猪DECR1基因的研究进展及其对猪肉品质的影响
2014-10-23堵晶晶张玉托
堵晶晶,张玉托,全 琪,罗 嘉,朱 砺*
(1.四川农业大学动物科技学院,四川 雅安 625014;2. 四川农业大学园艺学院,四川 雅安 625014)
随着生活水平的不断提高,人们对肉质要求也越来越高。猪肉品质很大程度上受到肌肉内脂肪组成比例和含量的影响[1]。线粒体2,4-双烯酰辅酶A还原酶1(2,4-dienoyl-CoA reductase 1, DECR1)在脂类代谢中扮演重要角色[2],随着研究的不断深入,发现猪DECR1基因可能与猪脂肪沉积和肉质性状有直接关系,极有可能成为影响猪肉性状的主效基因或遗传标记。因此,深入研究猪DECR1基因可为肉质研究带来新的突破口,为生猪产业创造巨大的经济利益。
1 猪DECR1基因的结构与定位
DECR1是多不饱和脂肪酸(PUFA)氧化分解途径中的关键限速酶,在多不饱和脂酰基CoA β-氧化过程中起重要作用,可作为猪脂肪沉积和肉质性能的候选基因[3]。人DECR1与猪DECR1基因具有较高的同源性[4],但与人DECR1基因位于8号染色体q21.3区域不同的是,猪DECR1基因位于4号染色体上q1.2区域[5-7],其cDNA全长2 352 bp,包括987 bp的开放阅读 框 (ORF),53 bp的 5’ 非 翻 译 区(UTR)和1 312 bp的3’-UTR;编码328个氨基酸,预测其蛋白分子质量为35.43 ku,等电点为9.37[8]。同时,该基因包括1个启动密码子,1个终止密码子,1个polyA加尾信号(AATAAA),3个addl(ATTTA)基元。但截至目前,尚未见到对猪DECR1基因进行精确定位的报道。
2 猪DECR1基因的差异化表达
2.1 同一品种不同组织中的差异化表达
分析相应的mRNA和蛋白质相对表达量是研究目的基因的常用方法。猪DECR1基因是一个表达谱广泛且组织间表达存在显著差异的基因。李步高等[9]研究了马身猪、大白猪不同组织中DECR1基因的表达,结果表明猪DECR1基因在各组织中都有表达,不同组织的表达量差异极显著(P<0.01),在脂肪、肝脏和小肠中呈高丰度表达。其中,肝脏的表达度最高,而在肺脏、胃与肾脏等组织中表达度相对较低。同时,史利华等[10]研究发现(如表1),滇南小耳猪和藏猪的DECR1基因在脂肪组织(背膘和腹脂)中的表达量高于肌肉组织中的表达量。据此说明,猪DECR1基因在同一品种不同组织中的表达存在较大差异。
2.2 不同品种同一组织中的差异化表达
在生物进化过程中,地理环境差异往往会造成遗传物质的差异化表达,DECR1基因作为一个影响肉质性状的候选基因,在猪上差异化表达的研究报道较少。以大白猪和马身猪作为实验材料,用荧光定量PCR技术检测DECR1基因在不同品种同一组织中的蛋白相对表达量,结果表明DECR1基因在大白猪皮下脂肪组织中表达量极显著高于马身猪(P<0.01),而在小肠、脾脏、心脏中大白猪极显著高于马身猪[9]。其他实验表明[10],肌肉组织中的DECR1基因表达量:大约克夏猪>滇南小耳猪>藏猪,但差异不显著;而腹脂中DECR1基因表达量:滇南小耳猪>藏猪>大约克夏猪,且差异显著(P<0.05)。
2.3 同一组织的不同时段的差异化表达
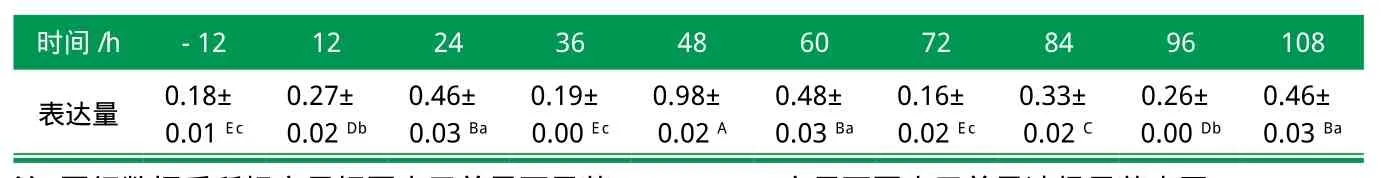
刘海峰等[11]选择猪前脂肪细胞诱导分化过程中的10个不同诱导分化时间点检测了DECR1基因的mRNA表达量,结果显示(如表2):DECR1基因诱导分化后的表达量在48 h时达到最大值,而48 h之前呈现有波动的上升表达,48 h之后为波动式的下降表达。
3 猪DECR1基因的多态性与功能
3.1 猪DECR1基因的多态性
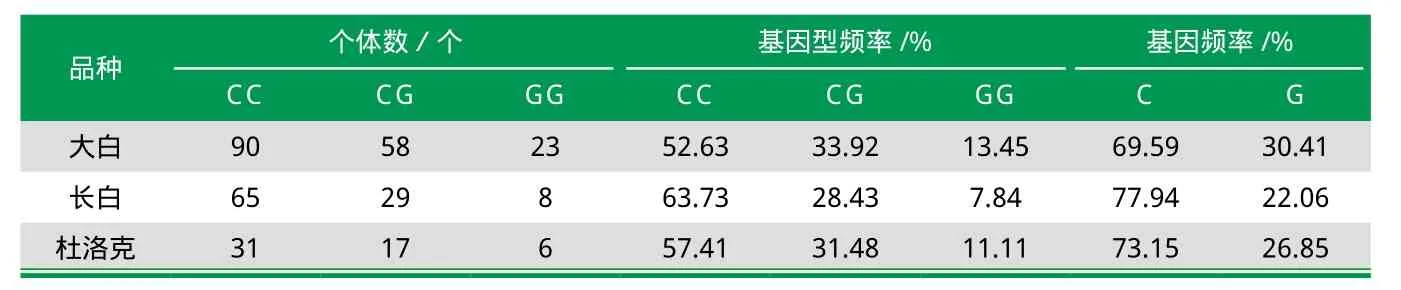
多态信息含量(PIC)是估计基因多态性的一个指标,PIC>0.5,为高度多态;0.25 目前,对DECR1基因的多态性研究主要集中在第2、第5、第6、第8外显子。Amills等[12]扩增了DECRl基因几乎全部编码序列,包括外显子2和10的937 bp片段,并通过对不同品种猪DECRl基因序列进行比对,结果发现,外显子2处存在G/C多态性,该SNP编码DECR1蛋白52号位的Val(C)-Leu(G)。李步高等[9]应用PCR-SSCP方法检测了192头山西白猪DECR1基因第5外显子位点的多态性,其表现出3种不同基因型,表明该位点存在多态性。黄生强等以大白、长白和杜洛克为样本,研究猪DECR1基因SNPs的筛选及其多态性分析时发现,第6外显子有一个SNP无义突变位点,第8外显子有一个SNP错义突变位点且样本猪群中均检测到3种基因型(如表3),表明样本猪群在该位点存在丰富的遗传多态性。 猪DECR1基因的多态性可能与生长性状、肉质性状等存在关联性,因此深入研究其遗传多态性显得极为重要。 表1 猪DECR1基因在不同猪种不同组织的表达差异 表2 诱导分化过程中DECR1基因mRNA相对表达量的最小二乘值及多重比较 表3 DECR1基因Exon8突变位点的遗传多态性 脂肪酸是肉质风味的影响因素之一,而在生物体内,饱和脂肪酸和不饱和脂肪酸之间的转变必须通过β-氧化途径完成。作为不饱和脂肪酸β-氧化途径降解中的关键限速酶,猪DECR1在线粒体内参与催化4-双烯辅酶A转变为3-单烯辅酶A的过程,使多不饱和脂肪酸获得饱和而完成氧化作用,从而调节脂肪酸的沉积。同时,DECR1基因的5个高度保守的酸残基中的1个可能是供体,它的高度表达可促进细胞内脂肪酸氧化。猪DECR1基因位于4号染色体上q1.2区域,该位置与影响猪胴体、生长及脂肪酸代谢的QTL可靠区域一致[13-14],推测其可影响脂肪酸含量、pH、肉色等肉质性状。医学研究发现其酶活性的变化或缺失对人是致命的[15]。 DECR1基因的表达受到多种因素的调控,表达量变化幅度较大,目前尚未彻底掌握猪DECR1基因的作用机制。目前研究认为猪DECR1基因的主要功能是影响脂肪沉积和肉质性状。 随着人们对猪肉品质与风味的追求,基因水平上的研究正逐渐成为寻找影响肉质因素新的突破口。目前,国内外对猪DECR1基因的研究正在不断深入,但成果性报道较少。综上所述,DECR1可能与猪脂肪沉积和肉质性状有直接关系,极有可能成为影响猪肉性状的主效基因或遗传标记,深入研究猪DECR1基因可充分挖掘其潜在的、巨大的科研和经济价值。 [1]Rosenvold K, Anderson H J.Factors of significance for pork quality-A review[J]. Met Sci,2003,64:219-237. [2]Ramirez O,Quintanilla R,Varona L,et al.DECR1 and ME1 genotypes are associated with lipid composition traits in duroc pigs[J].Animal Breeding and Genetics,2013,131:46-52. [3]Kunau W H,Dommes P.Degradation of unsaturated fatty acids.Identification of intermediates in the degradation of cis-4-decenoyl-CoA by extracts 0f beefliver mitochondria[J].Eur J Biochem,1978,9l:533-544. [4]黄生强,柳小春,蒋隽,等.猪DECRl基因SNPs筛选及其多态性分析[J].湖南农业大学学报,2011,37(2):192-194. [5]Clop A,Cercos A,Tomas A,et al.Assignment of the 2,4-dienoyl-CoA reductase(DECR)gene to porcine chromosome 4 [J].Anim Genet,2002,33:164-165. [6]Davoli R,Fontanesi L,Zambonelli P,et a1. Isolation of porcine expressed sequence tags for the construction of a first genomic transcript map of the skeletal muscle in pig[J], Anim Genet,2002,33:3-18. [7]Clop A,0vilo C,Perez-enciso M,et a1.Detection of QTL affecting fatty acid composition in the pig[J].Mamm Genome,2003,14:650-656. [8]李步高,郭晓红,曹果清,等.猪DECR1基因cDNA的克隆、序列分析及原核表达研究[J].畜牧兽医学报,2010,41(11):1371-1378. [9]李步高,郭晓红,曹果清,等.马身猪和大白猪不同组织DECR1基因的表达[J].畜牧兽医学报,2011,42(4):475-480. [10]史 利 华 ,李宝 玉 ,张 浩 ,等 .猪 DECR1基因在不同猪种及组织中的差异表达研究[J].中国畜牧杂志,2012,48(13):17-19. [11]刘海峰,阎振鑫,董涵,等.猪前脂肪细胞诱导分化过程中PPARγ、DECR1和ECHS1基因表达模式[J].四川农业大学学报,2010,28(2):215-218. [12]Amills M.Vidal O L,Varona A,et al.Polymorphism of the pig 2,4-dienoyl CoA reductase 1 gene(DECR1)and its association with carcass and meat quality traits[J].Anim Sci,2005.83:493-498. [13]Waling G A,Visscher P M,Anderssonl,et a1.Combined analyses of data from quantitative trait loci mapping studies.Chromosome 4 effects on porcine growth and fatness[J].Genetics,2000,155:1369-1378. [14]Perez-enciso M,Clop A,Noguera J L,et al.A QTL on pig chromosome 4 affects fatty acid metabolism:evidence from an Iberian by Landrace intercross[J].J Anim Sci,2000,78:2525-2531. [15]Roecr,Millington D S,Norwood D L,et a1.2,4-Dienoyl-coenzyme A reductase deficiency:a possible new disorder of fatty acid oxidation[J].J Clin Invest,1990,85(5):1703-1707.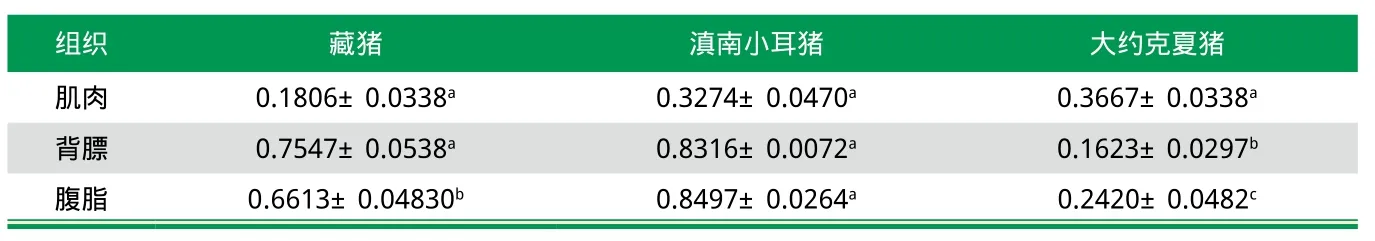
3.2 猪DECR1基因的功能
4 结语
