氮素对粳稻花后光合响应曲线与氮素利用的影响
2014-11-06刘小龙柯希欢
金 磊,李 霞,刘小龙,柯希欢
(江苏省农业科学院 粮食作物研究所/江苏省优质水稻工程技术研究中心,江苏 南京 210014)
氮肥是影响水稻(Oryza sativa L.)光合作用的重要营养元素,水稻籽粒产量的高低不仅取决于抽穗后光合产物的多少,而且取决于抽穗后光合同化能力的高低[1]。增施氮肥虽带来了水稻单产的持续增加[2],但过高的氮肥可能增加水稻倒伏的危险[3]。最近 Liu等[4]通过研究,系统揭示了过去 30年(1980—2010)来我国氮沉降动态及其与人为活性氮排放的关系,表明:其中农田施肥不合理是人为增加活性氮向大气排放的主要方式,影响了整个生态系统。因此,在现代水稻生产中如何合理调控氮营养,发挥其最大的产量潜力,是现代水稻生产重要的科学问题。
水稻对氮素利用率都存在极显著的品种间差异[5]。魏海燕等[6]和李敏等[7]均通过选用不同类型的产量和氮素差异的粳稻品种,研究在各自最适氮素水平下,研究了叶片光合、衰老特性的差异及其与氮效率的关系,表明维持水稻的净光合速率和延缓衰老是促进水稻高产品种进一步提升氮效率的重要途径[8]。以往的水稻氮效率光合特性研究大多是统一在同一氮素水平,主要是对光合参数的测定,而不同施氮量对不同光强和CO2浓度的光合作用响应则鲜见报道。因此,选择不同的氮水平,研究不同氮效率水稻基因型的光合作用特征,则是判断不同光强和CO2条件下叶片光合性能的发挥以达到高产与减氮协同的关键。本研究选取了8个来自江苏粳稻材料作为研究对象,研究不同施氮量在开花后不同时期对其光强和CO2光合响应曲线的影响,解析其在不同氮肥下光合作用参数与产量表现的关系,进而明确不同施氮量对粳稻光合作用的调节作用,旨为水稻生产中氮肥的合理施用和更高产量的形成提供重要理论依据。
1 材料与方法
1.1 试验材料
供试水稻(Oryza sativa L.)品种为8个,其中迟熟中粳包括南粳45,武育粳3号(中粳对照),02102,早熟晚粳包括晚粳武运粳7号(晚粳对照),宁74142,南粳44和南粳5055,中熟晚粳包括南粳46,水稻种子来自江苏省农业科学院粮食所。
1.2 试验设计
试验于2011—2012在江苏省农业科学院粮食作物研究所网室进行,土壤为马干土(含N 0.083%,P2O50.12%,K 1.00%),采用盆栽的方法,将田间采集土样风干破碎后过2 mm筛,混匀分别装于直径25 cm、高25 cm的塑料桶中(每桶6 kg)。每个品种,每个处理均设3个桶,每桶栽5株。试验参照李霞的方法[9],根据大田的施氮量(135.0 kg/hm2(LN)、180.0 kg/hm2(MN)、270.0 kg/hm2(HN))、栽培塑料筒的面积以及装土的质量换算,设3个施氮量处理,在水稻种植前分别按照每千克土壤中0.11 g N(LN处理)、0.15 g N(MN 处理)、0.22 g N(HN 处理)、0.10 g P 和 0.12 g K,氮肥为尿素(有效氮为 46%),磷肥为过磷酸钙(含13%P2O5),钾肥为氯化钾(含钾为52%)。氮肥作基肥、分蘖肥、保花肥的施用比例分别占总氮量的60%、30%、10%。磷肥一次性基肥施入,钾肥作基肥和茎长粗肥施用,各占50%。将水稻种子用药剂消毒后,浸种72 h后催芽,每年5月16日直接播种于桶中,湿润育秧。水分管理和农药等按常规栽培要求实施。
1.3 测定项目与方法
在开花后不同天数分别测定植株的各生理和生长指标,并在成熟期选择3株,考查供试材料产量结构。
1.3.1 叶片的光强-光合曲线 按李霞等[10]的方法,采用美国LI-COR公司生产LI-6400便携式光合测量系统,用6400-02B LED红蓝光源叶室进行连体叶片瞬时净光合速率(instantaneous net photosynthetic rate,IAPR)测定。将红蓝光源LED设定一系列光合光通量密度(photosynthetic photo flux density,PPFD)梯度,测定 PPFD 分别为 0,50,100,150,200,400,600,800,1 200,1 400 μmol/(m2·s)时叶片的净光合速率(net photosynthetic rates,Pn),每点测定4个重复,绘出光强-净光合速率响应曲线(light intensity-Pn),计算出光饱和条件下最大净光合速率(maximum net photosynthetic rate,Pnmax)。表观量子效率(apparent quantum yield,AQY)在光强为0~200 μmol/(m2·s),分别记录净光合速率,然后用直线回归求出AQY。
1.3.2 叶片的二氧化碳(CO2)-光合曲线和表观羧化效率(apparent carboxylation efficiency,ACE)测定按李霞的方法[11],测定时美国LI-COR公司生产LI-6400便携式光合测量系统,用6400-02B LED红蓝光源叶室进行连体叶片瞬时净光合速率(instantaneous net photosynthetic rate,IAPR)测定。控制PPFD(800±1)μmol/(m2·s)、温度(32±1)℃,外接钢瓶供应 CO2,浓度设定为:0,20,50,100,150,200,250,300,400,600,800,1 000 μmol/mol,用 SPSS 10.0 软件将所得数据绘出 Pn 对 CO2的响应曲线(CO2-Pn),求得CO2补偿点、CO2饱和点以及饱和CO2浓度下的Pnmax,并用CO2浓度低于250 μmol/mol的数据绘出响应直线,直线的斜率为ACE。
1.4 统计分析
采用SPSS 17.0软件,通过单因素方差分析Duncan检验来分析数据的差异显著性和Pearson系数分析参数的相关性。
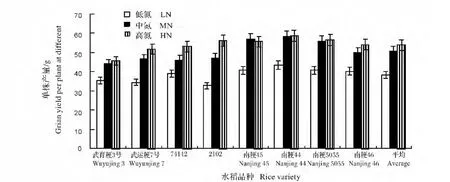
图1 不同施氮下供试水稻的单株产量Fig.1 Grain yield per plant at different N application in different rice varieties

图2 供试水稻不同氮效率分布Fig.2 Distribution of difference in N use efficiency of the rice cultivars
2 结果与分析
2.1 不同施氮量供试材料的产量表现
从图1看出,随施氮量的增加,与中氮相比,高氮的单株产量增加不明显,大多数表现为产量下降。按照Koutroubasa等[12]的计算方法,进一步将本研究中的中粳和晚粳材料进行氮素分析见图2。与对照武育粳3号和武运粳7号相比,供试材料在高氮条件下可以获得较高的产量,其中南京44,南粳5055,南粳45和南粳46在LN处理也可获得较高产量,是氮素双高效类型。
2.2 不同氮肥下供试材料开花后14th和35th剑叶净光合速率对光强响应
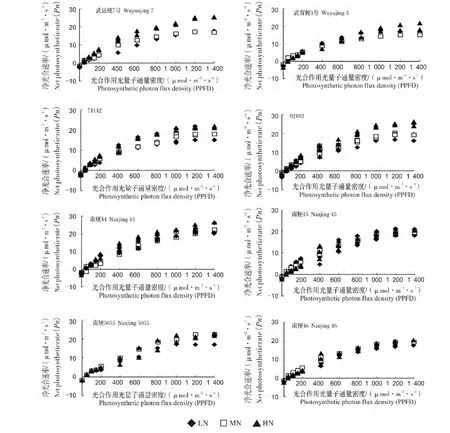
图3 不同施氮量下不同水稻品种剑叶开花后14th光合-光强响应的变化Fig.3 Changes in photosynthetic rate to PPFD of flag leaves of 8 rice varieties in 14thafter flowering under the different Nitrogen applications
2.2.1 不同氮肥下供试材料开花后14th剑叶净光合速率对光强响应 通过进一步测定不同水稻材料在开花后14th剑叶的光合作用对光强的响应曲线,从图3可以看出,高氮提高了武运粳7号和武育粳3号高光强下[>800 μmol/(m2·s)]的Pn,而在中等和低光强下则差异不明显,而对于其他氮素双高效类型南粳45,南粳44,南粳5055以及南粳46则在不同氮素施用下,Pnmax差异不大。施氮对表观量子效率(AQY)影响见表1,同样施氮可部分提高武运粳7号、武育粳3,74142和02102高氮下的AQY,而对于双高效类型则影响不显著。

表1 不同施氮量下不同水稻品种剑叶开花后14th表观量子效率的变化Tab.1 Changes in apparent quantum yield of eight rice varieties in 14th after flowering under the different nitrogen applications

图4 不同施氮量下不同水稻品种剑叶开花后35th光合-光强响应的变化Fig.4 Changes in photosynthetic rate to PPFD of flag leaves of 8 rice varieties in 35thafter flowering under the different Nitrogen applications
2.2.2 不同氮肥下供试材料开花后35th剑叶净光合速率对光强响应 从图4可以看出,与开花后14th相比,施氮量对开花后35th的光强-光合曲线的影响明显,高氮处理可维持剑叶Pn下降较少,光合保持相对稳定,但是不同类型的水稻,其光强-光合曲线则表现不同。对照材料武运粳7号和武育粳3号,随着施氮量的增加,Pn明显增加,对于高氮高效型水稻74142和02102,高氮显著提高其Pn,MN和LN差异不明显,而对于氮双高效类型则是随氮素施用的增加Pn变化不明显,在3个氮素施用条件下均呈现类似的光-光合响应曲线,对氮素的施用不敏感。进一步分析其光合参数(表3),表观量子效率(AQY)随施氮量的增加而增加,Pnmax和光饱和点(LSP)的变化趋势也是类似。而光补偿点(LCP)则随施氮量的增加不同类型表现有差异,与MN相比,对照材料武运粳7号和武育粳3号以及高氮高效型水稻74142和02102均在LN和HN下LCP均较高。而氮双高效类型则LCP在不同的施氮量下变化不大,可见,在灌浆后期,高氮量对这些氮双高效类型维持较高的光合能力关系不大。
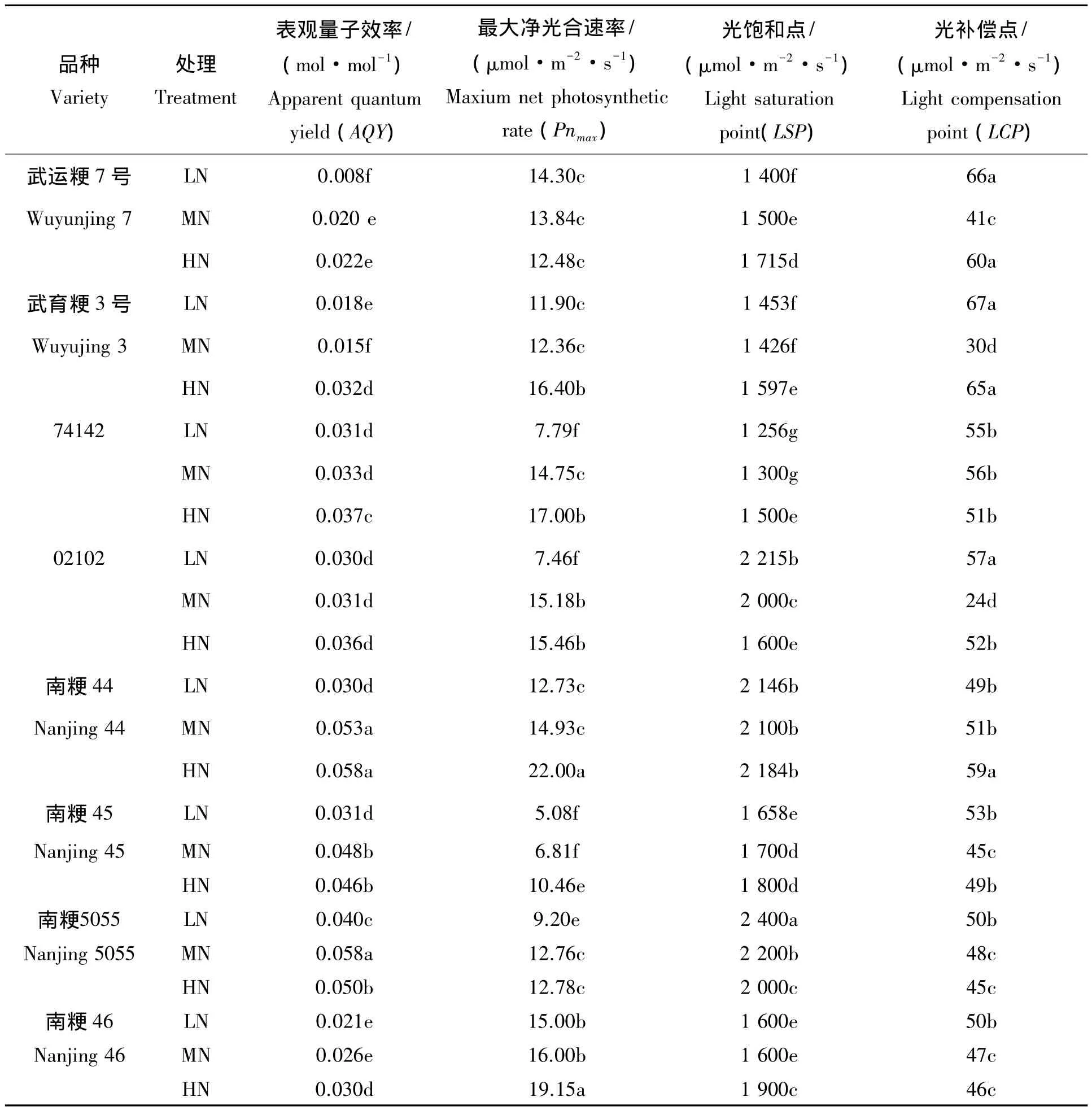
表2 不同施氮量下不同水稻品种剑叶开花后35th的光强-光合响应曲线特征参数Tab.2 Parameters of rice flag leaf photosynthetic light response curve of eight rice varieties in 35thafter flowering under the different nitrogen applications
2.3 不同氮肥供试材料剑叶开花后14th剑叶和35thCO2-光合作用响应曲线
2.3.1 不同氮肥供试材料剑叶开花后14th剑叶CO2-光合作用响应曲线 CO2也是光合作用底物之一,水稻光合作用对CO2浓度的利用也影响着其产量的形成。在一定CO2浓度范围内,植物的Pn随CO2浓度的增加而增加。图5给出了在不同施氮量下水稻剑叶开花后14th的光合作用对不同CO2浓度的依赖关系。从结果可以看出,与光强-光合曲线相比,施氮对CO2-光合曲线改变比较多,随着施氮量的增加,不同水稻品种Pn在不同浓度的CO2下均显著增加。从CO2-光合参数可进一步分析(表2),表观羧化效率(ACE)随着氮素的增加而增加,但是N和HN下,则增加不明显,表明在低氮下,Rubisco对CO2的羧化能力对氮素的依赖更大,很容易受到影响,CO2的饱和点(CSP)也随氮素的增加而增加,其中在HN下达到最大,CO2补偿点(CCP)则是水稻固有的属性,施氮对其影响不明显。有趣的是对于氮素双高效的水稻类型如南粳45,南粳44,南粳5055以及南粳46,它们在高氮的条件下,CCP则显著低于LN的处理,可见,其在高氮条件下,其光合作用具有较宽的CO2作用范围,也是其氮高效的重要生理基础。

图5 不同施氮量下供试水稻材料剑叶在开花14th时光合-CO2响应曲线的变化Fig.5 Changes in photosynthesis-CO2response curves in the flag leaves of different rice varieties in 14thafter flowering under the different nitrogen applications

表3 不同施氮量下不同水稻品种剑叶开花后14th的CO2-光合响应曲线特征参数Tab.3 Parameters of rice flag leaf photosynthetic CO2response curve of eight rice varieties in 14thafter flowering under the different nitrogen applications.
2.3.2 不同氮肥下供试材料开花后35th剑叶净光合速率对CO响应 从图6可以看出,与开花14th的2CO2-光合曲线相比,开花35th的则表现不同,在HN下Pn的优势均不明显,而与MN的类似,LN的下降比较多,各品种间表现类似。进一步分析光合参数(表4),可见,与开花14th的相比,开花后羧化效率下降不多,但是Pnmax、CO2饱和点以及CO2补偿点均明显下降,其中在低氮和高氮下下降显著,但中氮下,其Pnmax下降不明显,光合参数的变化暗示此时植株已遭受逆境,但是提高CO2浓度,可以缓解植物因叶片衰老导致的光合能力下降,表现为高氮和中氮的光合参数类似,低氮的则较低。

图6 不同施氮量下供试水稻材料剑叶在开花35th时光合-CO2响应曲线的变化Fig.6 Changes in photosynthesis-CO2response curves in the flag leaves of different rice varieties in 35thafter flowering under the different Nitrogen applications
2.4 不同施氮量供试水稻生理参数与单株产量聚类分析
产量是对整个生育期氮素利用效率的综合表现的指标,因此,本研究结合单株产量,进一步经过WARD法聚类分析(度量标准为Euclidean距离),以阈值为5作为聚类标准,在不同氮素下,可将供试材料分为2个类别(表5),但未能从供试材料中鉴定出具有高光合效率和高产的氮素高效利用的材料类型,可见供试材料的氮素利用能力还需改良。从产量角度,其中在LN下,类型1为氮素利用效率略好的类型,并表现在开花后14 d对高光强和开花后35 d对高CO2的利用能力较强,但是在开花14 d对高CO2以及开花35 d对高光强利用能力则小于类型2;而在MN下,从产量看,类型2为氮素利用效率略好的类型,并表现在开花后14 d对高光强和开花后35 d对高CO2的利用能力较强,但是其他光合参数则差异不大;在HN下,从产量的角度,类型2为高效型,其中表现在开花后14 d对高光强和高CO2以及开花后35 d对高光强的利用能力较强,但是在开花35 d对高CO2利用能力则小于类型1,可见不同施氮量下,供试材料对光强的响应能力调节作用比较明显,而对CO2的利用能力则调控有限。值得注意的是,南粳44和南粳5055则是对LN和HN都表现效率较高的材料。

表4 不同施氮量下不同水稻品种剑叶开花后35th的CO2-光合响应曲线特征参数Tab.4 Parameters of rice flag leaf photosynthetic CO2response curve of eight rice varieties in 35thafter flowering under the different Nitrogen applications.

表5 供试水稻对氮素利用能力的联合聚类分析Tab.5 Cluster analysis to N use ability in different rice materials
3 结论与讨论
水稻产量形成的关健时期,氮素是通过与光合作用有关酶的活化、CO2的活化及光系统单位和电子传递链构成来影响和调节光合作用,从而对水稻干物质生产和产量形成的影响[13]。Moll等[14]认为在低氮条件下主要是所积累的利用效率不同所致,即营养体氮向籽粒氮再转移的不同;而在高氮条件下,则主要是氮吸收效率起主要作用。水稻剑叶的光合特性与植株的氮肥吸收利用效率关系密切,当籽粒需求大量的光合产物供应时,可能会促使茎蘖储藏的物质过早地向籽粒运转,植株上部的叶片的光合功能却在迅速衰退,以至于作物出现减产[15],可见水稻叶片的衰老,不仅减少光合物质的积累,氮素吸收利用效率也随之下降。提高剑叶水稻光合生产能力主要体现在:1)提高单位面积的光合生产能力(净光合速率);2)保证叶片高光合能力的持续,即防止叶片光合功能的早衰,可见水稻的氮素利用效率不仅决定于施氮量,而且与水稻的基因型以及剑叶的发育时期有关。本文的研究发现:随着施氮量的增加,供试材料14th的Pnmax均增强。与中氮相比,在高氮下开花后14thPnmax的增幅不明显。而与开花后14th相比,供试材料在35th的Pnmax均下降,而且低氮条件下下降更明显,加剧了不同光强下中氮和高氮处理的Pn差距,而且高氮的则降低较少,可见,增施氮素有利于维持剑叶后期的光合功能稳定,可增强对生育后期逆境(寒露风等)的抵抗能力,表现保持光合作用稳定[16]。但氮双高效类型则在不同施氮量下Pn差异不大,表现对施氮并不敏感,表现在不同氮素下,表现类似的Pnmax,对氮肥的依赖较小,可见,选择氮双高效的水稻类型有利于实现减氮和高产的目标。
CO2也是光合作用的底物之一,随着全球气候变暖以及人类作物生产对氮素的依赖,势必使植物遭遇氮素和CO2浓度互作的影响。本文的研究表明:在开花14th,不同施氮量对供试水稻CO2-光合曲线的影响显著大于光强-光合曲线,均在高CO2浓度下显著提高了Pnmax,可进一步发挥其最大的光合潜力;但在开花后35th,与光强-光合曲线相比,CO2-光合曲线则表现不同,虽然在不同施氮量下,虽显著地降低了Pnmax,但是不同浓度施氮量下,供试水稻光合作用对CO2的响应能力则变化不明显。考虑到本研究产量的结果,虽然在中氮和高氮处理下,其单株产量均比低氮的单株产量提高,但与中氮相比,高氮处理对本文的供试粳稻的增产效果不明显。从不同氮素的光合作用特征曲线进一步看出,氮双高效类型高产粳稻,增施氮肥增加了前期的CO2的利用能力,而后期则增加了光强的利用能力,可见,结合未来大气CO2浓度的增加趋势和氮肥合理施用环境友好的要求,选取氮双高效的类型,可适当降低氮素的施用,发挥其对CO2的同化能力,可最大限度地发挥高产水稻的光合潜力,即可达到稳产高产,这可能是其高产栽培需要关注的重要方面。
[1]马群,杨雄,李敏,等.不同氮肥群体最高生产力水稻品种的物质生产积累[J].中国农业科学,2011,44(20):4159-4169.
[2]马群,李国业,顾海永,等.我国水稻氮肥利用现状及对策[J].广东农业科学,2010(11):126-129.
[3]杨惠杰,房贤涛,何花榕,等.福建超级稻品种茎秆结构特征及其与抗倒性和产量的关系[J].中国生态农业学报,2012,20(7):909-913.
[4]Liu X J,Zhang Y,Han,et al.Enhanced nitrogen deposition over China[J].Nature,2013,494:459-462.
[5]李敏,张洪程,李国业等.水稻氮效率基因型差异及其机理研究进展[J].核农学报,2011,25(5):1057-1063.
[6]魏海燕,张洪程,马群,等.不同氮肥吸收利用效率水稻基因型叶片衰老特性[J].作物学报,2010,36(4):645-654.
[7]李敏,张洪程,杨雄,等.高产氮高效型粳稻品种的叶片光合及衰老特性研究[J].中国水稻科学,2013,27(2):168-176.
[8]李霞,王超,任承钢,等.植物内生真菌B3和不同施氮量对水稻生长和产量的影响[J].江苏农业学报,2009,25(6):1207-1212.
[9]Li X,Cao K,Wang C,et al.Variation of photosynthetic tolerance of rice cultivars(Oryza sativa L.)to chilling temperature in the light[J].African Journal of Biotechnology,2010,9(9):1325-1337.
[10]李霞,戴传超,焦德茂,等.光照条件下低温对水稻籼粳亚种幼苗抗氧化物质含量的影响[J].植物生理与分子生物学学报,2006,32(3):345-353.
[11]Koutroubas S D,Ntanos D A.Genotypic differences for grain yield and nitrogen utilization in indica and japonica rice under mediterranean conditions[J].Field Crops Research,2003,83:251-260.
[12]Ladha J K,Pathak H,Krupnik T J.Efficiency of fertilizer nitrogen in cereal production:retropects and prospects[J].Advances in Agronomy,2005,87:85-156.
[13]Moll R H,Kamprath E J,Jackson W A.Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization[J].Agronomy Journal,1982,74:562-568.
[14]翟虎渠,曹树青,万建民.超高产杂交稻灌浆期光合功能与产量的关系[J].中国科学:C辑,2002,32(3):211-217.
[15]焦德茂,李霞,黄雪清,等.不同高产水稻品种生育后期叶片光抑制、光氧化和早衰的关系[J].中国农业科学,2002,35(5):487-492.
