西双版纳地区两种榕树上黄猄蚁生活习性调查
2014-11-25周树杰彭艳琼杨大荣
周树杰,王 波,彭艳琼,杨大荣*
(1.中国科学院西双版纳热带植物园,昆明 650223;2.中国科学院大学,北京 100049)
树栖性的蚂蚁能捕食植食性昆虫,从而降低植食昆虫对宿主植物的取食,为植物带来益处。植物也进化出许多适应性策略,例如提供食物或栖息场所来吸引蚂蚁(Janzen,1966),种间可建立紧密的互利关系(Davidson et al.,1989;Fiala et al.,1994;Pringle et al.,2011)。然而,蚂蚁也会捕食那些对植物有利的昆虫,比如传粉昆虫,在这样的情形下,蚂蚁的存在降低了植物的适合度,对于植物表现出不利的影响。栖息在植物上的蚂蚁与植物的关系呈现出非常复杂的模式(Schatz et al.,2006),两者间的相互作用模式关键在于蚂蚁的捕食对象以及对捕食者选择。因此,理解蚂蚁的捕食对象及食性对于理解蚂蚁及其宿主间的互作模式是至关重要的。
榕树是榕属Ficus 植物的总称,榕树与其传粉榕小蜂之间一一对应的专性共生关系使其成为了互利共生系统中最为特化的一个类群(Wiebes,1979;杨大荣等,1999;Weiblen,2000;Anstett,2001)。榕树的果实为热带雨林中许多生物(包括鸟类、哺乳类、昆虫等)提供了食物从而成为热带雨林的关键类群(杨大荣等,2004)。有关榕树的研究对于解释热带雨林生态系统维持机制以及保护政策的制定具有重要的意义。
榕树上栖息着数量众多,种类丰富的蚂蚁类群。蚂蚁在榕树上的活动会对榕蜂互惠系统产生复杂而多样的影响,这种影响即可能是正的也可能是负的(Schatz and Hossaert-Mckey,2003;魏作东等,2005)。前人研究发现榕小蜂是蚂蚁可靠的食物来源,以至于蚂蚁偏爱在榕树上筑巢(Schatz et al.,2008)。举腹蚁属的Crematogaster scutellaris 对传粉榕小蜂具有捕食上的偏好,而且对传粉小蜂的捕食效率显著高于对非传粉小蜂的捕食效率 (Schatz and Hossaert-Mckey,2003);而在聚果榕Ficus racemosa 上活动黄猄蚁则主要捕食非传粉榕小蜂,减少了非传粉小蜂的产卵,从而间接有利于传粉小蜂和榕树的互惠系统(魏作东等,2005)。此外,聚果榕上的黄猄蚁能够利用榕树的挥发物质来选择捕食环境,增加捕食榕小蜂成功的机率,而且这种能力是通过学习来获得的 (Ranganathan and Borges,2009)。黄猄蚁Oecophylla smaragdina 是热带和亚热带陆地和森林生态系统的优势物种,体型较大,具有很强的攻击性,是重要的捕食者,它们常常在树上捕食,并且能够利用树叶构筑蚁巢 (Holldobler,1983;Azuma et al.,2002)。在西双版纳除了聚果榕,其他的榕树上也有黄猄蚁的活动。但是,黄猄蚁的活动习性,以及怎样影响这些榕—蜂互惠系统仍不清楚。
本研究以西双版纳地区高榕和垂叶榕上的黄猄蚁为研究对象,通过野外实验以及行为测定的方式,对地区两种榕树上生活的黄猄蚁在三个不同季节的活动规律、蚁巢位置、食性、光选择偏好以及在不同季节和榕果不同发育时期对于榕小蜂的捕食情况进行了观察和实验测定。弄清黄猄蚁的生活习性,深入揭示黄猄蚁对于榕—蜂互惠系统的影响。
1 研究样地与方法
1.1 研究样地概况
中国科学院西双版纳植物园位于云南省南部勐腊县勐仑镇(21°55'N,101°15'E),海拔560-680 m 。地处热带北缘,为热带季风性气候,全年四季不分明,可分为雾涼季(11月至次年2月),干热季(高温少雨3-5月)和雨季(高温多雨6-10月)。年平均气温21.4-22.6℃年均降水量1,556.8 mm,平均相对湿度86%。
1.2 研究材料
高榕Ficus altissma 和垂叶榕Ficus benjamina 均为西双版纳地区较为常见的榕树种类,都为雌雄同株榕树,都隶属榕属Ficus,榕亚属Subgenus Urostigma,榕组Section Urostigma,环纹榕亚组Subsection Conosycea。高榕主要分布在亚洲南部在热带地区可以形成独木成林的独特景观,是热带和亚热带地区的绿化和观赏的主要树种(Corner,1965)。垂叶榕主要分布于广东、海南、广西、云南、贵州,在云南生于海拔500-800 m 湿润的杂木林中(张秀实等,1998)。在西双版纳热带植物园内选择有黄猄蚁活动的高榕、垂叶榕各三棵作为观察的样树。
1.3 研究方法
1.3.1 活动规律研究
在植物园内选择有黄猄蚁活动的垂叶榕和高榕作为观察对象进行定点观察,在黄猄蚁活动的路线上定点观察。每个小时观察5 min,记录黄猄蚁向上和向下运动的次数,作为黄猄蚁活动的指标。由于下雨时黄猄蚁活动停止,所以观察均在天晴时候进行,每个季节重复观察5 次,每次观察均记录当时的温度。观察时间2012年9月到2013年7月。
1.3.2 黄猄蚁筑巢位置
黄猄蚁蚁巢的观察实验,观察黄猄蚁在两种榕树上的筑巢位置和比较黄猄蚁在高榕和垂叶榕上蚁巢的特点并对其进行拍照。
1.3.3 黄猄蚁的食性研究
本文是针对添加有效时间标签来扩展的时态RDF数据模型,根据有效时间的现实意义分析时态RDF数据存在的不一致性,并对每一类的不一致性提出了修复的方法,针对执行变化操作时产生的不一致性,进行了预处理的研究,以保护时态RDF数据的一致性,并通过实验验证了可行性。
对于黄猄蚁食性的研究我们采用火腿肠、榕果和白糖各3 g 放置于直径10 cm的白色塑料盘内,在黄猄蚁活动的区域放置30 min 统计这段时间内进入盘内的黄猄蚁数量并观察其取食行为,三组实验共进行6 次重复。
1.3.4 黄猄蚁对光的反应
取30 头黄猄蚁放置在黑白箱中,10 min 后观察统计黄猄蚁在光亮处和黑暗处的数量以测定黄猄蚁对于光的偏好。
1.3.5 黄猄蚁对于榕小蜂的捕食
在这个研究的过程中我们主要采用观察的方法,首先是观察不同的季节黄猄蚁对于榕小蜂的捕食情况。我们在榕果发育的4个不同阶段:雌花前期,雌花期,间花期,雄花期。对每个阶段的榕果进行3 天连续观察,统计在每个阶段传粉榕小蜂和非传粉榕小蜂被捕食的数量。
2 结果与分析
2.1 黄猄蚁活动规律
黄猄蚁的活动随着季节的变化发生明显的变化,另外在不同的树种上的活动也存在一定的差异。黄猄蚁的活动变化按照观察的时间顺序结果如下:
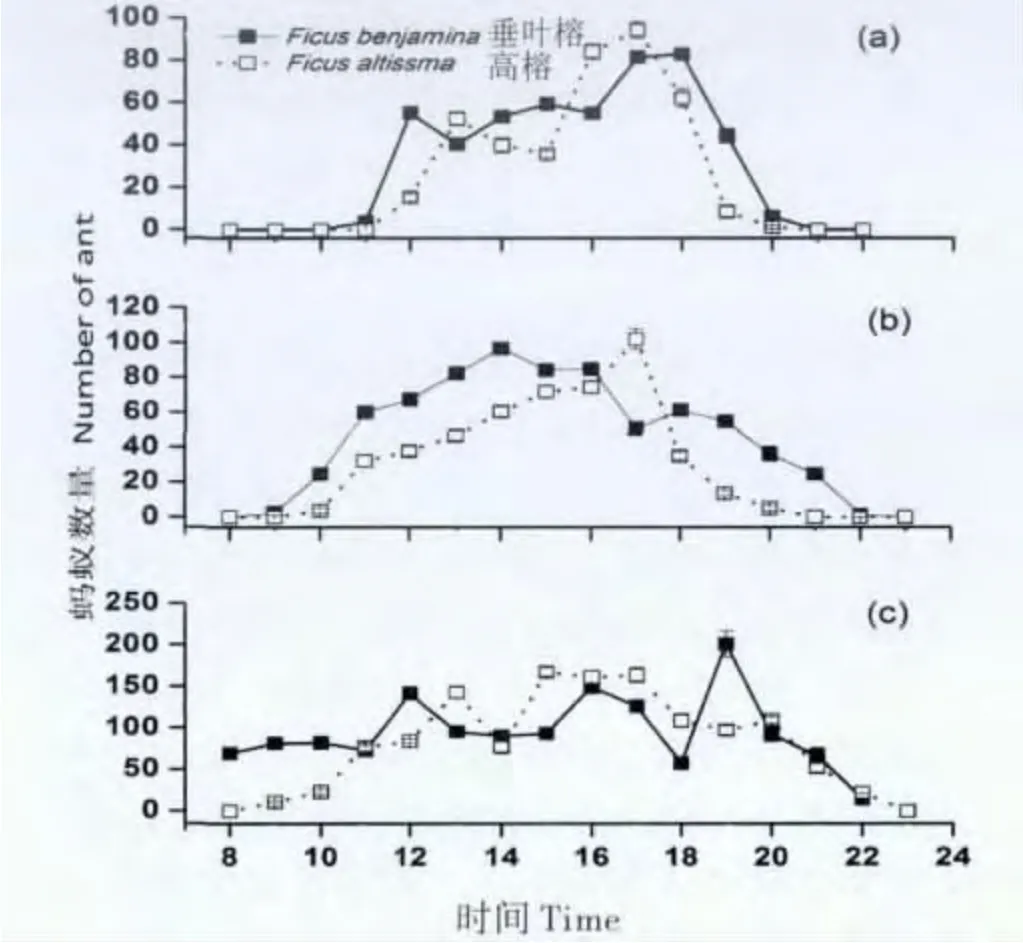
图1 不同季节黄猄蚁的活动规律。Fig.1 Oecophylla smaragdina’s activity pattern in different season
根据我们的观察结果黄猄蚁在三个季节的起始活动时间按时间的先后顺序依次为雨季>干热季>雾涼季,黄猄蚁活动时间的长短也是在雨季的活动时间最长15 h,在雾涼季的活动时间最短8 h,干热季的活动时间居于中间11 h。雨季垂叶榕上活动高峰为19 时,高榕高峰出现在17 时。雾涼季黄猄蚁在垂叶榕上活动的最高峰为18 时,高榕上黄猄蚁活动高峰17 时。干热季黄猄蚁活动介于雨季和雾涼季之间,垂叶榕黄猄蚁活动高峰14 时,高榕黄猄蚁活动高峰为17 时。记录的雨季平均温度25.05 ±0.76℃,雾凉季平均温度22.11±0.64℃,干热季平均温度29.41 ±0.50℃.
2.2 黄猄蚁的筑巢
生活在高榕和垂叶榕上的黄猄蚁,通过幼虫分泌的白色丝状粘液性物质将树叶粘合起来筑成蚁巢,在黄猄蚁蚁巢的上可以看到许多警戒的黄猄蚁工蚁。观察发现黄猄蚁在不同的榕树会选择不同的位置筑巢,在垂叶榕上,它们主要在树顶部筑巢,而在高山榕上则在杈下垂的小枝条上筑巢。

图2 垂叶榕和高榕上黄猄蚁的蚁巢Fig.2 Oecophylla smaragdina’s nest on the Ficus benjamina andFicus altissma
2.3 黄猄蚁的取食偏好
对实验数据进行单因素方差分析,结果表明:火腿肠诱饵的实验组与白糖组和榕果组之间存在极显著的差异(F2,15=111.65,P<0.001)。白糖组与榕果组之间没有差异。这说明黄猄蚁偏爱取食肉类食物,并不喜欢白糖和榕果食物,而且对后两种的取食没有差异(图3)。

图3 蚂蚁对不同种类食物的选择Fig.3 Ants’selectionon different kinds of food
2.4 黄猄蚁对光的反应
在光反应实验中,有26 头黄猄蚁选择光亮处,4 头黄猄蚁在黑暗处,Pearson 卡方的检验结果表明差异显著(χ2=16.13,P<0.001),说明黄猄蚁偏向选择有光的区域。另外我们发现黑暗处黄猄蚁活动较弱,光亮处的黄猄蚁活动性明显高于黑暗处。

表1 榕果不同发育时期黄猄蚁对榕小蜂的捕食Table 1 Oecophylla smaragdinas’prey on fig-wasps at the different fig phase
2.5 黄猄蚁对于榕小蜂的捕食
观察也发现黄猄蚁对于榕小蜂的捕食在季节上的变化与黄猄蚁的季节性活动变化一致,黄猄蚁活动的强度越大它对于榕小蜂的捕食也就越多。黄猄蚁对于榕小蜂的捕食强度随季节的变化情况为雨季>干热季>雾凉季。
3 结论与讨论
在我们的观察中黄猄蚁的起始活动温度一般不低于20℃,另外当温度高于32℃时黄猄蚁的活动也会明显的减弱。它们的最适活动温度在24-28℃之间,所以雨季黄猄蚁的活动强度最强,而在雾凉季和干热季活动较弱。在以上的实验观察结果在高榕和垂叶榕两种榕树上都有相同的结果,但是在细微的方面在垂叶榕上的黄猄蚁起始活动时间早于高榕,一般提早1 h,活动持续时间也长于高榕,相同时间活动的强度与大于高榕,我们进一步的观察发现造成以上结果的因素主要为人为干扰,垂叶榕样树位于道路旁,树不及高榕高大,人类的活动对于黄猄蚁的活动产生很大的影响,高榕样树受到的人为干扰较少。对于人类的活动具体是怎样影响黄猄蚁的活动还有待于进行具体的研究。
黄猄蚁以树叶为材料筑巢于树枝上,黄猄蚁幼虫能够分泌具有粘性的纤维把树叶粘合起来,黄猄蚁分泌的这种纤维的平均直径 (39.0 ±9.8um),表面密度为(0.22 ±0.03 g/cm3),这种纤维具有很好的粘合性(Siri and Maensiri,2010)。垂叶榕上黄猄蚁的筑巢位置一般较高,而高榕上蚁巢的位置高低都有,这可能与树叶的大小有关。垂叶榕叶子较小,因此垂叶榕上的蚁巢一般位于树叶密集的顶部。高榕叶片较大便于黄猄蚁筑巢。另外在西双版纳地区除了黄猄蚁以外很多其他种和蚂蚁也筑巢于树上或高处。热带雨林的降水量比较大而且比较集中2011 和2012年雨季6、7、8、9 四个月的平均降雨量为194.2 mm 和238.2 mm。因为降雨量很大很集中所以这些蚂蚁选择较高的地方筑巢就可以避免筑巢被雨水淹没,保证了蚁巢的安全。另外黄猄蚁的蚁巢并不会一直的使用,每隔一段时间就会有新建的蚁巢,另有一些蚁巢将会被弃用,对于其中的原因目前还没有相关的研究。我们在观察中发现雨季末是黄猄蚁筑巢的高峰时段,这个时间可能是黄猄蚁繁殖和分巢的活跃时期所以筑很多的新蚁巢。
黄猄蚁是典型的肉食性昆虫并且能够捕食许多种节肢动物,在我们的实验中再次证明了这一点。在以火腿肠为诱饵的实验组黄猄蚁的数量明显多于其他的组,而且黄猄蚁有明显的取食行为。在其他两组都只有少量的黄猄蚁,这与黄猄蚁的肉食性食性相关。
黄猄蚁对光反应的实验,通过比较发现黄猄蚁明显更偏向于有光的一侧(χ2=16.13,P<0.001)。但是黄猄蚁对于光的喜好与其他一些种类昆虫的趋光性有着明显的不同,在我们夜间的观察中发现光亮区域没有发生黄猄蚁的聚集,因此可以认为黄猄蚁的趋光性与一般昆虫的趋光性不同,只是趋向于热光源,它可能是利用光产生的热量进行自身和蚁巢的除湿。黄猄蚁的活动性与光照有明显关系,观察发现光亮处黄猄蚁的活动性明显高于黑暗处黄猄蚁的活动性。黄猄蚁的这种光选择性可能决定了黄猄蚁集中在白天活动,大部分的榕小蜂也是白天活动,黄猄蚁的活动时间也便于其进行捕食。少数种类的榕小蜂选择在夜晚进行产卵活动,可能就是要避免黄猄蚁的捕食(Ranganathan et al.,2010)。
在许多榕树种类上(主要是雌雄异株榕树)榕小蜂是黄猄蚁的主要食物来源(Schatz et al.,2008),在高榕和垂叶榕上也是黄猄蚁的重要食物来源。黄猄蚁对榕小蜂的捕食随季节的变化明显是与黄猄蚁自身的活动相关。在雨季黄猄蚁的活动最活跃对榕小蜂的捕食强度也是最大的,在雾凉季黄猄蚁的活动最弱对于榕小蜂的捕食也是最弱的,干热季的情况介于两者之间。黄猄蚁捕食榕小蜂的种类和榕树榕果生长阶段密切联系,非传粉榕小蜂在花前期和间花期数量较多,这就提供了黄猄蚁捕食非传粉榕小蜂的机会,所以在花前期和间花期观察到黄猄蚁捕食非传粉榕小蜂而没有发现其捕食传粉榕小蜂。在雌花期传粉榕小蜂通过苞片口进入榕果,雄花期成熟榕小蜂从榕果中飞出,这两个时期就给黄猄蚁捕食传粉榕小蜂的机会。我们可以推想出在花前期和间花期黄猄蚁以捕食非传粉榕小蜂为主,而在雌花期和雄花期以捕食传粉榕小蜂为主,所以榕树上栖息的黄猄蚁在花前期和间花期对榕树-榕小蜂互利共生有利,而雌花期和雄花期由于取食榕小蜂,则可能对榕蜂互利共生产生负面影响。
我们对于黄猄蚁相关生活习性的研究,目的在于为进一步开展黄猄蚁与榕蜂互惠系统相关研究打下一个基础,下一步的研究重点将集中于黄猄蚁在榕树上的活动会对榕蜂互惠系统产生什么样的影响,已及这种影响是通过怎样的途径实现的。在之前有关黄猄蚁与其他传粉系统相互影响的研究中发现,野牡丹Melastoma malabathricum 可以通过吸引黄猄蚁来驱赶体型较小的无效传粉蜂类,而不影响体型较大的同时也是其有效传粉者的木蜂属Xylocopa 蜂类。结果显示:相对于没有黄猄蚁的野牡丹花朵,被黄猄蚁占据的花朵具有更高的繁殖成功率。(Francisco et al.,2013)黄猄蚁捕食传粉昆虫,减少了传粉昆虫对于植物的访问,导致了传粉效率下降。(Kazuki et al.,2004)另外有研究观察了黄猄蚁对澳大利亚热带地区和西双版纳地区32个科的48 种植物的拜访情况,并记录他们与访花昆虫的相互关系。发现在其中的31 种植物上黄猄蚁攻击访花昆虫的情况,并在15种植物上成功捕捉到它们。黄猄蚁可以通过合作一起捕捉到比自己体型更大的蜂类。这项调查暗示了在热带亚洲地区黄猄蚁的存在很可能显著地影响了很多植物传粉者构成。(Miguel et al.,2013)由此可见黄猄蚁与许多的植物昆虫传粉系统都存在着密切的联系,黄猄蚁的活动对于相关发现系统产生重要的影响,因此研究黄猄蚁对于榕蜂互惠系统的影响,将对了解榕蜂互惠系统产生巨大的意义。
References)
Anstett MC.Unbeatable strategy,constraint and co-evolution,or how to resolve evolutionary conflicts:the case of fig/wasp mutualism[J].Oikos,2001,(95):476-484.
Azuma N,Kikuchi T,Ogata K,et al.Molecular phylogeny among local populations of weaver ant Oecophylla smaragdina [J].Zoological Science,2002,(19):1321-1328.
Corner EJH.Checklist of Ficus in Asia and Australasia with keys to identification [J].The Gardens’ Bulletin Singapore,1965,(21):1-186.
Davidson DW,Snelling RR,Longino JT.Competition amongants for myrmecophytes and the significance of plant trichomes [J].Biotropica,1989,(21):64-73.
Fiala B,Grunsky H,Maschwitz U,et al.Diversity of ant-plant interactions-Protective efficacy in Macaranga species with different degrees on ant association [J].Oecologia,1994,(97):186-192.
Francisco GG,Luis S,Richard TC,et al.Flowers attract weaver ants that deter less effective pollinators[J].Journal of Ecology,2013,(101):78-85.
Holldobler B.Territorial behavior in the green ant (Oecophylla smaragdina)[J].Biotropica,1983,(15):214-250.
Janzen DH.Coevolution of mutualism between ants and acacias in Central America [J].Evolution,1966,(20):249-275.
Miguel A,Rodriguez G,Francisco GG,et al.Possible role of weaver ants,Oecophylla smaragdina,in shaping plant-pollinator interactions in South-East Asia [J].Journal of Ecology,2013,(101):1000-1006.
Pringle EG,Dirzo R,Gordon DM.Indirect benefits of symbiotic coccoids for an ant-defended myrmecophytic tree [J].Ecology,2011,(92):37-46.
Ranganathan Y,Borges RM.Predatory and trophobiont-tending ants respond differently to fig and fig wasp volatiles [J].Animal Behavior,2009,(77):1539-1545.
Ranganathan Y,Ghara M,Borges RM.Temporal associations infig-wasp-ant interactions:diel and phenological patterns [J].The Netherlands Entomological Society,2010,(137):50-61.
Schatz B,Hossaert-Mckey M.Interaction of the ant Crematogaster scutellaris with the fig-fig wasp mutulism [J].Ecology Entomology,2003,(28):359-368.
Schatz B,Kjellberg F,Nyawa S,et al.Fig Wasps:A staple food for ants on Ficus [J].Biotroic,2008,(40):190-195.
Schatz B,Proffit M,Rakhi BV,et al.Complex interaction on fig trees:ants capturing parasitic wasps as possible indirect mutualists of the fig-fig wasp interaction [J].Oikos,2006,(113):344-352.
Siri S,Maensiri S.Alternative biomaterials;Natural,non-woven,fibroin-based silk nonfibers of weaver ants (Oecophylla smaragdina) [J].Internation Journal of Biological Macromolecules,2010,(46):529-534.
Tsuji K,Hasyim A,Ahsol H,et al.Asian weaver ants,Oecophylla smargdina,and their repelling of pollinators [J].Ecological Research,2004,(19):669-673.
Wei ZD,Peng YQ,Yang DR,et al.Impact of Oecophylla smaragina on the percentage number of offspring of pollinator and non-pollinating wasps on Ficus racemosa [J].Zoological Research,2005,(26):386-390.[魏作东,彭艳琼,杨大荣,等.聚果榕上黄猄蚁对传粉小蜂和非传粉小蜂后代数量的影响[J].动物学研究,2005,(26):386-390]
Weiblen GD.Phylogenetic relation of functional dioecious Ficus(Moraceae)based on ribosomal DNA sequence variation and morphology [J].American Journal of Botany,2000,(87):1342-1357.
Wiebes JT.Co-evolution of figs and their insect pollinator [J].Annual Review of Ecology and Systematics,1979,(10):1-12.
Yang DR,Li CD,Han TB.The impact of tropical rainforest fragmentation on figs and fig wasps [J].Zoological Research,1998,(20):126-130.[杨大荣,李朝达,韩灯保.热带雨林片段化对榕小蜂和榕树物种的影响[J].动物学研究,1998,(20):126-130]
Yang DR,Xu L,Peng YQ,et al.Species composition and diversity of fig wasps and figs in Yunan [J].Biodiversity Science,2004,(12)6:611-617.[杨大荣,徐磊,彭艳琼等.云南省榕小蜂和榕树的物种组成及多样性[J].生物多样性,2004,(12):611-617]
Zhang XS,Wu ZY,Cao ZY.Flora of China,Vol.23,Issue 1,Angiosperm,Dicotyledoneae,Moraceae [M].Beijing:Science Press,1999,104-106.[张秀实,吴征镒,曹子余.中国植物志,第23 卷,第1 分册,被子植物门,双子叶植物纲,桑科[M].北京:科学出版社,1999,104-106]
