间的黏附降低。由此说明STIM1分子有控制内皮屏障功能的能力。Chen等[19]建立人宫颈癌SiHa细胞移植瘤SCID小鼠模型,15d后,对照组小鼠腹腔新生血管数量明显多于SOCE通道抑制组和STIM1干扰组。Yang等[29]用毒胡萝卜素耗竭内皮细胞内钙离子,用凝血酶或VEGF激活SOCE通道,证明了SOCE对内皮细胞的增殖有重要作用。Abdullaev等[30]研究发现敲除STIM1基因可使内皮细胞增殖停滞在细胞周期生物S期或G2/M期,从而抑制内皮细胞增殖。
我们大胆猜测,STIM1促肿瘤侵袭转移的作用不仅与其改变肿瘤细胞的黏附有关,也可能与其影响肿瘤血管内皮细胞有关。STIM1介导的肿瘤转移与肿瘤血管内皮的关系如何,STIM1通过何种方式影响血管内皮增殖,是调节血管内皮成熟过程还是通过影响细胞因子变化间接调控血管内皮增殖,这些都是值得深究和解决的问题。
4 展望
虽然,STIM1对肿瘤细胞增殖作用和机制尚不清楚,但较明确的是其促肿瘤的转移侵袭和促进血管内皮增殖的作用。深入研究STIM1与肿瘤细胞相互作用及机制,有可能为肿瘤靶向治疗提供新的靶点。
[1]Berridge MJ,Bootman MD,Roderick HL.Calcium signalling:Dynamics,homeostasis and remodeling[J].Nat Rev Mol Cell Biol,2003,4(7):517-529.
[2]Zheng L,Stathopulos PB,Li GY,etal.Biophysical characterization of the EF-hand and SAM domain containing Ca2+sensory region of STIM1and STIM2 [J].Biochem Biophys Res Commun,2008,369(1):240-246.
[3]Roos J,DiGregorio PJ,Yeromin AV,etal.STIM1,an essential and conserved component of store-operated calcium channel function[J].J Cell Biol,2005,169(3):435-445.
[4]Stathopulos PB,Zheng L,Li GY,etal.Structural and mechanistic insights into STIM1-mediated initiation of storeoperated calcium entry[J].Cell,2008,135(1):110-122.
[5]Martin M,Rainer S,Marc F,etal.Ca2+release-activated Ca2+(CRAC)current,structure,and function,Cellular and Molecular Life Sciences[J].Cell Mol Life Sci,2012,69(24):4163-4176.
[6]Covington ED,Wu MM,Lewis RS.Essential role for the CRAC activation domain in store-dependent oligomerization of STIM1[J].Mol Biol Cell,2010,21(11):1897-1907.
[7]Muik M,Fahrner M,Derler I,etal.A cytosolic homomerization and a modulatory domain within STIM1C-terminus determine coupling to ORAI1channels [J].J Biol Chem,2009,284(13):8421-8426.
[8]Oh-Hora M,Yamashita M,Hogan PG,etal.Dual functions for the endoplasmic reticulum calcium sensors STIM1and STIM2in T cell activation and tolerance[J].Nat Immunol,2008,9(4):432-443.
[9]Lu W,Wang J,Peng G,etal.Knockdown of stromal interaction molecule 1attenuates store-operated Ca2+entry and Ca2+responses to acute hypoxia in pulmonary arterial smooth muscle[J].Am J Physiol Lung Cell Mol Physiol,2009,297(1):17-25.
[10]Soboloff J,Spassova MA,Hewavitharana T,etal.STIM2is an inhibitor of STIM1-mediated store-operated Ca2+Entry[J].Curr Biol,2006,16(14):1465-1470.
[11]Jonathan S,Brad SR,Muniswamy M,etal.STIM proteins:dynamic calcium signal transducers[J].Nat Rev Mol Cell Biol,2012,13(9):549-565.
[12]Hewavitharana T,Deng X,Soboloff J,etal.Role of STIM and Orai proteins in the store-operated calcium signaling pathway[J].Cell Calcium,2007,42(2):173-182.
[13]Sabbioni S,Barbanti-Brodano G,Croce CM,etal.GOK:a gene at 11p15involved in rhabdomyosarcoma and rhabdoid tumor development [J].Cancer Res,1997,57(20):4493-4497.
[14]Wenjun L,Minhong Z,Lei X,etal.The apoptosis of non-small cell lung cancer induced by cisplatin through modulation of STIM1 [J].Exp Toxicol Pathol,2013,65(7-8):1073-1081.
[15]Sun S,Li W,Zhang H,etal.Requirement for store-operated calcium entry in sodium butyrate-induced apoptosis in human colon cancer cells[J].Biosci Rep,2012,32(1):83-90.
[16]Chen YF,Chiu WT,Chen YT,etal.Calcium store sensor stromal-interaction molecule 1-dependent signaling plays an important role in cervical cancer growth,migration,and angiogenesis[J].Proc Natl Acad Sci USA,2011,108(37):15225-15230.
[17]Yang S,Zhang JJ,Huang XY.Orai1and STIM1Are Critical for Breast Tumor Cell Migration and Metastasis[J].Cancer Cell,2009,15(2):124-134.
[18]Yang N,Tang Y,Wang F,etal.Blockade of store-operated Ca2+entry inhibits hepatocarcinoma cell migration and invasion by regulating focal adhesion turnover [J].Cancer Lett,2013,330(2):163-169.
[19]Chen JP,Luan Y,You CX,etal.TRPM7regulates the migration of human nasopharyngeal carcinoma cell by mediating Ca(2+)influx[J].Cell Calcium,2010,47(5):425-432.
[20]Guo W,Giancotti FG.Integrin signalling during tumour progression [J].Nat Rev Mol Cell Biol,2004,5(10):816-826.
[21]Zhuang R,Rao JN,Zou T,etal.miR-195competes with HuR to modulate stim1mRNA stability and regulate cell migration[J].Nucleic Acids Res,2013,41(16):7905-7919.
[22]Eulalio A,Huntzinger E,Izaurralde E.Getting to the root of miRNA-mediated gene silencing[J].Cell,2008,132(1):9-14.
[23]Baltimore D,Boldin MP,O'Connell RM,etal.MicroRNAs:new regulators of immune cell development and function[J].Nat Immunol,2008,9(8):839-845.
[24]Leung AK,Sharp PA.MicroRNA functions in stress response[J].Mol Cell,2010,40(2):205-215.
[25]Lee KJ,Woo JS,Hwang JH,etal.STIM1negatively regulates Ca2+release from the sarcoplasmic reticulum in skeletal myotubes[J].Biochem J,2013,453(2):187-200.
[26]Fodor J,Matta C,Oláh T,etal.Store-operated calcium entry and calcium influx via voltage-operated calcium channels regulate intracellular calcium oscillations in chondrogenic cells[J].Cell Calcium,2013,54(1):1-16.
[27]Lodola F,Laforenza U,Bonetti E,etal.Store-Operated Ca2+Entry Is Remodelled and Controls In Vitro Angiogenesis in Endothelial Progenitor Cells Isolated from Tumoral Patients[J].PLoS One,2012,7(9):42541.
[28]Shinde AV,Motiani RK,Zhang X,etal.STIM1controls endothelial barrier function independently of Orai1and Ca2+entry[J].Sci Signal,2013,6(267):18.
[29]Yang IH,Tsai YT,Chiu SJ.Involvement of STIM1and Orai1 in EGF-mediated cell growth in retinal pigment epithelial cells[J].J Biomed Sci,2013,20:41.
[30]Abdullaev IF,Bisaillon JM,Potier M,etal.Stim1and Orai1 mediate CRAC currents and store-operated calcium entry important for endothelial cell proliferation [J].Circ Res,2008,103(11):1289-1299.
