血清对U937细胞分化成熟及细胞周期的影响
2014-11-24曹林林杜珍武杨麒巍侯治富
曹林林,杜珍武,杨麒巍,侯治富*
(1.淄博市中心医院 检验科,山东 淄博255036;2.吉林大学中日联谊医院 中心研究室,吉林 长春130033)
肿瘤细胞的分化程度是癌症诊断和治疗中一个重要的参考的数据,在研究肿瘤细胞分化过程中,发现细胞分化与细胞停滞联系密切[1,2],每一次细胞分化都伴有细胞周期停滞[3]。
本研究利用乙酸肉豆蔻佛波醇(PMA)[4]体外诱导单核白血病细胞系——U937细胞分化为研究对象,通过对在有无血清培养条件下细胞形态及周期的变化,探讨调控细胞分化成熟的机制,并为白血病的研究性治疗提供依据。
1 材料与方法
1.1 实验材料
U937细胞株来自吉林大学中日联谊医院中心研究室,胎牛血清(FCS),IMDM培养基,青链霉素均购于Hyclone公司,细胞周期检测试剂盒购于碧云天生物技术公司,PMA购于美国Sigma公司。
1.2 U937细胞培养
将U937细胞株复苏后接种于含15%胎牛血清和100ml/L青链霉素的IMDM培养基中。置于含有5%CO2,37℃培养箱内进行培养。
1.3 血清为培养调节因素情况下PMA诱导U937细胞分化
根据在U937细胞培养过程中是否添加血清与PMA,将细胞分成4组:①FCS组:U937细胞置于含有10%FCS的IMDM培养基内培养;②FCS+PMA组:U937细胞置于含有10%FCS与10ng/ml PMA的IMDM培养基内培养;③NOFCS组:U937细胞置于无FCS的IMDM培养基内培养;④NOFCS+PMA组:U937细胞置于无FCS但含有10 ng/ml PMA的IMDM培养基内培养。培养24h后应用倒置显微镜进行形态学观察、贴壁率计算与细胞周期检测。
1.4 血清恢复实验
分别取NOFCS和NOFCS+PMA组培养24 h U937细胞,以1×106/瓶将其接种于含血清培养基的培养瓶内进行培养,培养24h后应用倒置显微镜进行形态学观察、贴壁率计算以及细胞周期检测。
1.5 分化后细胞的无血清培养
将FCS+PMA组细胞培养24h后,分别置于含有血清培养基与无血清培养基内,培养24h后应用倒置显微镜进行形态学观察、贴壁率计算与细胞周期检测。
1.6 细胞贴壁率测定
分别收集各组悬浮细胞与贴壁细胞,应用细胞计数仪(TC10,Bio-Rad)进行细胞计数,按照下面公式进行贴壁率计算:细胞贴壁率%=贴壁细胞数/(悬浮细胞数+贴壁细胞数)。
1.7 流式细胞术检测细胞周期
取各组细胞,用PBS洗涤1次。经75%的冷酒精固定18h,1 000r/min离心5min,去除酒精,用PBS涤洗3次,加入PBS 0.5ml。用350目纱网过滤后加入RNA酶100μl振荡,37℃水浴30min。加入碘化丙啶(PI)400μl振荡,4℃30min后上流式细胞仪进行细胞周期分析。
1.8 统计分析
采用SPSS17.0软件进行统计学处理,细胞贴壁率、细胞周期数据均以Mean±SD表示,组间比较采用单因素方差分析。
2 结果
2.1 血清对U937细胞分化过程的细胞形态影响
在FCS组细胞在血清培养基内为圆形、悬浮、分散生长(图1A),FCS+PMA组细胞为贴壁生长,细胞相互粘附成团,细胞颗粒度增多(图1B);NOFCS组细胞悬浮、分散生长(图1C);NOFCS+PMA组细胞部分聚集生长,大部分细胞仍悬浮生长(图1D),说明血清在PMA诱导的U937细胞分化过程对于细胞的形态具有一定影响。
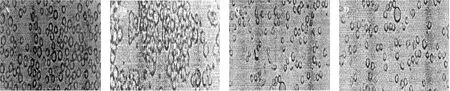
图1 U937细胞在不同培养条件下形态学变化(200×)
2.2 血清对U937细胞分化过程的细胞贴壁率影响
FCS+PMA组细胞贴壁率明显高于其他3组,而NOFCS+PMA组细胞的贴壁率与FCS组,NOFCS组无明显差异,说明血清在PMA诱导的U937细胞分化过程对于细胞的贴壁具有一定影响。结果见表1。
2.3 血清对U937细胞分化过程的细胞周期影响
与FCS组(图2A)相比,FCS+PMA(图2B)、NOFCS(图2C)和NOFCS+PMA组(图2D)细胞周期的G1细胞明显增多,而S期细胞明显减少,相比有显著差异,说明血清可以影响U937细胞的细胞周期改变,结果见图2,表2。

表1 各组细胞的贴壁率

图2 各组细胞的细胞周期直方图

表2 各组细胞的细胞周期分布
2.4 血清恢复后细胞形态学观察与细胞周期结果
NOFCS组与NOFCS+PMA组细胞置于含有15%血清培养基,培养24h后,NOFCS组细胞悬浮生长(图3A),细胞周期结果分析显示与血清恢复前NOFCS组相比,G1期细胞数目明显减少,S期细胞明显增多(图4A,表3);NOFCS+PMA组细胞贴壁生长,聚集成团(图3B),细胞周期结果分析显示与血清恢复前NOFCS+PMA组相比,细胞周期无明显改变(图4B,表3)。
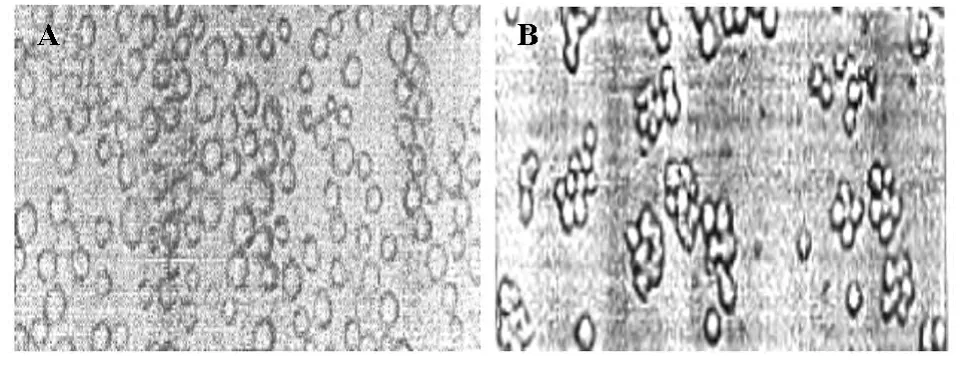
图3 血清恢复后NOFCS组与NOFCS+PMA组细胞形态学观察结果
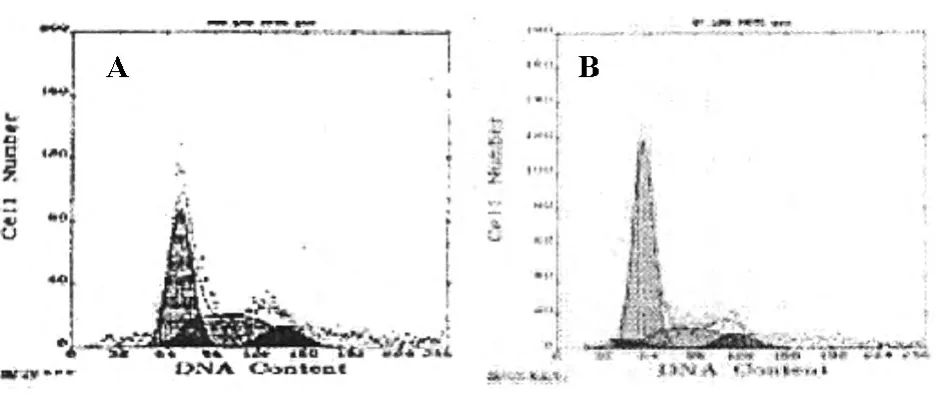
图4 血清恢复后NOFCS组与NOFCS+PMA组细胞周期观察结果

表3 血清作用前后U937细胞周期分布的变化
3 讨论
U937细胞是一种前单核细胞株,其特点为经一定刺激后可向单核巨噬细胞方向分化,目前,U937细胞已经成为体外研究细胞分化机制常规使用的细胞模型之一。乙酸肉豆蔻佛波醇(Phorbol-12-myristate 13-acetata,PMA),是二脂酰甘油(DG)的类似物,可以进入细胞内,代替DG的活性激活蛋白酶(PKC)的活性[5],引起细胞内一系列的生物学效应。在体外,PMA可以诱导多种肿瘤细胞系的分化[6,7],是一种实用良好、性能稳定的诱导分化剂。在细胞培养过程中,血清作为独立的影响因素,在肿瘤细胞体外生长过程中起着重要作用,主要由于血清含有细胞增殖与生存的必需营养素如某些生长因子。在无血清情况下,细胞停止增殖,细胞周期停滞于G1期,而这些细胞是否分化,各研究结论是不同的[8-11],主要是由于研究对象及所用诱导剂不同的缘故。本研究利用PMA诱导分化U937细胞,观察并研究以血清作为调节因素情况下,U937细胞分化成熟及细胞周期的变化。
研究结果表明,在有血清培养条件下,经10ng/ml PMA作用24小时后,U937细胞形态学发生巨大变化,大量细胞由悬浮变为贴壁生长,细胞之间由分散状态变为彼此粘附状态,细胞形态由圆形变为多角形或椭圆形,细胞周期表现为G1期细胞明显增多,S期细胞明显减少,并出现细胞凋亡现象。通过细胞形态、贴壁率及细胞周期的改变,可见血清促进了细胞分化成熟,也说明细胞诱导分化成功。另外细胞周期测定结果显示,未贴壁细胞凋亡率明显高于贴壁细胞,可能是由于细胞贴壁生长后有利于细胞的存活。本实验分别测定了在正常血清(FCS)、血清加PMA(FCS+PMA)、无血清(NOFCS)、无血清+PMA(NOFCS+PMA)4种情况下细胞周期的变化,FCS组与NOFCS组相比细胞周期停滞于G1期。血清恢复实验结果表明,NOFCS+PMA组细胞周期仍停滞于G1期,而NOFCS组细胞周期开始恢复。出现这种现象的可能原因有两个:一是PMA作用,因为在整个实验过程中PMA在细胞体内不易代谢,可长期发挥作用。所以在血清恢复后可继续发挥作用;二是据文献报道在无血清情况下,细胞CyclinD表达会下降,但在血清恢复后,可能重新恢复表达,促进细胞周期的进程[12]。
细胞周期的进程主要由两组相互消长的蛋白控制即细胞周期蛋白及细胞周期依赖性激酶、细胞周期蛋白依赖性激酶抑制蛋白[13]。对于细胞周期G1期的停滞可以通过下调细胞周期蛋白酶的活性或上调细胞周期蛋白依赖性激酶抑制蛋白活性来进行调控。本研究同时证实了PMA对U937细胞的分化发挥重要调控作用,有利于寻找新型抗白血病药物,但具体的调控机制还在进一步探讨之中。
[1]Pardee Ab.G1event and regulation of cell proliferation[J].Science,1989,246:603.
[2]Hayden A,Johnson PWM,Packham G,et al.S-adenosylhomocys-teine hydrolase inhibition by 3-deazaneplanocin A analogues induces anti-cancer effects in breast cancer cell lines and synergy with both histone deacetylase and HER2inhibition[J].Breast cancer research and treatment,2011,127(1):109.
[3]Saito Y,Miyahara R,Gopalan B,et al.Selective induction of cell cycle arrest and apoptosis in human prostate cancer cells through adenoviral transfer of the melanoma differentiation-associated-7(mda-7)/interleukin-24 (IL-24)gene[J].Cancer gene therapy,2005,12(3):238.
[4]Kikuchi H,Yuan B,Nishimura Y,et al.Cytotoxicity of Vitex agnus-castus fruit extract and its major component,casticin,correlates with differentiation status in leukemia cell lines[J].International Journal of Oncology,2013,43(6):1976.
[5]Blumberg.Protein Kinase C as the Receptor for the Phorbol Ester Tumor Promoters:Sixth Rhoads Memorial Award Lecture1[J].Cancer Research,1988,48:1.
[6]Zhou L,Shen L,Hu L,et al.Retinoid X receptor agonists inhibit phorbol-12-myristate-13-acetate(PMA)-induced differentiation of monocytic THP-1cells into macrophages[J].Molecular and cellular biochemistry,2010,335(1-2):283.
[7]Jalagadugula G,Dhanasekaran D N,Rao A K.Phorbol 12-Myristate 13-Acetate (PMA)Responsive Sequence in Gαq Promoter During Megakaryocytic Differentiation:Regulation by EGR-1and MAP Kinase Pathway[J].Thrombosis and haemostasis,2008,100(5):821.
[8]Huo XF,Yu J,Peng H,et al.Differential expression changes in K562cells during the hemin-induced erythroid differentiation and the phorbol myristate acetate (PMA)-induced megakaryocytic differentiation[J].Molecular and cellular biochemistry,2006,292(1-2):155.
[9]Gottipamula S,Muttigi MS,Kolkundkar U,et al.Serum-free media for the production of human mesenchymal stromal cells:a review[J].Cell Proliferation,2013,doi:10.1111/cpr.12063.
[10]Haddad H,Papoutsakis ET.Low oxygen tension and autologous plasma enhance T-cell proliferation and CD49dexpression density in serum-free media[J].Cytotherapy,2001,3(6):435.
[11]Xin-xin S,Duncan NA,Lin LIN,et al.Serum-free media for articular chondrocytes in vitro expansion[J].Chinese medical journal,2013,126(13):2523.
[12]Resnitzky D,Reed SI.Different roles for cyclins D1and E in regulation of the G1-to-S transition[J].Molecular Cell.1995,15(7):3463.
[13]Lim S,Kaldis P.Cdks,cyclins and CKIs:roles beyond cell cycle regulation[J].Development,2013,140(15):3079.
