亚洲玉米螟生物学特性的研究
2014-11-06杨慧中涂小云夏勤雯薛芳森
杨慧中,涂小云,夏勤雯,薛芳森*
(1.江西农业大学 昆虫研究所,江西 南昌 330045;2.江西师范大学 生命科学院,江西 南昌 330022)
亚洲玉米螟(Ostrinia furnacalis(Guenée)),属鳞翅目(Lepidoptera)草螟科(Crambidae),俗称“钻心虫”,是我国最具有经济重要性的害虫之一,广泛分布于中国从南至北的玉米产区。该虫以幼虫蛀食茎秆、穗柄、穗轴等,严重影响玉米产区的玉米产量和质量。在我国主产区北方,一般年份春玉米受玉米螟为害而减产10%左右,大年发生可使玉米减30%以上,造成巨大损失[1]。由于玉米螟在北方玉米产区的严重为害,我国对亚洲玉米螟的北方种群生物学特性做了较详细的研究[2-7]。在我国南方,由于玉米的种植面积较小,玉米仅作为零食,因此对玉米螟的生物学特性缺乏系统研究[8]。然而,随着南方玉米种植面积的不断扩大,亚洲玉米螟对玉米的为害明显加重,成为重要的防治对象。因此,本课题组于2011—2013年对亚洲玉米螟南昌种群的年生活史、温度对其生长发育和繁殖的影响及越冬生物学进行了较系统的研究,试图为该虫发生动态的预测预报和防治策略的制定提供基础资料。
1 材料与方法
1.1 供试虫源
用于实验的亚洲玉米螟种群采自江西南昌(28°68'N,115°89'E)玉米地,用扫虫网在玉米杂草丛中网捕成虫。将网捕的雌雄成虫放入带有10%糖水棉花球的保鲜袋中供其产卵,每天收集卵块。幼虫孵化后装有人工饲料的养虫盒(直径12 cm,高15 cm)中饲养,人工饲料配方参照乔利等[9]。幼虫饲养到4龄后转入24孔板中单头饲养,详细观察其化蛹和羽化时间。所有试验均在LRH-250-GS型光照培养箱中进行,光强为500~700 lx,温度变动幅度为±1℃,湿度维持在70%~80%。
1.2 羽化性
将越冬代最早批和最迟批羽化的成虫分别在室外进行系统连续繁殖,即以后各代均采用最早和最迟羽化的成虫进行繁殖,直至幼虫全部进入越冬,从而得出1年可能发生的世代数。
1.3 各虫态发育历期
将卵、幼虫和蛹置于20,22,25,28,31℃,光周期为16L∶8D观察其发育历期。每天记载卵的孵化、幼虫化蛹和成虫羽化时间。获得的结果采用最小二乘方公式计算卵、幼虫和蛹的发育起点温度(lower development threshold,LDT)和有效积温(lum of effective temperatures,SET)。
1.4 成虫繁殖特性
将当日羽化的第3代成虫一雌一雄配对放入带有10%糖水棉花球的保鲜袋中供其产卵,保鲜袋置于25℃、RH75%、光周期16L∶8D的人工气候箱中。逐日更换保鲜袋和棉球,将保鲜袋上的卵于解剖镜下计数,记录开始产卵日期、终止产卵日期和雌雄死亡日期,观察20对以上。
1.5 越冬生物学
从8月下旬至9月中旬止,每隔2~3 d接一批幼虫放置在室外饲养,记录其化蛹数。至11月初仍未见化蛹的幼虫判断为滞育。将上一年越冬代幼虫于4月初逐头转移到24孔板进行观察,单个记录其化蛹日期和成虫羽化日期。
1.6 数据统计与分析
实验数据采用SPSS13.0数据处理系统进行分析,用one-way ANOVA进行方差分析,结果用平均值±标准差(mean±SD)表示。
2 结果与分析
2.1 年生活史
从年生活史(表1)可以看出,越冬个体次年4月下旬开始化蛹,至6月初结束。越冬代成虫于5月上旬开始羽化,至6月中旬结束。越冬代羽化的成虫5月上旬末开始产卵,至6月下旬初结束。第1代幼虫5月中初旬开始孵化,至7月初结束。越冬代最早羽化的少量个体在室外连续繁殖1年可发生5代,而越冬代最迟羽化的少量个体1年只繁殖3代,大多数个体1年发生4代。因此,亚洲玉米螟在南昌1年可发生3~5代,以4代为主。从表中还可看出,亚洲玉米螟存在明显世代重叠现象,6月上旬可见越冬代和第一代成虫并存,7月份可见第2代和第3代成虫并存,8月底可见第3代和第4代成虫并存。
2.2 主要生活习性
成虫白天和晚上均可羽化,大部分在晚上羽化,羽化后1~2 d即可交尾。成虫交尾在晚上进行。据室内20对成虫繁殖情况的观察,在25℃、光周期16L∶8D下,产卵前期为(3.23±0.87)d,产卵历期为(10.45±1.95)d,产卵量为(500.9±66.77)粒,雌虫寿命为(13.68±2.06)d,雄虫寿命为(12.50±2.92)d。据田间调查,成虫白天一般潜藏于玉米地旁茂密的杂草间,于日落后至午夜前活动,飞翔力强,有一定的趋光性。成虫常选择低于50 cm、生长浓绿的玉米植株上产卵,一般卵产于玉米叶背靠近叶脉处。
初龄幼虫孵化后先群集取食卵壳,经1 h左右开始爬行。行动敏捷、活泼、迅速,受惊可吐丝下垂,移到其他部位或植株。幼虫有趋糖、趋湿、背光性,4龄前表现为潜藏,喜藏于含糖高、湿、隐蔽之处,如心叶丛、雄穗包、雌穗顶端花丝基部、腋芽等处。幼虫危害玉米心叶时,被害叶片长出后,显示出“排孔”。4龄后为钻蛀,幼虫蛀入玉米茎秆内形成孔道,茎秆外留下一个小孔。幼虫老熟后吐一层薄丝作茧,于被害部位化蛹。

表1 亚洲玉米螟在南昌的年生活史Tab.1 Life history of Ostrinia furnacalis in Nanchang
2.3 各虫态发育历期
温度对亚洲玉米螟卵、幼虫和蛹的发育历期有显著影响(P>0.05),随着温度升高,卵和蛹的发育历期均逐渐缩短;有趣的是,在温度20~28℃,幼虫期随着温度升高逐渐缩短,但当温度上升到31℃时,幼虫的发育历期比25℃和28℃稍有延长。在20℃和22℃较低的温度下,雄幼虫的发育历期长于雌幼虫,而在较高温度25~31℃,雄幼虫的发育历期短于雌幼虫(表2)。

表2 不同温度下亚洲玉米螟的发育历期Tab.2 Developmental duration of Ostrinia furnacalis at different temperatures
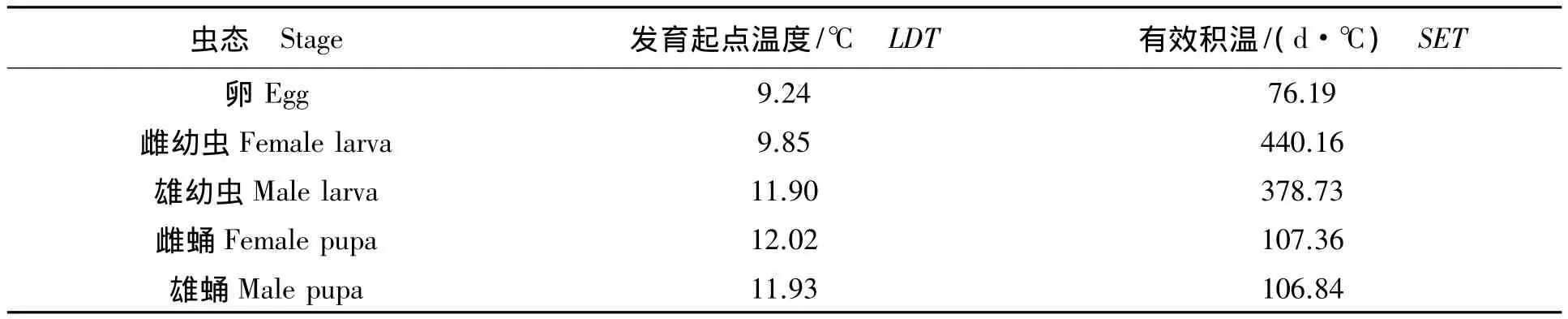
表3 亚洲玉米螟的发育起点温度和有效积温Tab.3 The lower development threshold and sum of effective temperatures of Ostrinia furnacalis

图1 亚洲玉米螟自然条件下幼虫滞育的发生(每观察点样本数为54~120头)Fig.1 Incidence of larval diapause in Ostrinia furnacalis under natural conditions(n=54~120 for each point)
2.4 亚洲玉米螟的发育起点温度和有效积温
从表3可以看出,各虫态的发育起点温度分别为卵9.24 ℃、雌幼虫9.85 ℃、雄幼虫 11.90℃、雌 蛹 12.02℃、雄 蛹11.93 ℃,有效积温分别为 76.19,440.16,378.73,107.36,106.84日·度。各虫态中卵的发育起点温度最低,为9.24℃;雌蛹的发育起点温度最高,为12.02℃。
2.5 越冬生物学
图1表明,1987年、2011年和2012年自然条件下,8月25日开始孵化幼虫有部分进入滞育,8月28日至8月29日滞育率达到50%,9月3日及以后孵化的幼虫全部进入滞育。从图2可以看出,2012年亚洲玉米螟越冬幼虫从4月22日开始化蛹,5月9日化蛹率达到50%,5月24日化蛹终止。2013年越冬幼虫从4月17日开始化蛹,5月2日化蛹率达到50%,5月22日化蛹终止。2012年亚洲玉米螟江西种群越冬幼虫从5月4日开始羽化,5月16日羽化率达到50%,6月5日羽化结束,越冬羽化持续时间达32 d。2013年越冬幼虫从4月27日开始羽化,5月16日化蛹率达到50%,6月2日羽化结束,越冬羽化持续时间达37 d。根据室外572个越冬个体单头观察,越冬个体的幼虫期为220~268 d,平均240.5 d。
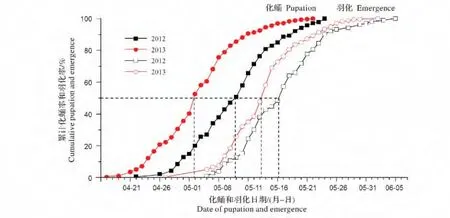
图2 亚洲玉米螟越冬代幼虫在自然条件下的化蛹和成虫羽化时间(2012年观察化蛹和羽化总数为140头,2013年观察化蛹和羽化总数为572头)Fig.2 The date of pupation and emergence of overwintering larvae in Ostrinia furnacalis under natural conditions(n=140 in 2012,and n=572 in 2013)
3 结论与讨论
本实验3年的田间试验结果揭示了亚洲玉米螟8月下旬初孵化幼虫有少部分进入滞育,8月下旬末孵化幼虫的滞育率达到50%,9月3日及以后孵化的幼虫全部进入滞育。在南昌地区8月下旬至9月上旬的温度仍相当高,日平均气温一般高于25℃。这表明,在此段期间内,不存在低温诱导的滞育问题,显然,亚洲玉米螟田间滞育的发生主要是由秋季逐渐缩短的日照所诱导。该种群滞育诱导的光周期反应已表明,即使在28℃高温的条件下,绝大多数个体在11~13 h短日照下也被诱导进入滞育[10]。亚洲玉米螟这种滞育形成的机制有其重要的生态意义,它确保了亚洲玉米螟在秋季较高的温度下进入滞育,避免了下一代的出现,因为继后的环境条件不利于下一代幼虫的存活。这种滞育特点也确保了该虫能够有足够的虫源越冬,致使该虫的第一代就能够对玉米造成较严重的为害。本实验结果也揭示了越冬个体第二年春季成虫的羽化分散在很长时间,从4月下旬至6月上旬均有羽化。这种分散羽化导致了该虫化性发生变化,最早羽化的个体一年能繁殖5代,而最迟羽化的个体仅繁殖3代;越冬个体的分散繁殖也导致了田间从6月下旬开始就出现世代重叠现象。这些研究结果揭示了该虫复杂的生活史特性,有助于我们准确预测该虫的发生期及防治策略的制定。
本实验中亚洲玉米螟南昌种群(低纬度)各虫态的发育起点温度分别为卵9.24℃;雌幼虫9.85℃、雄幼虫11.90℃、两者的发育起点温度存在较大差异,这可能是由于幼虫发育历期个体差异所导致;雌蛹 12.02 ℃、雄蛹 11.93 ℃,有效积温分别为 76.19,440.16,378.73,107.36,106.84日·度。夏新等[11]报道,亚洲玉米螟黑龙江省双鸭山种群(高纬度),各虫态的发育起点温度分别为卵8.45℃、幼虫5.46℃、蛹 6.09 ℃,有效积温分别为 85.63,517.56,160.21日·度。亚洲玉米螟高纬度(黑龙江)种群的卵、幼虫、蛹的发育起点温度均分别低于低纬度(江西)种群,而前者的有效积温均分别高于后者,这与槐尺蠖(Semiothisa cinerearia)研究结果一致[12-14]。进一步探明了亚洲玉米螟的发育起点温度和有效积温存在地理变异现象。
[1]《中国农作物病虫害》编辑委员会.中国农作物病虫害(上册)[M].北京:农业出版社,1979:492-502.
[2]杜正文,蔡蔚琦.玉米螟在江苏光周期的反应初报[J].昆虫学报.1964,13:129-132.
[3]刘宁,文丽萍,何康来,等.不同地理种群亚洲玉米螟抗寒力研究[J].植物保护学报,2005,32:163-168.
[4]弓惠芬,陈霈,王瑞,等.光周期和温度对亚洲玉米螟滞育形成的影响[J].昆虫学报,1984,27:280-286.
[5]鲁新,李建平.亚洲玉米螟化性类型的初步研究[J].玉米科学,1995,3,75-78.
[6]王振营,鲁新,何康来,等.我国研究亚洲玉米螟历史、现状与展望[J].沈阳农业大学学报,2000,31:402-412.
[7]马瑞,钱海涛,董辉,等.不同地理种群亚洲玉米螟越冬幼虫复苏后的发育历期研究[J].湖北农业科学,2008,47:541-543.
[8]沈荣武,薛芳森,朱杏芬.玉米螟化性及田间滞育发生时间的研究[J].江西植保,1988(1):18-19.
[9]乔利,郑坚武,成卫宁,等.不同饲料配方对亚洲玉米螟生长发育和繁殖的影响[J].西北农林科技大学学报:自然科学版,2008,36(5):109-112.
[10]Xia Q W,Chen C,Tu X Y,et al.Inheritance of photoperiodic induction of larval diapause in the Asian corn borer Ostrinia furnacalis[J].Physiological Entomology,2011,37:185-191.
[11]夏新,丛斌,宋立秋,等.不同地理种群和不同化形亚洲玉米螟的有效积温和存活率的研究[J].安徽农业科学,2007,35(19):5795-5797.
[12]檀华蓉,唐燕平,蔡燕华,等.槐尺蛾卵期及幼虫期的预测预报[J].生物数学学报,1997,12(4):381-384.
[13]蔡燕苹,王翠英,宋凯,等.槐尺蛾预测预报的研究[J].安徽农业大学学报,1998,25(3):240-243.
[14]柳丽婷,苏宝玲,刘广纯,等.国槐尺蠖发育起点温度与有效积温[J].昆虫知识,2010,47(1):126-128.
