贵州白背飞虱灯下种群发生动态区域性比较研究
2014-11-25薛文鹏金道超
薛文鹏,金道超,杨 洪
(贵州山地农业病虫害重点实验室,贵州大学昆虫研究所,贵阳 550025)
白背飞虱Sogatella furcifera (Horváth)(White-Backed Planthopper,WBPH)是重要的水稻害虫,在东亚、南亚和东南亚广泛分布危害。自上世纪70年代以来白背飞虱继褐飞虱Nilaparvata lugeh (Stål)(Brown Planthopper,BPH)后呈现出猖獗肆虐的发生趋势 (Cheng,2009;翟保平,2011),在我国各稻区均有发生,其终年繁殖的地区仅限于海南岛和云南省最南部,国内其它区域鲜有越冬场所(全国白背飞虱科研协作组,1981;四川、贵州、云南三省稻飞虱科研协作组,1982;胡国文等,1988;秦厚国等,2003),多从境外迁飞而来。因此灯下监测成为了解其种群发生动态的重要手段,利用白背飞虱长翅型成虫的趋光性,灯光诱集能及时掌握其始见日、高峰期、发生量的季节性动态变化等相关信息 (Bundy and McPherson,2000)。
西南喀斯特地区是我国的重要稻区,毗邻中南半岛,而多年的研究表明,越南、老挝、缅甸等地处中南半岛的国家正是稻飞虱滋生繁衍的巢穴(齐会会等,2011;沈慧梅等,2011;翟保平等,2011),所以西南喀斯特地区成为稻飞虱迁入我国的“第一站”,对该地区稻飞虱的发生规律、虫源地等的研究尤为必要。为了摸清喀斯特地貌最为广泛和典型的贵州省的灯下白背飞虱种群数量动态,本文对近4年来黔北、黔中、黔东和黔南4个稻区代表站点的白背飞虱灯下种群动态进行了调查分析。
1 材料与方法
1.1 灯诱地点
在贵州省内选择有代表性的稻区对白背飞虱进行灯光诱集,包括黔北稻区(道真仡佬族苗族自治县,简称道真)、黔中稻区(惠水县)、黔东稻区(锦屏县)、和黔南稻区(三都水族自治县,简称三都),各稻区代表地点详情见表1。
1.2 诱虫灯设置
以200 W 白炽灯为诱虫光源;将测报灯安装固定在高60 cm,宽110 cm的基台上,远离其他光源;每天黄昏时段开灯,次日清晨关灯,逐日取虫,并进行室内鉴别,记录诱到白背飞虱的雌雄成虫数量;记录每日天气及气温情况:采用中央气象台(http://www.nmc.gov.cn/)的每日天气数据;测报灯具体开关灯日期见表1。
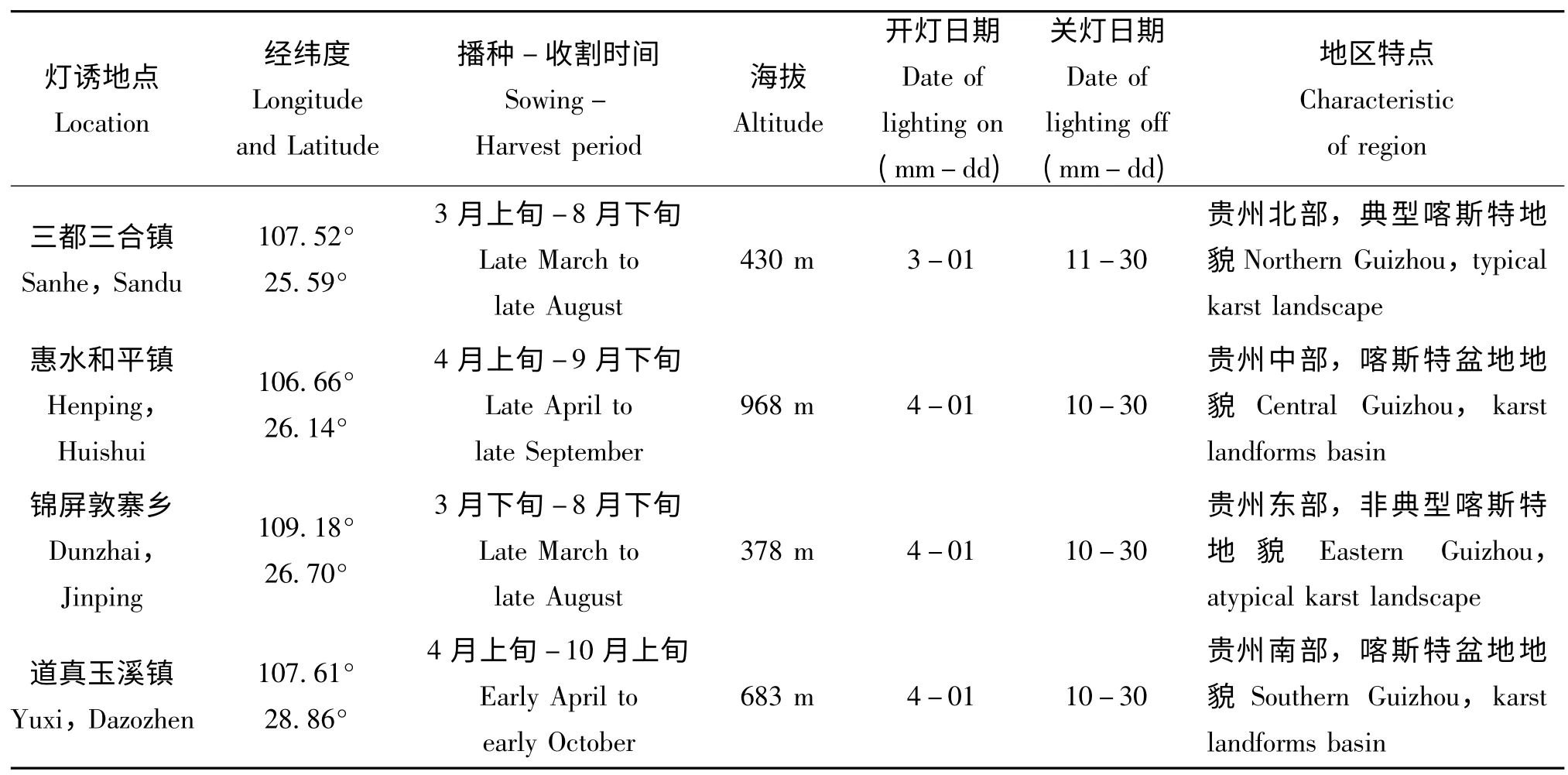
表1 灯诱地点基本信息Table 1 Basic information of light trapping location inGuizhou
1.2 白背飞虱灯下发生世代的划分
采用《贵州省稻飞虱监测技术规范》(贵州省质量技术监督局,2005) (下称《规范》)的世代划分标准,即4月20 日以前发生的划分为第1 代;4月21 日至5月20 日为第2 代;5月21 日至6月20 日为第3 代;6月21 日至7月20 日为第4 代;7月21 日至8月20 日为第5 代;8月21 日至9月20 日为第6 代;9月21 日至10月20 日第7 代;10月21 日以后划分为第8 代。
2 结果与分析
2.1 始见日
贵州地区少有白背飞虱的越冬场所,灯下始见即表明其外地虫源开始迁入,近4年的灯诱资料表明,三都稻区的灯下始见日多在4月中下旬,最早的年份(2010年)与最晚的年份(2011)的灯下始见日相差将近2个月;惠水稻区的灯下始见日与三都稻区相近,在4月中旬,最早年份与最晚年份的灯下始见日相差不大;锦屏稻区的灯下始见日较三都稻区和惠水稻区要晚,在4月下旬至5月中旬,最早年份与最晚年份的灯下始见日相差接近1个月;道真稻区的灯下白背飞虱多数在5月初始见,其中2009年最晚,在5月11 日(表2)。总的来看,贵州省三都稻区和惠水稻区的白背飞虱灯下始见日期较早,在4月中下旬,锦屏稻区和道真稻区的灯下始见日稍晚,在4月下旬至5月中旬。4个代表稻区中,始见日诱到的白背飞虱数量都较少,大都在个位数级别,最多的不过10 头。
2.2 始盛日
始盛日是指从该日起连续5 天以上均能诱获到白背飞虱且诱获的数量有逐日增加的趋势。从表2 可以看出,三都稻区4年间的灯下始盛日差异较大,最早的是2010年,为4月19 日,最晚的是2011年,为6月1 日;惠水稻区的灯下始盛日多出现在5月上中旬,各年份之间差别不大;锦屏稻区的灯下始盛日各年份之间差异也较大,从5月上旬至6月下旬均有出现;道真稻区的灯下始盛日的年度差异也较大,最早在5月上旬,最晚至7月初(表2)。对比始见日可以发现,在多数年份,始见日越早,始盛日也会相应提前。
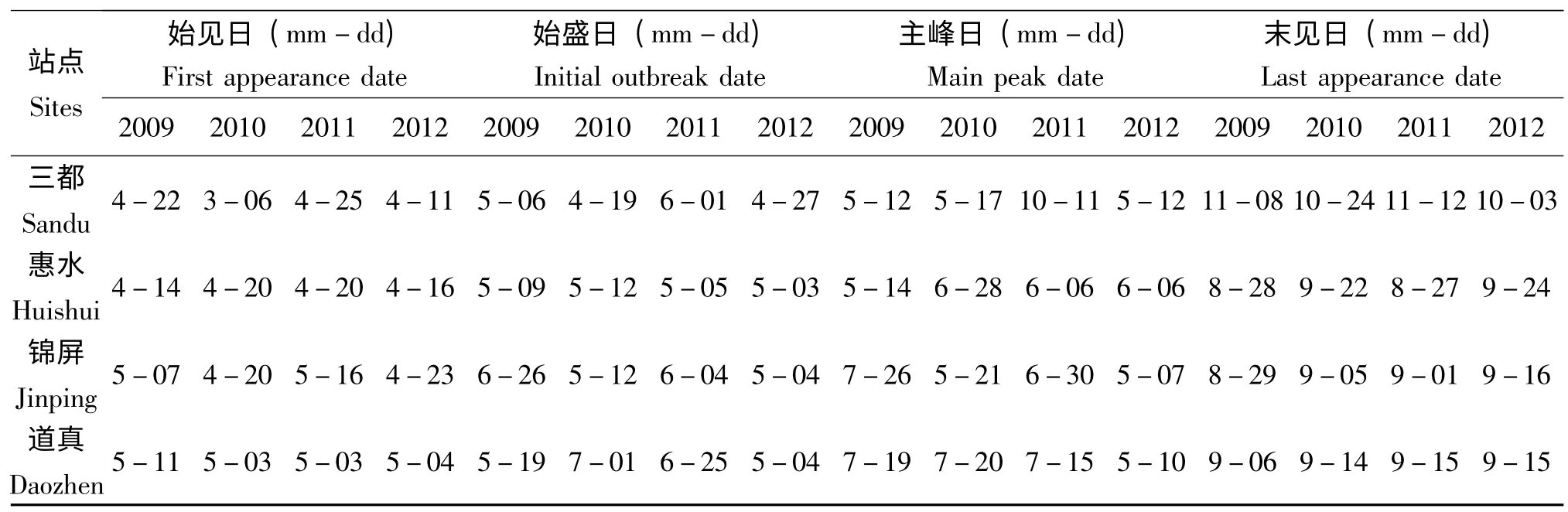
表2 2009-2012年灯下白背飞虱各发生期Table 2 Occurrence date of light trap catches of WBPH from 2009 to 2012 in Guizhou
2.3 高峰日
三都稻区灯诱高峰日多在5月中旬,2011年情况较为异常,在10月11 日达到高峰;惠水稻区较三都有所推迟,高峰日多在6月上旬出现;锦屏稻区的灯下高峰日各年份间差异较大,最早的在5月7 日,最晚的在7月26 日;道真稻区的灯下高峰日多在7月中旬出现(表2);从三都—惠水—锦屏—道真,白背飞虱的灯下高峰日依次向后推迟;各站点灯下主峰日与始见日、始盛日均呈显著正相关,相关系数均在0.9 以上(三都灯下主峰日与始见日、始盛日的相关系数为r=0.991**、r=0.991**,惠水灯下主峰日与始见日、始盛日的相关系数为r=1.000**、r=1.000**,锦屏灯下主峰日与始见日、始盛日的相关系数为r=0.999**、r=1.000**,道真灯下主峰日与始见日、始盛日的相关系数为r=0.999**、r=1.000**)。
2.4 始见日和始盛日至高峰日的间隔时间
统计近4年来4个稻区白背飞虱灯下始见日和始盛日至高峰日的时间间隔,发现三都稻区灯下白背飞虱从始见日到高峰日的时间为19-167 d,平均70.75 ±33.69 d,始盛日到高峰日的时间为5-130 d,平均43.75 ±29.07 d;惠水稻区从始见日到高峰日需29-68 d,平均50.75 ±8.16 d,始盛日到高峰日的时间为4-46 d,平均为28.25 ±8.82 d;锦屏稻区从始见日到高峰日需13-79 d,平均41.50 ±14.01 d,始盛日到高峰日的时间为2-25 d,平均为16.00 ±6.52 d;道真稻区从始见日到高峰日需5-68 d,平均55.50 ±16.75 d,始盛日到高峰日的时间为5-60 d,平均为25.50 ±11.93 d。
4个稻区白背飞虱灯下各发生期间的时间间隔并未表现出显著差异,但从近4年的数据可以看出不论是始见日至高峰日还是始盛日至高峰日均是三都稻区所需时间最长,锦屏稻区所需时间最短,惠水和道真稻区各发生期间的间隔天数较为接近,居于锦屏和三都稻区之间。
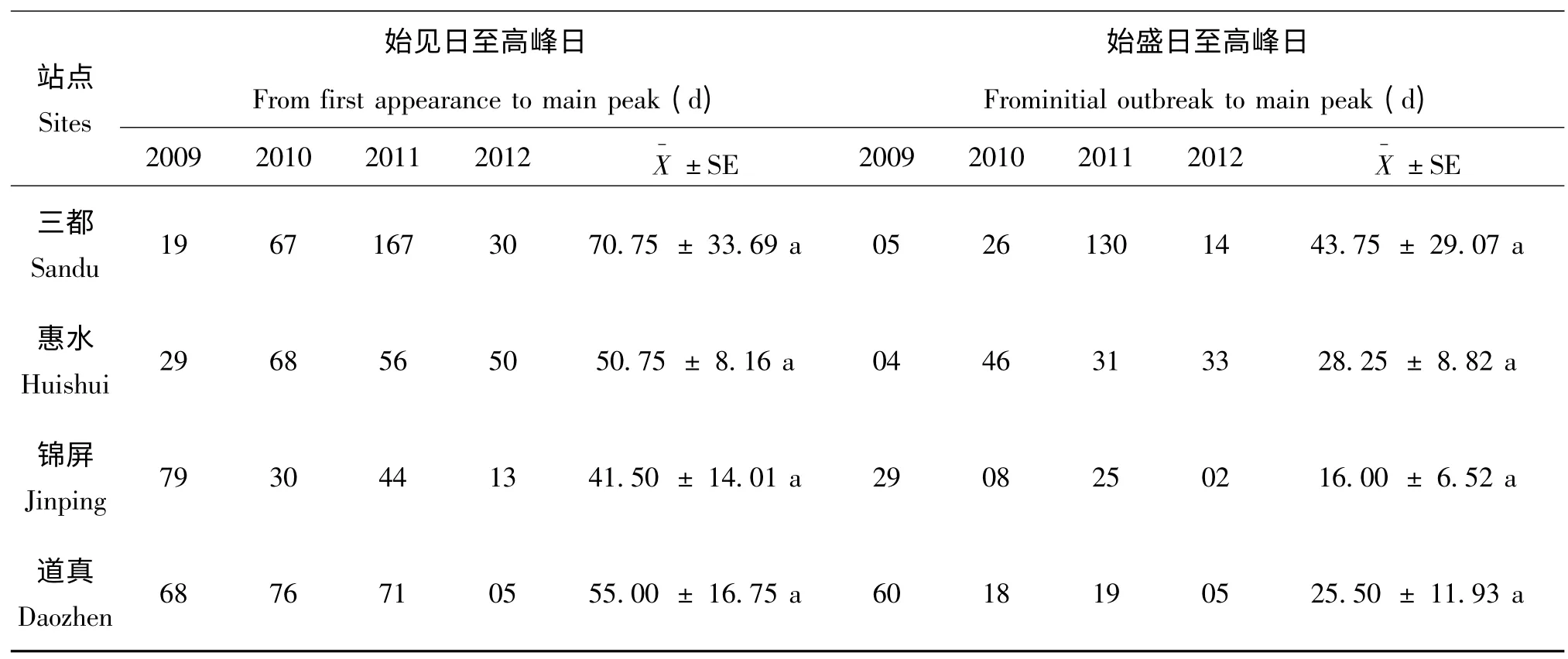
表3 2009-2011年灯下白背飞虱始见日和始盛日至高峰日的时间Table 3 Intervals fromfirst appearance to main peak and initial outbreak to main peak from 2009 to 2012 in Guizhou
2.5 末见日
4年之中,三都稻区的白背飞虱灯下末见日多出现在10月下旬至11月中上旬,最早的是在2012年,为10月3 日;惠水稻区则是在8月下旬和9月下旬末见,最早的是2011年的8月27 日,最晚的是2012年的9月24 日;锦屏稻区的灯下末见日出现较为集中,多在9月中上旬;道真稻区的灯下末见日出现也较为集中,多在9月中旬(表2)。比较4个稻区可以发现,三都稻区的灯下末见日出现最晚,惠水稻区次之,锦屏稻区和道真稻区的灯下末见日出现最早。
2.6 灯下发生量
4年间各稻区年度诱虫量差异较大,从图1 可以看出,白背飞虱的灯下最高诱获量出现在2012年三都,在25 万头以上,其次是2010年三都,诱获量在10 万头以上,2012年惠水的诱获量在5 万头以上,其余各年份和代表稻区的灯下诱获量都在5 万头以下,最少的是2009年道真,只有5083头。从4年间的平均数据来看,三都的灯下虫量最多(100348 头),其次是惠水(31117 头),道真(15670 头)和锦屏(11915 头)。
以世代来划分,白背飞虱灯下诱获量主要集中在第2-6 代:三都稻区灯下第2 代虫量最多,第3 至5 代依次减少,到第6 代又有较大幅度的上升;惠水稻区的灯下最高虫量同样出现在第2 代,此后依次减少,至第6 代减少至100 头;锦屏稻区第2 至5 代灯下虫量基本趋于平稳,第5 代诱获量最多,至第6 代锐减;道真稻区的情况较为复杂,有两次虫量高峰,分别出现在第2 代和第4 代,而第3 代的虫量却跌至谷底(图2)。对不同稻区4年间灯下虫量的方差分析表明,三都在第3 代灯下虫量上与道真存在显著差异,在第6 代虫量上与惠水、锦屏也存在显著差异,其他各代虫量间不同稻区并无显著差异(表4)。
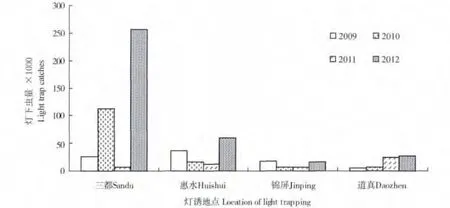
图1 灯诱白背飞虱的年度间变化Fig.1 The interannual fluctuation of the light-trap catches of WBPH from 2009 to 2012 in Guizhou
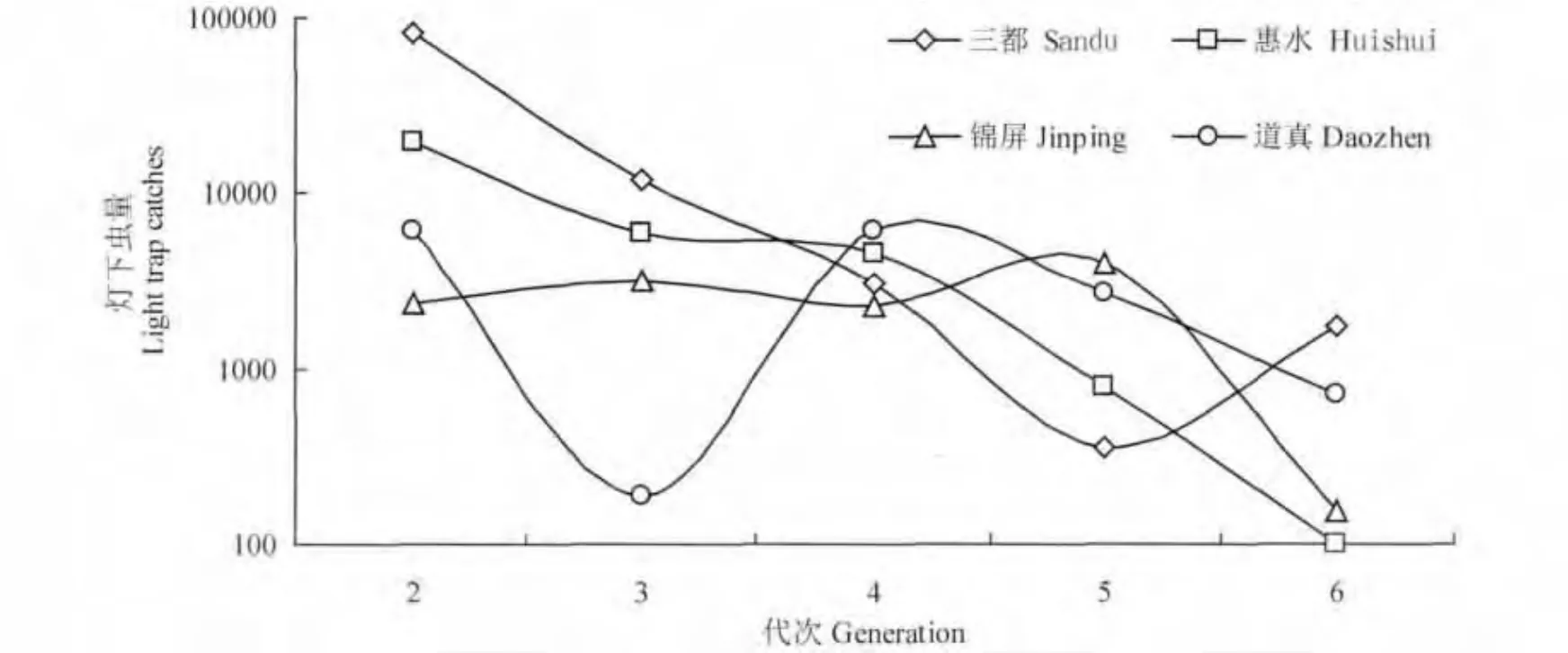
图2 2009-2012年灯下各代白背飞虱平均诱获量Fig.2 The average value of the light-trap catches of 5 generations of WBPH from 2009 to 2012 in Guizhou
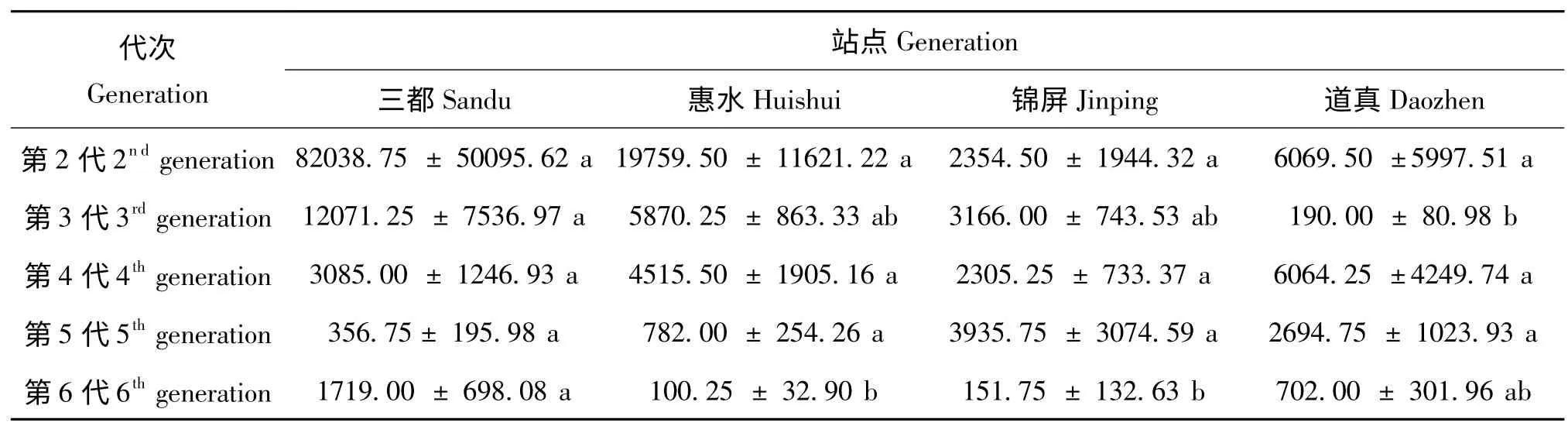
表4 不同稻区各世代灯下虫量的差异性Table 4 The differences ofthe light-trap catches of WBPH of 5 generations in different rice regions in Guizhou
2.7 灯下发生量的性别比例
性别构成是衡量种群的重要参数,如表5 所示,4年间各稻区中,白背飞虱灯下种群的雌雄性比均大于1,表明白背飞虱灯下种群中雌虫数量较多。不同稻区的雌雄性比未表现出统计学上的显著差异,但是就现有数据来看,道真稻区和锦屏稻区的灯下种群雌雄性比高于三都稻区和惠水稻区。

表5 2009-2012年贵州各稻区灯下白背飞虱雌雄性比Table 5 Sex ratio of light-trap population of WBPH from 2009 to 2012 in Guizhou
3 结论与讨论
白背飞虱作为一种长距离迁飞型害虫,在贵州本地很少有越冬场所,灯下始见日和高峰日可以在一定程度上反应其初迁入期与高峰迁入期,从近4年的灯下数据可以看出:在4月中下旬白背飞虱开始迁入南部和中部稻区,东部和北部稻区稍微延后,在4月下旬至5月上旬迁入;高峰迁入期在各稻区间也有一定差异,南部稻区在5月中旬达到迁入高峰,中部稻区高峰日在多在6月上旬出现;东部稻区的灯下高峰日在5月上旬至6月下旬,北部稻区的灯下高峰日多在7月中旬出现,这与前人的(贵州省稻飞虱联合测报组,1984;金星等,1998)的研究结果基本一致,但2011年三都的情况较为特殊,白背飞虱灯下始见日、始盛日与高峰日均呈现异常,通过分析三都的逐日气象数据,发现从3月22 日至3月26 日,连续5 d的日均温都在10℃以下(日均温分别为6℃、9.5℃、8℃、9.5℃、8.5℃)并普遍伴随着降雨现象,达到了中度倒春寒的标准 (许炳南,1999),而3月下旬西南气流盛行,正是白背飞虱从中南半岛随气流迁入我国西南边境的时期,恰逢这次倒春寒,使白背飞虱的前期迁入严重受挫,从而导致了灯下始见日异于常年;同时因为倒春寒的影响,三都水稻的育苗和插秧进度有所延缓,这也增长了白背飞虱食料的“真空期”,导致了灯下始盛日的推迟;由于倒春寒形成的低温屏障和食料“真空期”的延长,在前期并没有出现大的白背飞虱迁入峰,10月份回迁出境的越冬虫源反而形成了白背飞虱的灯下主峰。
从灯下发生量来看,三都最多,惠水次之,锦屏和道真较少,与贵州稻飞虱的发生程度区划相符(贵州省稻飞虱联合测报组,1984),各站点灯下发生量的差异可能与各稻区的海拔、水热、地形有关,南部稻区的代表站点三都地处苗岭山脉南侧的河谷地带,北高南低、水热充足,十分有利于外来稻飞虱的迁入与降落,故发生量最大。在所掌握的4年数据中,4 站点灯下白背飞虱的雌雄性比均大于1,其中,锦屏稻区和道真稻区白背飞虱的灯下种群雌雄性比的年度间差异大于三都稻区和惠水稻区,这可能是由于锦屏稻区与道真稻区白背飞虱灯下种群数量较少而导致的统计学误差较大所造成的。
灯下白背飞虱雌虫数量多于雄虫,在一定程度上表明雌虫的迁入多于雄虫。研究表明:白背飞虱雌虫的取食量要显著高于雄虫,且在羽化后2-4 d 达到取食高峰(黄次伟和冯炳灿,1993),而这段时间正是白背飞虱开始迁飞行为的时间,即成虫的幼嫩阶段(陈若箎等,1979;陈宇等,2012);在这段时期内稻飞虱的飞行能力最强(陈若箎等,1984),且雌虫的飞行时间要长于雄虫(汪远昆和翟保平,2004);在这段时间内稻飞虱大量取食的原因可能就是为迁飞行为储备能量,正是由于雌虫取食量大、体内能量物质存储多,所飞行时间长于雄虫,而且雌虫是大多数基因流动和散布的承担者,与雄虫相比更喜欢扩散,具有更强的迁飞潜力和迁飞欲望 (Johnson,1963;James,1975),这可能是灯下雌虫数量多于雄虫的原因。
References)
Bundy CS,McPherson RM.Dynamics and seasonal abundance of stink bugs (Heteroptera:Pentatomidae)in a cotton-soybean ecosystem[J].Journal of Economic Entomology,2000,93 (3):697-706.
Chen RC,Cheng XN,Yang LM,et al.The ovarial development of brown planthopper (Nilaparvata lugens Stål)and it's relation to migration [J].Acta Entomologica Sinica,1979,22 (3):280-288.[陈若箎,程瑕年,杨联民,等.褐飞虱卵巢发育及其与迁飞的关系[J].昆虫学报,1979,22 (3):280-288]
Chen RC,Wu JR,Zhu SD,et al.Fight capacity of the brown planthopper Nilaparvata lugens Stål [J].Acta Entomologica Sinica,1984,27 (2):121-127.[陈若箎,吴家荣,祝树德,等.1984.褐飞虱的飞翔能力[J].昆虫学报,1984,27 (2):121-127]
Chen Y,Fu Q,Lai FX,et al.The effects of rice growth stages on the ovarian development and take-off of Nilaparvata lugens and Sogatella furcifera [J].Acta Ecologica Sinica,2012,32 (5):1546-1552.[陈宇,傅强,赖凤香,等.水稻生育期对褐飞虱和白背飞虱卵巢发育及起飞行为的影响[J].生态学报,2012,32 (5):1546-1552]
Cheng JA.Rice planthopper problems and relevant causes in China.In:Heong KL,Hardy B,eds.Planthoppers:new threats to the sustainability of intensive rice production systems in Asia [C].Los Baños:International Rice Research Institute,2009,157-178.
Guizhou Province Planthopper Forecasting Group.Guizhou planthopper research (1981-1983)[J].Guizhou Agricultural Sciences,1984,(5):24-28.[贵州省稻飞虱联合测报组,1984.贵州稻飞虱研究 (1981-1983年)[J].贵州农业科学,1984,(5):24-28]
Hu GW,Xie MX,Wang YC.Asuggestion for delimitation of the incidence of areas of white-backed planthoppers in China [J].Acta Entomologica Sinica,1988,31 (1):42-48.[胡国文,谢明霞,汪毓才.对我国白背飞虱的区划意见[J].昆虫学报,1988,31 (1):42-48]
Huang CW,Feng BC.Quantitative aspects of feeding activity in thewhite-backed planthoppers Sogatella furcifera (Horváth)and the brown planthopper Nilaparvata lugens (Stål)[J].Acta Entomologica Sinica,1993,36 (2):251-255.[黄次伟,冯炳灿.水稻白背飞虱,褐飞虱取食动态研究[J].昆虫学报,1993,36 (2):251-255]
James AS.Flight patterns among eleven species of diurnal Lepidoptera[J].Ecology,1975,56 (6):1367-1377
Jin X,Wang DQ,Jin DC,et al.A review on the occurrence dramatics and management of rice planthopper Sogatella furcifera (Horváth)and Nilaparvata lugens (Stål) in Guizhou [J].Journal of Mountain Agriculture and Biology,1998,17 (4):208-214.[金星,王德其,金道超,等.贵州省稻飞虱发生危害规律与防治对策[J].山地农业生物学报,1998,17 (4):208-214]
Johnson CG.Physiological factors in insect migration by flight [J].Nature,1963,198 (4879):423-427
National Research Group of white-backed planthopper.Studies on the migration of white back planthoppers (Sogatella furcifera,Horváth)[J].Scientia Agriculture Sinica,1981,(5):25-31.[全国白背飞虱科研协作组.白背飞虱迁飞规律的初步研究[J].中国农业科学,1981,(5):25-31]
Qi HH,Zhang YH,Jiang CX,et al.Analysis of the source areas of the early immigration of white-backed planthopper in northeast Guangxi of China [J].Scientia Agriculture Sinica,2011,44(16):3333-3342.[齐会会,张云慧,蒋春先,等.广西东北部稻区白背飞虱早期迁入虫源分析[J].中国农业科学,2011,44 (16):3333-3342]
Qin HG,Ye ZX,Shu C,et al.Sogatella furcifera Populations Governance Theory and Practice [M].Nanchang:Jiangxi Science and Technology Press,2003,1-362.[秦厚国,叶正蘘,舒畅,等.白背飞虱种群治理理论与实践[M].南昌:江西科学技术出版社,2003,1-362]
Scientific Research Coordinated Group for Planthoppers ofSichuan,Yunan and Guizhou Provinces.Migration and occurrence of whitebacked planthoppers and brown planthoppers in Southwest China[J].Acta Phytophylacica Sinica,1982,9 (3):179-186.[四川、贵州、云南三省稻飞虱科研协作组.我国西南稻区白背飞虱,褐稻虱的迁飞和发生特点[J].植物保护学报,1982,9(3):179-186]
Shen HM,Lv JP,Zhou JY,et al.Source areas and landing mechanism of early immigration of white-backed planthoppers Sogatella furcifera (Horváth)in Yunnan,2009 [J].Acta Ecologica Sinica,2011,31 (15):4350-4364.[沈慧梅,吕建平,周金玉,等.2009年云南省白背飞虱早期迁入种群的虫源地范围与降落机制[J].生态学报,2011,31 (15):4350-4364]
Wang YK,Zhai BP.Re-migration capacity of the white-backed planthopper,Sogatella furcifera (Horváth) [J].Acta Entomologica Sinica,2004,47 (4):467-473.[汪远昆,翟保平.白背飞虱的再迁飞能力[J].昆虫学报,2004,47 (4):467-473]
Xu BN.Criteria for the classification of meteorological disasters in Guizhou [J].Journal of Guizhou Meteorology,1999,23 (3):42-47.[许炳南.贵州气象灾害的划分标准[J].贵州气象,1999,23 (3):42-47]
Zhai BP.Rice planthoppers:A China problem under the international perspectives [J].Chinese Journal of Applied Entomology,2011,48 (5):1184-1193.[翟保平.稻飞虱:国际视野下的中国问题[J].应用昆虫学报,2011,48 (5):1184-1193]
