内生细菌EBS05对番茄病程相关蛋白的诱导作用研究
2014-09-26李俊州赵玉华文才艺
李俊州, 赵玉华, 文才艺, 臧 睿, 张 俊
(河南农业大学植物保护学院,河南 郑州 450002)
内生细菌EBS05对番茄病程相关蛋白的诱导作用研究
李俊州, 赵玉华, 文才艺, 臧 睿, 张 俊
(河南农业大学植物保护学院,河南 郑州 450002)
以番茄江蔬14为材料,研究了内生细菌EBS05对番茄病程相关蛋白基因表达的诱导作用,同时,测定了番茄体内β-1,3-葡聚糖酶和几丁质酶活性变化.结果表明,内生细菌EBS05可诱导番茄体内PR1,PR2和PR3基因持续增量表达,并有效激活PR5基因表达,进而提高番茄对番茄黄化曲叶病毒(TYLCCV)的系统抗性;内生细菌EBS05诱导处理再接种TYLCCV后,番茄体内β-1,3-葡聚糖酶和几丁质酶活性均明显高于对照,并分别于接种后第7 d和第9 d达到活性峰值.结合TYLCCV侵染番茄的病程分析,β-1,3-葡聚糖酶和几丁质酶活性的升高与内生细菌EBS05诱导番茄对TYLCCV的系统抗性呈正相关.
内生细菌;中国番茄黄化曲叶病毒;诱导抗性;病程相关蛋白
番茄黄化曲叶病毒病是由中国番茄黄化曲叶病毒(Tomato yellow leaf curl Chinavirus,TYLCCV)引起的一种番茄生产上的毁灭性病害[1],主要由烟粉虱或嫁接传播.近年来,随着全球气候变化和烟粉虱危害加重,番茄黄化曲叶病毒病在世界各地大面积暴发流行,发病地块减产严重,给番茄产业造成了严重的损失[2,3].关于番茄黄化曲叶病毒病的防治,目前生产上主要通过选育抗病品种和化学药剂控制烟粉虱等途径.然而,由于该病害的发生机理尚不明确,抗源材料缺乏,利用抗病基因工程手段获得抗病品种的研究还处在探索阶段,生产上并没有稳定有效的抗病品种[4],此外,化学防治容易导致作物药害、农药残留及昆虫抗药性等安全生产问题.因此,利用生物防治手段控制番茄黄化曲叶病毒病的研究工作渐渐引起人们的关注和重视[5,6].
生防细菌是一类具有产生抗生素、抗肿瘤类药物和蛋白酶等生物活性物质潜力的微生物资源[7].利用其作为生防因子进行植物病毒病的防治已成为生物防治的一种有效途径,如RYU等[8]利用生防细菌沙雷氏菌90-166诱导拟南芥对黄瓜花叶病毒(Cucumber mosaic virus,CMV)产生系统抗性,可明显降低拟南芥病情指数.WANG等[9]系统研究了芽胞杆菌诱导番茄对CMV的抗病作用,结果表明,芽胞杆菌对番茄植物具有明显的系统诱导抗性,能诱导番茄植物中扩张蛋白基因LeEXP2,LeEXP5和LeEXP18、调控基因NPR1以及防卫基因PR1a的增强表达.生防菌EBS05(CGMCC No.5239)是从樟树周皮组织中分离的1株内生枯草芽胞杆菌,该菌株具有较强的植物根际定殖能力,且对多种植物病原真菌和烟草花叶病毒(Tobacco mosaic virus,TMV)具有稳定的拮抗活性.前期研究结果显示,内生细菌EBS05所产生的脂肽类抗生素Surfactin A是诱导烟草对系统抗性的有效激发子[10].同时,EBS05对番茄植物具有明显的促生作用,可有效诱导番茄对TYLCCV的系统抗性,诱导抗病性效果达53.60%[11].本研究测定了EBS05诱导番茄对TYLCCV系统抗性过程中,病程相关蛋白基因的诱导表达差异,以及番茄体内β-1,3-葡聚糖酶和几丁质酶活性变化,为进一步阐明内生细菌EBS05的诱导抗性机理和新型抗病毒微生物农药的研发提供理论依据.
1 材料与方法
1.1试验材料
内生细菌EBS05由河南农业大学植物保护学
院植物病害生物防治研究室分离、保存;供试植物材料番茄(江蔬14号)和TYLCCV侵染性克隆,均由江苏农业科学院提供;总RNA提取试剂TRIzol(美国Invitrogen公司)和反转录试剂盒(TaKaRa)均购自宝生物工程(大连)有限公司.
1.2试验方法
1.2.1 接种处理 将活化的侵染性克隆接种于LB培养基,28 ℃,170 r·min-1,振荡培养16 h即为侵染性克隆接种液.接种处理时,选取长势一致的5叶期番茄幼苗,用50 mL EBS05菌悬液(108cfu·mL-1)灌根处理,24 h后,用灭菌的牙签蘸取TYLCCV侵染性克隆菌液,在番茄幼苗根基部针刺接种,处理后的番茄植株置于防虫网隔离的温室中自然生长.
1.2.2 EBS05诱导处理后番茄体内病程相关蛋白基因检测 选取长势一致的5叶期番茄幼苗,分别做如下处理:① 50 mL EBS05菌悬液灌根处理1 d后接种TYLCCV(G1);②50 mL EBS05菌悬液灌根处理1 d后用清水做接种处理(G2);③ 50 mL清水灌根处理1 d后用清水做接种处理(CK).每组处理15株,3次重复;分别于接种后1,3,5和7 d,选取番茄幼苗生长点下第3片叶,采用RT-PCR分别测定PR1,PR2,PR3和PR5基因的相对表达量.总RNA的提取参照试剂RNA TRIzolRReagent,采用反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)进行反转录,以不同处理的cDNA为模板进行PCR扩增,引物序列如表1所示;PCR反应条件为95 ℃预变性2 min;95 ℃变性30 s,56 ℃退火30 s,72 ℃ 延伸30 s,35个循环;72 ℃延伸10 min.
1.2.3 EBS05诱导处理后番茄植株内β-1,3-葡聚糖酶和几丁质酶活性的测定 选取长势一致的5叶期番茄幼苗,分别做如下处理:① 50 mL清水灌根处理1 d后接种TYLCCV(T1);② 50 mL EBS05菌悬液灌根处理1 d后用清水做接种处理(T2);③ 50 mL EBS05菌悬液灌根处理1 d后接种TYLCCV(T3);④ 50 mL清水灌根处理1 d后用清水做接种处理(CK).分别于接种处理后1,3,5,7,9,11,13和15 d,选取番茄幼苗生长点下第3片叶用于酶活性的测定,每组处理24株,3次重复.β-1,3-葡聚糖酶活性和几丁质酶活性测定方法参照文献[17,18].

表1 PCR扩增引物Table 1 Primer Sequence for PCR
2 结果与分析
2.1EBS05诱导处理后番茄体内病程相关蛋白基因的表达差异
试验结果如图1所示,内生细菌EBS05诱导处理及诱导处理后再挑战接种TYLCCV时,番茄体内PR1,PR2和PR3基因表达量均比对照显著增加,而且呈逐渐增加的趋势,并于接种处理后第7 d表达量最大.PR5基因在对照中未见表达,而EBS05诱导处理再接种TYLCCV后1 d,PR5基因被激活,并持续增量表达,而仅用EBS05诱导处理后,PR5基因从第3 d开始表达,至接种后3~7 d,PR5持续增量表达,其相对表达量低于接种TYLCCV处理.由此可见,内生细菌EBS05诱导处理后,能诱导番茄体内PR1、PR2和PR3基因的增量表达,同时,可有效激活番茄PR5基因的诱导表达,且在挑战接种TYLCCV时,其诱导表达效果更加明显.

处理1:EBS05诱导处理后接种TYLCCV;处理2:EBS05诱导处理后接种清水;对照:清水灌根处理后接种清水;M:DNA Marker;1,3,5,7分别代表接种后的天数G1:Inoculated with TYLCCV after treatment by EBS05; G2:Inoculated with water after treatment by EBS05; CK: Inoculated with water after treatment by water; M:DNA Marker;Number of 1, 3,5and 7 means days post inoculation, respectively图1 不同处理后番茄体内病程相关蛋白基因的表达Fig.1 Expression of pathogensis-related proteins genes in tomato plants after different treatments
2.2EBS05诱导处理对番茄植株内β-1,3-葡聚糖酶活性的影响
内生细菌EBS05诱导处理以及EBS05诱导处理再接种TYLCCV后,番茄体内β-1,3-葡聚糖酶活性明显升高,且呈先升高后下降的变化趋势.接种处理后1~7 d,β-1,3-葡聚糖酶活性逐渐升高,并于第7 d达到活性峰值,接种处理后7~15 d,酶活性逐渐降低,但活性值均明显高于对照健康植株.仅接种TYLCCV后,番茄体内β-1,3-葡聚糖酶活性也呈先升高后降低的变化趋势,但是酶活性明显低于EBS05诱导后再接种TYLCCV处理.番茄植株感染TYLCCV后第11 d,β-1,3-葡聚糖酶活性低于对照健康植株(图2).结合番茄黄化曲叶病毒病病程分析,番茄植株在仅接种TYLCCV后第9 d开始发病,接种后11~15 d为发病高峰,而EBS05诱导处理后,番茄黄化曲叶病毒病病害明显减轻,发病时间延迟,仅在接种后第15 d才出现顶端叶片黄化和卷曲现象.由此可见,内生细菌EBS05对番茄抗TYLCCV的诱导抗性效果与番茄体内β-1,3-葡聚糖酶活性升高有一定的相关性.

T1:清水灌根处理后接种TYLCCV;T2:EBS05诱导处理后接种清水;T3:EBS05诱导处理后接种TYLCCV;CK:清水灌根处理后接种清水.
T1: Inoculated with TYLCCV after treatment by water; T2: Inoculated with water after treatment by EBS05; T3: Inoculated with TYLCCV after treatment by EBS05; CK: Inoculated with water after treatment by water.
图2不同处理后番茄叶片内β-1,3-葡聚糖酶活性的变化
Fig.2Changesofβ-1,3-glueanasintomatoleavesafterdifferenttreatments
2.3EBS05诱导处理对番茄植株内几丁质酶活性的影响
试验结果如图3所示,对照健康番茄植物体内几丁质酶活性变化不大,而仅接种TYLCCV处理、内生细菌EBS05诱导处理和EBS05诱导处理再接种TYLCCV后,番茄植株体内几丁质酶活性均显著高于对照健康植物,且呈现先升高后下降的变化趋势,并于挑战接种处理后第9 d达到活性峰值.EBS05诱导处理和EBS05诱导处理再接种TYLCCV后9~15 d,番茄体内几丁质酶活性逐渐降低,但活性值均明显高于仅接种TYLCCV处理和对照健康植物;仅接种TYLCCV处理9 d后,几丁质酶活性显著降低,至15 d时,明显低于对照健康植株.由此可见,内生细菌EBS05诱导处理能有效提高番茄体内几丁质酶活性,进而提高番茄对TYLCCV的系统抗性.
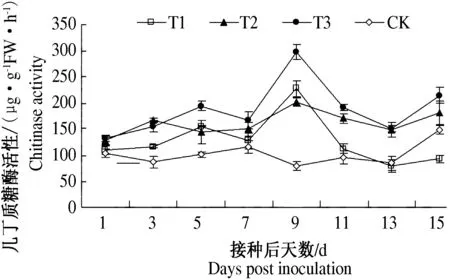
T1:清水灌根处理后接种TYLCCV;T2:EBS05诱导处理后接种清水;T3:EBS05诱导处理后接种TYLCCV;CK:清水灌根处理后接种清水.
T1: Inoculated with TYLCCV after treatment by water; T2: Inoculated with water after treatment by EBS05; T3: Inoculated with TYLCCV after treatment by EBS05; CK: Inoculated with water after treatment by water.
图3不同处理后番茄叶片内几丁质酶活性的变化
Fig.3Changesofchitinaseintomatoleavesafterdifferenttreatments
3 讨论
病程相关蛋白(Pathogensis-related proteins, PRPs)是指植物在受到病原物侵染或非生物因子诱导协迫条件下产生一类与植物抗性密切相关的水溶性蛋白.一般认为,PRPs在植物病害系统中被诱导表达,并参与植物对病原物的防卫反应[19],如苏杭等[20]用丁香酚处理烟草后,诱导烟草体内病程相关蛋白基因PR1,PR3和PR5表达,明显提高了植株对TMV的抗病性.也有研究表明,PRPs在植物体内为组成型表达,以维持植物对病原物的自然抗病性[21].从本研究结果来看,未经接种处理的番茄体内均可检测出PR1,PR2和PR3基因的表达,内生细菌EBS05诱导处理后,其表达量持续显著增加,而PR5基因在未经接种处理的番茄体内未见表达,内生细菌EBS05诱导处理后,PR5基因被激活,并持续增量表达.由此可见,病程相关蛋白PR1,PR2和PR3在番茄体内存在组成型表达,而PR5则属于诱导性表达.PR5蛋白又称类甜蛋白(Thaumatin-like proteins, TLPs),是植物自然免疫系统的组成之一[22].近年来,PR5蛋白因具有抗真菌活性和激活植物防卫反应信号途径而倍受关注[23].现已在水稻、烟草、葡萄等多种植物真菌病害系统中被证实存在PR5蛋白的诱导性表达,并参与植物的防卫反应[24~26],但尚未见PR5蛋白在番茄病毒病害系统中诱导表达,并诱导番茄对病毒侵染产生系统抗性的报道.本研究结果表明,内生细菌EBS05可有效诱导番茄体内PR5基因的表达,尤其是挑战接种TYLCCV后,诱导表达效果更加明显.该研究结果对基于转PR5基因的番茄抗TYLCCV基因工程育种策略具有重要的参考价值.
目前,已鉴定的PRPs包括17个蛋白家族,其中,11个PRPs家族来自烟草和番茄植物[20].鉴于PRPs与植物抗病性密切相关,有关PRPs诱导合成的信号转导途径及其在植物防卫反应中的功能已成为植物诱导抗性机理研究的重要内容,其中,以PR1,PR2和PR3研究最为深入.在PRs家族中,PR1分布最广泛、含量最高,约占植物叶片总蛋白的1%~2%,是植物防卫反应中最重要的功能蛋白之一,但目前对其功能尚不明确;PR2和PR3分别具有β-1,3-葡聚糖酶和几丁质酶活性,在植物抗真菌病害的防卫反应中发挥重要作用,因此,基于上述3类病程相关蛋白基因的转基因策略被广泛应用于植物抗病基因工程育种实践,测定β-1,3-葡聚糖酶和几丁质酶活性也成为研究植物诱导抗病性效果的重要手段.本研究结果表明,内生细菌EBS05诱导处理后1~7 d,β-1,3-葡聚糖酶和几丁质酶活性均显著高于对照,且呈逐渐升高的趋势,该结果与PR2和PR3基因在mRNA水平上的检测结果一致.EBS05诱导处理能诱导番茄PR1,PR2和PR3基因增量表达,进而增加PR1,PR2和PR3蛋白的积累,在防卫反应中,表现为β-1,3-葡聚糖酶和几丁质酶活性的持续升高.此外,菌株EBS05诱导处理番茄植物后,TYLCCV侵染症状明显减轻,且发病时间延迟,由此可见,β-1,3-葡聚糖酶和几丁质酶活性与番茄对TYLCCV的诱导系统抗性呈正相关,此研究结果与文献的研究[27~29]结果一致.
[1] 刘剑峰,肖启明,张德咏,等. 番茄黄化曲叶病(TYLCV)的研究进展[J]. 中国农学通报,2013,29(13):70-76.
[2] PICO B, DIEZ M J, NUEZ F. Viral diseases causing the greatest economic losses to the tomato cropⅡ. The tomato yellow leaf curl virus: a review [J]. Scientia Horticulturae, 1996, 67(3-4): 151-196.
[3] 宋建军,刘红宵,仇 燕,等. 番茄黄化曲叶病毒病的发生分布及防治对策[J]. 北方园艺,2010(7):147-150.
[4] 孙书娥,唐前君,刘 勇,等. RNA沉默介导的转基因烟草抗中国番茄黄化曲叶病毒研究[J]. 农业生物技术学报,2013,21(5):530-536.
[5] 邢卫锋,丁雪玲,柯红娇,等. 番茄黄化曲叶病毒病生防菌的筛选及防治效果研究[J]. 江苏农业科学, 2013, 41(9):110-112.
[6] 王春梅. 丁子香酚和接种番茄黄化曲叶病毒对番茄几丁质酶和β-1,3-葡聚糖酶活性的影响[J]. 江西农业学报,2013,25(9):78-80.
[7] STROBEL G A, DASISY B, CASTILLO U, et al. Natural products from endophytic microorganisms[J]. Journal of Natural Products, 2004, 67(2):257-268.
[8] RYU C M, MUROHY J F, MYSORE K S, et al. Plant growth-promoting rhizobacteria systemically protectArabidopsisthalianaagainst cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway [J]. The Plant Journal, 2004, 39(3):381-382.
[9] WANG S, WU H J, ZHAN J, et al. The role of synergistic action and molecular mechanism in the effect of genetically engineered strainBacillussubtilisOKBHF in enhancing tomato growth and cucumber mosaic virus resistance[J]. Biological Control, 2011,56(1):113-121.
[10] 周 蕊. 内生细菌EBS05诱导烟草抗TMV机理的研究[D]. 郑州:河南农业大学,2013.
[11] 李俊州,王春梅,文才艺,等. 内生细菌EBS05对番茄抗中国黄化曲叶病毒的诱导作用[J]. 江苏农业学报,2014,30(4):746-751.
[12] 唐平华,陈国平,潘 宇,等. 番茄多蛋白桥梁因子基因LeMBF1的克隆及抗病性分析. 2012, 16(2):138-143.
[13] KAN J L, JOOSTEN M H, WAGEMAKERS C A,et al. Differential accumulation of mRNAs encoding extracellular and intracellular PR proteins in tomato induced by virulent and avirulent races ofCladosporiumfulvum[J]. Plant Molecular Biology, 1992, 20(3):513-527.
[14] DANHASH N, WAGEMAKERS C A, KAN J A, et al. Molecular characterization of four chitinase cDNAs obtained fromCladosporiumfulvum-infected tomato [J]. Plant Molecular Biology, 1993, 22(6):1017-1029.
[15] REP M, DEKKERr H L, VOSSEN J H,et al. Mass spectrometric identification of isoforms of PR proteins in xylem sap of fungus-infected tomato [J]. Plant Physiology, 2002, 130(2): 904-917.
[16] 李 仁,吴 新,李 蔚,等. 番茄水通道蛋白基因SlAQP的克隆与序列分析[J]. 中国农业科学, 2012,45(2):302-310.
[17] 曲明静,黄忠林,鞠 倩,等. 花生网斑病原菌和黄曲霉菌对对花生几丁质酶诱导研究[J]. 花生学报,2008,37(1):1-6.
[18] 孙 斌,李多川,慈晓艳,等. 小麦叶片β-1,3-葡聚糖酶的诱导、纯化与抗菌活性[J]. 植物生理与分子生物学报,2004,30(4):399-404.
[19] SEBASTIEN A,CHRISTELLE C, CLAUDE A, et al. Comparative analysis of PR gene expression in tomato inoculated with virulentFusariumoxysporumf. sp.lycopersiciand the biocontrol strainF.oxysporumFo47 [J]. Physiological and Molecular Plant Pathology, 2008, 73(1-3):9-15.
[20] 苏 杭,王春梅,陈 浩,等. 丁香酚对烟草抗烟草花叶病毒的诱导作用初探[J]. 农药学学报, 2012,14(1):24-29.
[21] EDREVA A. Pathogenesis-related proteins: research progress in the last 15 years [J]. General and Applied Plant Physiology, 2005, 31(1-2):105-124.
[22] PARK C J, AN J, SHIN Y C, et al. Molecular characterization of pepper germin-like protein as the novel PR-16 family of pathogenesis-related proteins isolated during the resistance response to viral and bacterial infection [J]. Planta, 2004, 219:797-806.
[23] EI-KEREAMY A. EI-SHARKAWY I, RAMAMOORTHY R, et al.Prunusdomesticapathogenesis-related protein-5 activates the defense response pathway and enhances the resistance to fungal infection [J]. PLoS ONE, 2011, 6(3): e17973.
[24] DATTA K, VELAZHAHAN R, OLIVA N, et al. Over-expression of the cloned rice thaumatin-like protein (PR-5) gene in transgenic rice plants enhances environmental friendly resistance toRhizoctoniasolanicausing sheath blight disease [J]. Theoretical and Applied Genetics, 1999, 98(6-7):1138-1145.
[25] ABADA L R, URZOA M P, LIU D, et al. Antifungal activity of tobacco osmotin has specificity and involves plasma membrane permeabilization [J]. Plant Science, 1996, 118:11-23.
[26] SALZMAN R A, TIKHONOVA I, BORDELON B P, et al. Coordinate accumulation of antifungal proteins and hexoses constitutes a developmentally controlled defense response during fruit ripening in grape[J]. Plant Physiology,1998,117:465-472.
[27] 左豫虎,康振生,杨传平,等. β-1,3-葡聚糖酶和几丁质酶活性与大豆对疫霉根腐病抗性的关系[J]. 植物病理学报,2009,39(6):600-607.
[28] 吴艳兵,谢荔岩,谢联辉,等. 毛头鬼伞多糖对烟草酶活性和同工酶谱的影响[J]. 微生物学杂志,2007,27(5):29-33.
[29] 黑银秀,朱为民,郭世荣,等. 核黄素和接种番茄黄化曲叶病毒对番茄几丁质酶和β-1,3-葡聚糖酶活性的影响[J]. 南京农业大学学报,2012,35(4):135-139.
(责任编辑:朱秀英)
Pathogensis-relatedproteinsinducedbyendophyticbacteriaEBS05intomatoplant
LI Jun-zhou, ZHAO Yu-hua, WEN Cai-yi, ZANG Rui, ZHANG Jun
(College of Plant Protection, Henan Agricultural University, Zhengzhou 450002, China)
Genes expression of pathogensis-related proteins(PRPs) induced by endophytic bacteria EBS05 in tomato plant were investigated using var. Jiangshu 14 as test material, and the activities of β-1, 3-glueanas and chitinase were determined too. The results showed that strain EBS05 could induce the overexpression ofPR1,PR2 andPR3 genes, and activate the genePR5 to high level expression in tomato plant, thus increasing the systemic resistance to Tomato Yellow Leaf Curl Chinavirus(TYLCCV). The activities of β-1, 3-glueanas and chitinase in tomato plants inoculated with TYLCCV after pretreated by strain EBS05 were higher than those in the control plants, and the activity peak appeared at the 9thand 7thdays post inoculation, respectively. The increase of β-1, 3-glueanas and chitinase activities were positively correlated with the systemic resistance to TYLCCV in tomato plants mediated by strain EBS05 based on the pathogensis of TYLCCV-infected tomato plants.
endophytic bacteria; tomato yellow leaf curl China virus; induced resistance; pthogensis-related proteins
S 432.1
:A
2014-03-26
国家自然科学基金项目(31272083);河南省教育厅科学技术研究重点项目(13A210473)
李俊州,1990年生,男,河南柘城人,硕士研究生,从事植物病害生物防治研究工作.
文才艺,1965年生,男,湖北孝感人,教授,博士.
1000-2340(2014)05-0602-06
