亲子鉴定中四配子异源嵌合体报道1例
2014-10-26陈芳,陈俭,付颖,黄琼
陈 芳,陈 俭,付 颖,黄 琼
(浙江大学,浙江 杭州 310000;浙江大学司法鉴定中心,浙江 杭州 310000)
1 简要案情
2013年6月,当事人李某带着女儿来鉴定中心,要求鉴定是否存在亲生血缘关系,在案件受理过程中,当事人提出曾经去两家鉴定机构做过亲子鉴定,且两家鉴定机构鉴定结论不一致,要求重新鉴定。按照知情同意原则,采集了被鉴定人三份不同的检材,分别是血样,口腔拭子及带毛囊毛发,并要求当事人提供孩子生母血样。当事人李某(以下称父亲)、孩子生母(以下称母亲)及女儿无输血史,无骨髓移植及放疗化疗史等,李某女儿外观正常,女性特征明显。
2 检验方法
2.1 染色体核型检测
外周血淋巴细胞培养[1],常规制备染色体,G显带,镜下计数及分析核型。
2.2 DNA提取
采用常规Chelex-100法[2]分别提取父亲、母亲及女儿血样、口腔拭子、带毛囊毛发检材DNA,分别采用GoldeneyeTM20A试剂盒(北京基点认知),Identifiler试剂盒(美国AB公司),AGCU21+1试剂盒(无锡中德美联)进行39个常染色体STR基因座的分型,采用GoldeneyeTM20Y试剂盒(北京基点认知)进行20个Y-STR基因座的分型,采用Investigator Argus X-12试剂盒(德国QIAGEN)进行12个X-STR基因座的分型,扩增体系和扩增程序按相关操作手册进行。
2.3 DNA分型
采用9700型PCR仪(美国AB公司)进行PCR扩增,使用3130型遗传分析仪(美国AB公司)进行电泳分离,使用 GeneMapper ID v3.2.1 软件(美国 AB 公司)处理电泳数据,实验过程参照操作手册和相关文献。
3 检验结果
3.1 染色体核型分析
外周血淋巴细胞:核型为46,XX/46,XY两个细胞系(见图 1),其中 46,XX(54%),46,XY(46%)。

图1 李某女儿核型分型图,显示有两个细胞系,分别为46,XX/46,XY
3.2 STR基因分型
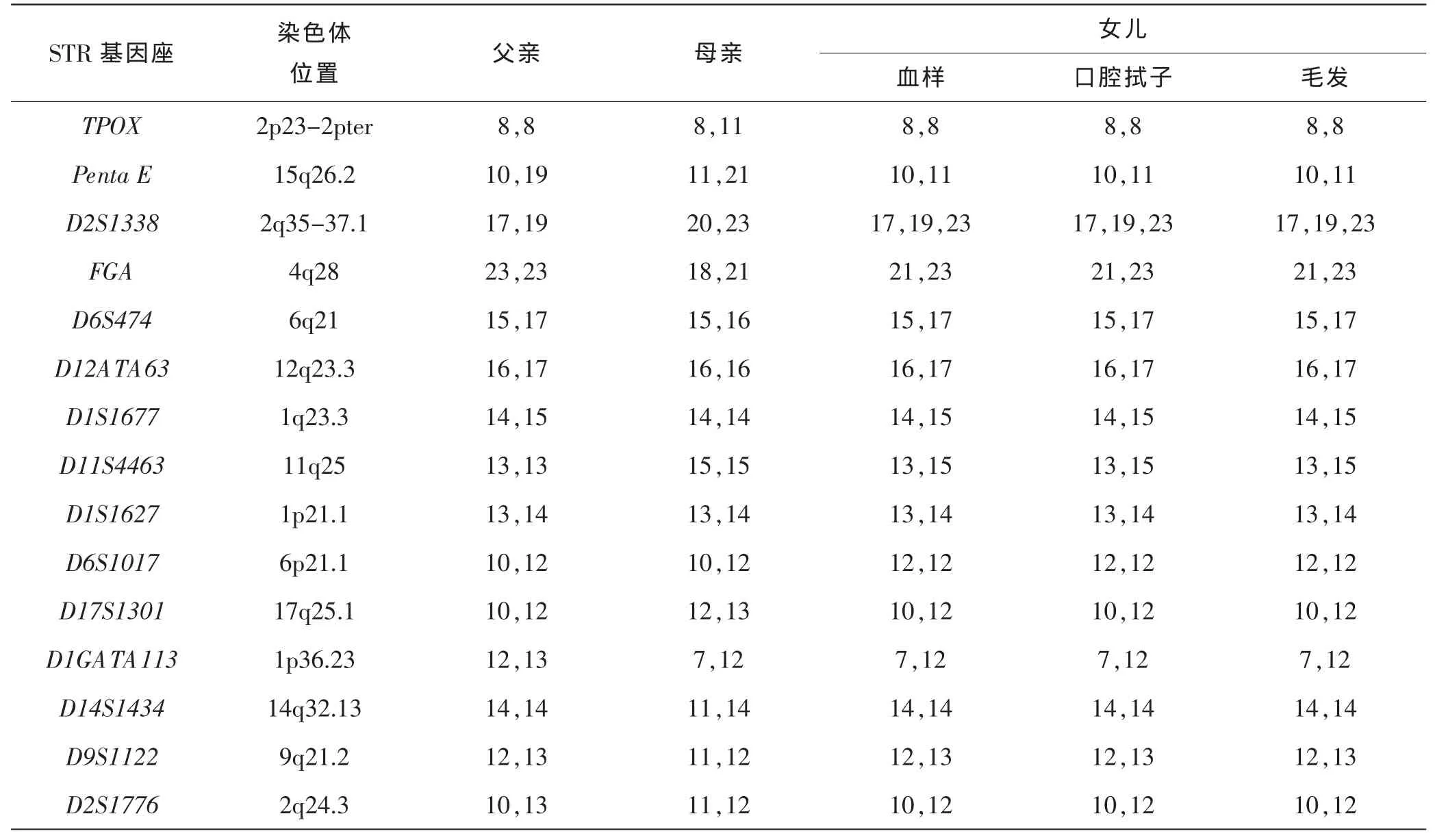
在 39个常染色体 STR检测中(见表 1~2),父亲及母亲常染色体STR均正常。女儿血样、口腔拭子、毛发检材基因分型一致,在D19S433、D21S11、D18S51、CSF1PO、D8S1179、D2S1338、D22S1045 等 7个基因座出现三等位基因,且在D5S818、D3S1358、D2S441、D1S1677等4个基因座出现等位基因峰高不均衡现象,并且Amelogenin基因座均扩增出等位基因Y,且等位基因X和Y峰高不均衡(见图2)。

图2 依次为父亲、母亲、女儿血样、口腔拭子、毛发常染色体STR分型图
D19S433基因座,有3个等位基因12.2/13/14,峰高不均衡,父亲的两个等位基因12.2/14全部遗传给子代,等位基因13来源于母亲;D21S11基因座,有3个等位基因31/32.2/33.2,峰高不均衡,父亲的2个等位基因31/33.2全部遗传给子代,等位基因32.2来源于母亲;D18S51基因座,有3个等位基因16/19/21,峰高不均衡,父亲的2个等位基因16/19全部遗传给子代,等位基因21来源于母亲;CSF1PO基因座,有3个等位基因10/11/12,峰高不均衡,父亲的2个等位基因10/11全部遗传给子代,等位基因12来源于母亲,D8S1179基因座,有3个等位基因10/12/15,峰高不均衡,父亲的2个等位基因10/15全部遗传给子代,等位基因12来源于母亲;D2S1338基因座,有3个等位基因17/19/23,峰高不均衡,父亲的2个等位基因17/19全部遗传给子代,等位基因23来源于母亲;D22S1045有3个等位基因11/15/16,峰高不均衡,父亲的2个等位基因11/16全部遗传给子代,等位基因15 来源于 母 亲 ;D5S818、D3S1358、D2S441、D1S1677等基因座,存在等位基因不均衡的现象,因父亲与母亲有一个相同的等位基因,父亲的等位基因全部遗传给子代,母亲遗传给子代的等位基因与父亲相同,故峰高比接近3∶1。以上11个基因提示女儿在血样、口腔拭子及毛发中存在着明显的嵌合现象,存在着两个不同的细胞系(46,XX/46,XY),且两个细胞系所占有的比例不一致。

表1 父亲、母亲、女儿血样、口腔拭子、毛发常染色体近着丝粒端STR分型结果

表2 父亲、母亲、女儿血样、口腔拭子、毛发常染色体末梢STR分型结果
续表2
分别对血痕,口腔拭子和毛发检材采用GoldeneyeTM20Y试剂盒进行19个Y-STR基因座扩增,女儿血样、口腔拭子及毛发检材均扩增出等位基因,且基因座分型与父亲基因座分型一致。
分别对血痕,口腔拭子和毛发检材采用Investigator Argus X一12试剂盒进行12个X-STR基因座扩增,女儿血样、口腔拭子及毛发X-STR分型结果除Amelogenin基因座外,其它基因座分型结果一致,女儿血样、口腔拭子的Amelogenin基因座等位基因为X/Y.峰高不均衡,而毛发Amelogenin基因座未见Y等位基因,除DXS10146基因座外,女儿血样,口腔拭子及毛发检材等位基因均能从父亲及母亲中找到来源,DXS10146基因座女儿等位基因为29/30,父亲等位基因为30,母亲等位基因为30/32,不符合孟德尔遗传定律,等位基因29找不到来源,推断女儿在DXS10146基因座发生基因突变。
12个X-STR基因座检测中,女儿血样Amelogenin基因座检测出Y等位基因,说明有两个细胞系存在,而其它基因座峰高比基本均衡,女儿口腔拭子Amelogenin基因座也检测出Y等位基因,说明有两个细胞系存在,其他基因座等位基因峰高不均衡,来自父亲的等位基因峰高明显比来自母亲的等位基因峰低,口腔拭子中46,XY细胞系所占的比例比血样中46,XY细胞系所占比例高。女儿毛发检材Amelogenin基因座未检测出Y等位基因,且等位基因峰高均衡,未见明显嵌合现象存在。
通过对女儿血样,毛发常染色体STR、Y-STR及X-STR结果的分析,以上组织均有2个细胞系(46,XX/46,XY)存在,在39个常染色体STR基因座中,均能清楚的显示其遗传方式,其中有11个基因座出现三等为基因,三等位基因中有2个是来源父亲的,1个是来源于母亲的,且来源于母亲的等位基因峰高比来源于父亲的等位基因峰高两倍,所有的39个常染色体STR基因座均只有1条母源性等位基因,没有1个基因座显示有2个母源性等位基因。12个X-STR基因座中,等位基因1个来源于父亲,1个来源于母亲,并且部分基因座峰高比接近2∶1,所有的常染色体基因座没有出现四等位基因,X染色体也没有出现三等位基因。
4 分析讨论
嵌合体也被叫做奇美拉(chimera),在希腊神话中,奇美拉是狮头,羊身,蛇尾的吐火怪物,在医学界,它指的是人体中的遗传物质并非只是来自一个细胞系,可能来自于两个或者以上的细胞系所构成的生物体,可自发产生或人工产生。人类嵌合体是极其罕见的,一般是在血型检测中发现有2种红细胞群体或真两性畸形来就诊而被发现的。
嵌合体在生物学上可以分为两种,同源嵌合体和异源嵌合体。嵌合成分来源于同一受精卵的嵌合体称为同源嵌合体,自发或者诱发导致的基因突变以及染色体畸变均可导致同源嵌合体的产生。由2个不同的受精卵融合而形成的个体为异源嵌合体,异源嵌合体[3]可分为人造异源嵌合体(artificial chimerism)、孪生子异源嵌合体(tiwn chimerism)和四配子异源嵌合体(tetragametic chimerism)。人造异源嵌合体大部分是医源性的,是由于外周造血干细胞的输入、异源骨髓移植或器官移植导致的,也可由其他原因产生。异卵双生导致的血型嵌合是天然的奇美拉,主要是胎盘融合或者子宫内造血干细胞的交换而导致的,异卵双生的孪生子异源嵌合体细胞嵌合仅限于血细胞中,为两种合子来源的细胞嵌合,据统计血型奇美拉8%来自双胞胎,21%来至于三卵多胞胎[3]。在人类中四配子异源嵌合体是极其罕见的,常常由于两性畸形患者就诊或者ABO检测时发现2种红细胞群体而被发现。由于两个合子融合并最终发育成的一个个体而导致的四配子异源嵌合体,这种四配子异源嵌合个体在遗传学上含有两种独立的细胞系(如46,XX/46,XY细胞系),在鼠类[4-7]和其他哺乳动物[8]中均发现这种四配子嵌合体,在人类[9,26,28]同样也已发现。 根据文献推测有4种机制可以导致四配子异源嵌合体的形成[10]:(1)两个胚胎的融合;(2)卵细胞和第一极体双受精作用;(3)次级卵母细胞和第一极体双受精作用;(4)卵细胞和第二极体双受精作用。人类四配子异源嵌合体的形成机制在目前的文献报道已发现了4种,第一种为两个胚胎的融合[11-14,26];第二种为卵细胞和第二极体双受精作用[15];第三种为卵细胞经过孤雌分裂,形成两个相同的配子,与两个精子进行双受精[16-17,23,28];第四种为产雄孤雌生殖[20-22]。两个胚胎融合患者常染色体STR检测显示含有父源和母源各两个不同的等位基因,也就是说四个等位基因分别来源于四个配子,随着现代医学辅助生殖技术的发展,体外受精[13-14]和卵胞浆内单精子注射等技术增加了胚胎融合的风险[18]。卵细胞和第二极体双受精应显示在远离染色体着丝粒的位点上有母源的两个不同的等位基因[19],因为染色体要发生交换。卵细胞孤雌分裂形成两个相同的配子双受精,在常染色体STR检测显示含有两个父源的等位基因和一个母源的等位基因,因孤雌分裂形成2个相同的配子,DNA完全相同,故显示只有1个配子成分,也就是说只显示一个母源的等位基因。本案例中女儿有两个细胞群,核型为46,XX/46,XY,STR结果显示有父源的2个不同配子成分和母源的1个配子成分,因此推断为四配子异源嵌合体。这种情况可解释为来自母亲的生殖细胞经孤雌分裂形成2个相同的配子与来自父亲的X精子和Y精子双受精,然后2个合子融合到1个个体,从而形成有2个细胞系的四配子异源嵌合体(见图3)。由母亲生殖细胞经孤雌分裂形成2个相同的配子双受精形成2个合子,经融合后发育成的个体,这种四配子异源嵌合体案例目前报道较少见。
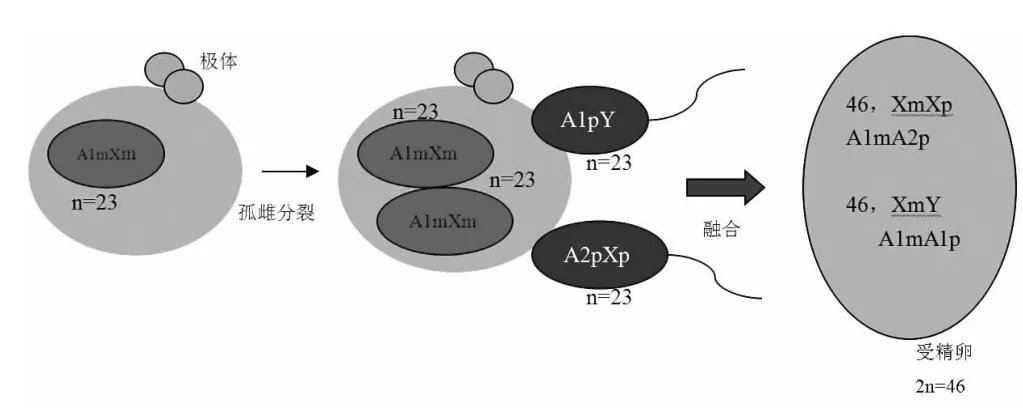
图3 四配子异源嵌合体形成机制图解,母亲的生殖细胞经孤雌分裂形成2个相同的配子与来自父亲的X精子和Y精子双受精,然后2个合子融合到一个个体,从而形成有2个细胞系的四配子异源嵌合体
1953年英国医学杂志(the British Medical Journal)报道了第一例血型嵌合体 “一名女子被发现血液中含有两种不同的血型”[24],1962年Gartler etal.[25]报道了第一例由于双受精作用而导致的人类嵌合体,而美国母亲莉迪娅事件[26]揭开了人类的嵌合体现象。到目前为止,报道的人类嵌合体有50余例[27],随着医学辅助生殖技术的不断发展,人类嵌合体现象越来越常见,目前常用的检测嵌合体的方法主要有三种:(1)ABO 血型基因分型法;(2)HLA(人类白细胞抗原)法;(3)STR法。随着技术的不断发展,新的检测方法也不断在应用,2012年,So Youn Shin等[28]首次采用SNP手段对四配子异源嵌合体进行检测,新的检测微嵌合体技术[29]也在不断发展中。在亲子鉴定中遇到三等为基因或者性分化异常者(存在两个细胞系46,XX/46,XY),在染色体核型分析的基础上,首先要确定是同源嵌合体还是异源嵌合体,采用常染色体STR试剂盒进行检测,如果所有基因座都只显示1个父源等位基因和1个母源等位基因,可判定为同源嵌合体,如果某些基因座出现2个母源等位基因和 (或)父源等位基因可判定为异源嵌合体。确定为异源嵌合体之后需要明确嵌合体发生的机制,首先采用常染色体靠近着丝粒STR基因座进行检测,如果某些基因座出现2个母源等位基因和2个父源等位基因,说明是由于胚胎融合而形成的嵌合体,如果某些基因座出现1个母源等位基因和2个父源等位基因,这时候需要进一步检测,采用常染色体末梢STR位点进行检测,如果所有基因座出现1个母源等位基因,说明是由于母亲生殖细胞孤雌分裂而形成的嵌合体,如果某些基因座存在2个母源等位基因,说明是由于卵子和第二极体双受精形成的嵌合体。
在亲子鉴定实践中,当被检者有2种不同性别的红细胞群体时则容易被发现是嵌合体,而当被检者有2种细胞群性别相同或者只在血液中存在1种细胞群的时候就不容易被发现,值得注意的是,当嵌合体者的血液中只存在1种细胞群的时候,容易造成亲权鉴定结果错误。在亲子鉴定或个体识别中,如若遇到嵌合体现象,应该引起关注,首先要排除污染因素,可通过反复实验,更换不同试剂盒扩增来排除,若仍为嵌合体现象,分析其发生机制,方可作出定论,以保证鉴定结果的准确性和科学性。
[1]Gartler SM,Waxman SH,Giblett E.An XX/XY humanhermaphrodite resulting from double fertilization[J].ProcNatl Acad Sci U SA, 1962,(48):332-335.
[2]Yu N, Kruskall MS, Yunis JJ, etal.Disputed maternity leading to identification of tetragametic chimerism[J].N Engl JMed, 2002,(346):1545-1552.
[3]Vandijk BA, Boomsma DI, Denman AJM.Blood group chimerism in human multiple births is not rare[J].American Journal of Medical Genetics,1996,(61):264-268.
[4]Mintz B,Palm J.Gene control of hematopoiesis.I.Erythrocyte mosaicismand permanent immunological in allophonic mice[J].J Exp Med,1969,129(5):1013-1027.
[5]Tucker EM ,Dain AR,Moor RM.Sex chromosome chimaerism and the transimission of blood group genes by tetraparental rams[J].JRepro Fertil,1978,54(1):77-83.
[6]Fundele RH,Surani MA.Experimental embryological analysis of genetic imprinting in mouse development[J].Dev Genet 1994,(15):515-522.
[7]Kono T,Obata Y,Wu Q,etal.Birth of parthenogenetic mice that can develop to adulthood[J].Nature, 2004,(428):860-864.
[8]Sumantri C,Boediono A,Ooe M, etal.Fertility of sperm from a tetraparental chimeric bull[J].Anim Repod Sci,1997,46(1-2):35-45.
[9]Strain L,Warner JP,Johnston T,etal.A human parthenogenetic chimaera[J].Nat Genet, 1995,(11):164-169.
[10]Verp MS,Harrison HH,Ober C,etal.Chimerism as the etiology 0f a 46,XX/46,XY fertile true hermaphrodite[J].Fertil Steril, 1992,57(2):346-9.
[11]Green AJ, Barton DE, Jenks, etal.chimaerism shown by cytogenetics and dna polymorphism analysis[J].J Med Genet,1994,31(10):816-817.
[12]Uehara S,Nata M,Nagae M,etal.Molecular biologic analyses of tetragametic chimerism in a true hermaphrodite with 46,XX/46,XY[J].Fertil Steril, 1995,(63):189-192.
[13]Bonthron DT.Amalgamation of in vitro fertilized embryos,resulting in birth of a true hermaphrodite chimera[J].Am J Hum Genet, 1997,(61):A147.
[14]Strain L,Dean JC,Hamilton MP,etal.A true hermaphrodite chimera resulting from embryo amalgamation after in vitro fertilization[J].N Engl JMed, 1998,338(3):166-169.
[15]Chen CP, Chern SR, Sheu JC, etal.Prenatal diagnosis,sonographic findings and molecular genetic analysis of a 46,XX/46,XY true hermaphrodite chimera[J].Prenat Diagn,2005,(25):502-506.
[16]Giltay JC,Brunt T,Beemer FA,etal.Polymorphic detection of a parthenogenetic maternal and double paternal contribution to a 46,XX/46,XYhermaphrodite[J].Am J Hum Genet, 1998,62(4):937-40.
[17]Cui Y,Zhu P,Ye X,etal.The mechanism of tetragametic chimerism in a true hermaphroditism with 46, XX/46,XY[J].Zhonghua Nan Ke Xue, 2004,10(2):107-12.
[18]Friedler S, Mashiach S, Laufer N.Births in Israel resulting from in-vitro fertilization/embryo transfer, 1982-1989: National Registry of the Israeli Association for Fertility Research[J].Hum Reprod, 1992,(7):1159-1163.
[19]Malan V,Vekemans M,Turleau C.Chimera and other fertilization errors[J].Clin Genet, 2006,(70):363-373.
[20]Kaiser-Rogers KA,McFadden DE,Livasy CA,etal.Androgenetic/biparental mosaicism causes placental mesenchymal dysplasia[J].JMed Genet, 2006,(43):187-192.
[21]Giurgea I, Sanlaville D, Fournet JC, etal.Congenital hyperinsulinism and mosaic abnormalities of the ploidy[J].J Med Genet, 2006,(43):248-254.
[22]Surti U,Hill LM,Dunn J,etal.Twin pregnancy with a chimeric androgenetic and biparental placenta in one twin displaying placental mesenchymal dysplasia phenotype[J].Prenat Diagn, 2005,(25):1048-1056.
[23]Yamazawa K,Nakabayashi K,Kagami M,etal.2010.Parthenogenetic chimaerism/mosaicism with a Silver-Russell syndrome-like phenotype[J].JMed Genet,(47):782-785.
[24]Seabright M.A rapid banding technique for human chromosomes[J].Lancet,1971,2(7731):971-972.
[25]公安部物证鉴定中心.GA/T383—2002法庭科学 DNA实验室检验规范[S].北京:中国标准出版社,2002.
[26]Bowley C.C,Ann M.Hutchison,Joan S.Thompson,etal.A human blood-group chimera[N].British Medical Journal,2011-10-02(81).
[27]Malan V,Gesny R,Morichon-Delvallez N,etal.Prenatal diagnosis and normal outcome of a 46, XX/46, XY chimera:a case report[J].Ham Reprod, 2007,22(4):1037-41.
[28]Shin SY,Yoo HW,Lee BH,etal.Identification of the mechanism underlying a human chimera by SNP array analysis[J].Am JMed Genet A, 2012,(9):2119-23.
[29]Eun Jeong Won, Hye Ryoen Park, Tae Sung Park.Amplification refractory mutation system-PCR is essential for the detection of chimaeras with a minor allele population:a case report[J].JClin Pathol, 2013,(66):446-448.
