百山祖两种阔叶木本植物的光合蒸腾作用特性研究
2014-09-21吴雁雯张金池顾哲衍
吴雁雯, 张金池, 顾哲衍, 刘 鑫, 韩 诚
(南京林业大学, 南京 210037)
百山祖两种阔叶木本植物的光合蒸腾作用特性研究
吴雁雯, 张金池, 顾哲衍, 刘 鑫, 韩 诚
(南京林业大学, 南京 210037)
利用Li-6400便携式光合测定仪分别对凤阳山—百山祖国家级自然保护区阔叶混交林中2种常见木本植物尖萼紫茎(Stewartiaacutisepala)和小叶白辛树(Pterostyraxcorymbosus)的光合蒸腾特性进行了研究,并应用相关性分析与逐步回归分析法就上述植物各生理生态因子对净光合速率(Pn)和蒸腾速率(Tr)的影响进行了分析。结果表明:(1) 尖萼紫茎的Pn主要影响因子是水分利用率(WUE)、太阳辐射(PYR),而小叶白辛树的主要影响因子则是胞间CO2摩尔分数(Ci),PYR,且在Pn日变化过程中,尖萼紫茎的值始终大于小叶白辛树,可以判断尖萼紫茎的光合作用比小叶白辛树要强;(2) 影响尖萼紫茎Tr的主要因子是气孔导度(Gs)及光合有效辐射(PAR),影响小叶白辛树的主要因子则是Gs、空气相对湿度(RH),而据Tr日变化可发现尖萼紫茎的蒸腾作用强于小叶白辛树;(3) 对2种植物WUE的分析可知,尖萼紫茎的碳水平衡性优于小叶白辛树。且总体而言,在该生境中,制约尖萼紫茎光合蒸腾作用的因子比小叶白辛树少。因此,尖萼紫茎对该生境适应性优于小叶白辛树,分布更为广泛。
百山祖; 净光合速率; 蒸腾速率; 生理因子; 生态因子
森林生态系统水源涵养功能与生态效益评价都与其光合—蒸腾作用的准确估算密切相关[1]。由于森林生态系统由不同等级水平的系统组成,而从林木个体到森林生态系统之间具有不同的尺度特征,因此尺度外推是完成从林木个体水平光合蒸腾作用观测到森林生态系统能量流动推算的重要手段。林分的树种组成多样性以及不同树种的林木个体光合蒸腾特性的差异成为近年研究的重点[2-4]。生理生态因子诸如气孔导度、胞间CO2浓度、气孔限制值、有效辐射、温度、相对湿度等对光合蒸腾作用的影响不是单一孤立的,而是相互联系、相互制约的[5]。因此不同地区、不同植物的光合蒸腾作用日变化特征各异,且影响不同植物日变化的主要因子也不尽相同[6]。
浙江省凤阳山—百山祖国家级自然保护区由福建省武夷山脉向东伸展而成,受到亚热带湿润季风气候影响[7]。林地内树冠繁茂,种类繁多,其中阔叶混交林是我国最具代表性的生物群落之一[8]。对该林分类型植物光合蒸腾特性及相关环境因子的影响进行探究,不仅能对植物生长状况的好坏及对生境适应性的强弱进行合理判断[9],达到为林分优化工作提供改良思路的目的,亦对我国其它地区同类研究具有重要的借鉴和参考意义。尖萼紫茎(Stewartiaacutisepala)、小叶白辛树(Pterostyraxcorymbosus)是凤阳山阔叶混交林中较常见的2种木本植物,多生存于林冠下层。通过对相同生境条件下2个树种光合蒸腾特性以及相关生理因子的测定,比较该2种植物在凤阳山地区的适应能力,对其环境适应性与水土保持功能进行评估,以期为亚热带阔叶混交林的生态效益评估以及生态系统恢复与管理提供一定的理论依据。
1 研究区概况
凤阳山自然保护区位于浙西南丽水市,是浙江凤阳山—百山祖国家级自然保护区的一部分,地处北纬27°46′—27°58′、东经119°06′—119°15′。保护区位于东南沿海,属亚热带湿润季风气候,最高气温30.2℃,极端最低气温-11.0℃,年均气温12.3℃,年降水量2 438.0 mm,年蒸发量1 171.0 mm。海拔一般为300~1 500 m,其主峰黄茅尖海拔高达1 929 m。凤阳山土壤的分布可划分为:红壤、黄壤、山地草甸土及粗骨土。其中黄壤土类是凤阳山的主要土壤类型,多分布于800 m以上的高海拔山坡地。红壤土类属地带性土壤,分布于本区海拔800 m以下山坡。保护区内物种多样,生物资源丰富,主要植被类型包括针叶林、针阔叶混交林、常绿阔叶混交林、常绿阔叶林等[10]。
2 材料与方法
在对保护区内阔叶混交林进行调查的基础上,选择该林分中坡向、坡度、坡位相近的2种主要阔叶乔木进行研究,样地基本情况如表1所示。在阔叶混交林内通过每木检尺选择标准木作为测定样本,并于待测样本树上选择生长良好且叶面朝向较一致的顶端成熟叶,以保证所测叶片受光良好。因植株高大,且经测定2个树种离体叶片在1 h内均有较高、较稳定的水分供应和光合活力,叶片光合—蒸腾日变化采用离体测定,将离体枝条或复叶插入水中,环剥去除切口处3 cm左右的韧皮部和剩余叶片进行测定。应用Li-6400便携式光合仪,测定了包括蒸腾速率(Tr)、胞间CO2摩尔分数(Ci)、净光合速率(Pn)、气孔导度(Gs)和叶片水汽压亏缺(VPD)等在内的相关生理因子参数,同时测定了气温(Ta)、太阳辐射(PYR)、光合有效辐射(PAR)、空气相对湿度(RH)等生态因子参数。

表1 样地基本特征
日变化测定选择于2013年7月中旬的1个典型晴朗天气,测定时间为8:00—18:00,间隔2 h测定1次,每次测定10棵树作为重复,每棵树进行3个叶片样品处理,取平均值进行分析。
气孔限制值计算公式[11]:
式中:Ls——气孔限制值(%);Ca——大气CO2摩尔分数;Ci——胞间CO2摩尔分数;J——CO2补偿点,可忽略不计。
水分利用率计算公式[11]:
式中:WUE——水分利用率(μmol/mmol)
所得数据的初步分析与整理通过Excel完成,相关性分析及逐步回归分析等统计由SPSS 19.0软件完成。
3 结果与分析
3.1 光合蒸腾作用日变化
3.1.1 净光合速率(Pn)与相关因子日变化 自然条件下,尖萼紫茎和小叶白辛树在8:00—18:00观测时段内的净光合速率(Pn)日变化曲线都呈“单峰型”(图1),且变化趋势相近。2树种的Pn都在10:00达到最大值,分别为3.45 μmol/(m2·s),3.33 μmol/(m2·s),随后呈不同幅度的下降,尖萼紫茎的下降趋势较为缓慢,最终降至0.24 μmol/(m2·s),而小叶白辛树降幅稍大,在18:00时降至负值[-0.32 μmol/(m2·s)]。在一天的日变化过程中,两种树木Pn相对其他树种较低,可推断与林下低光强有较大关系。同时,尖萼紫茎的Pn值始终高于小叶白辛树,可以证明尖萼紫茎单位面积可积累的干物质量高于小叶白辛树,在14:00—16:00间小叶白辛树从积累干物质转变为消耗干物质,可能由于受气孔限制或该树种利用弱光能力较差[12],导致其在该时段内的呼吸作用大于实际光合作用。

图1 净光合速率(Pn)日变化
当前对植物光合日变化的研究已深入到气孔限制和非气孔限制的水平,对植物羧化效率、RUBP再生能力、气孔限制值为0时的光合速率等研究也有较
大的进展[13]。植物叶片的胞间CO2摩尔分数(Ci)是外界CO2进入叶片叶肉细胞所受阻力与动力及叶片内光合与呼吸作用后达到的最终平衡[14],因此理论上说,假设植物叶片同化CO2的能力保持不变,那么中午气孔导度(Gs)的下降必然限制植物体内CO2的扩散,从而导致Ci的下降。而根据近年来的相关研究可以发现[15-16],午间植物Pn下降的同时Gs虽然也随之下降,Ci却基本保持不变,有时还会上升。经大量研究,有学者提出[17],只有当Pn与Ci的变化方向相同,且Ls与两者方向相反时,方可认为Pn的变化主要受气孔因子影响。例如,当Pn,Ci两者同时减小,而Ls增大,此时可推断导致Pn的下降主要因素是气孔。否则,若Pn下降,Ci上升,同时Ls下降,可断定Pn的下降主要由叶肉细胞利用CO2能力降低引起[18],即非气孔因素造成。
由图2可知,在14:00—18:00间,尖萼紫茎与小叶白辛树Ci的值随Pn的下降而上升,且Ls减小,可见此时Pn下降的主要因素是叶肉细胞的同化能力下降。结合样地的温度日变化可以判断,此时以上2树种Pn的降低的主要原因都是由于受到午后环境高温与高光照的影响[19],致使叶肉细胞中参与羧化作用的酶活性降低,从而导致Pn的下降。
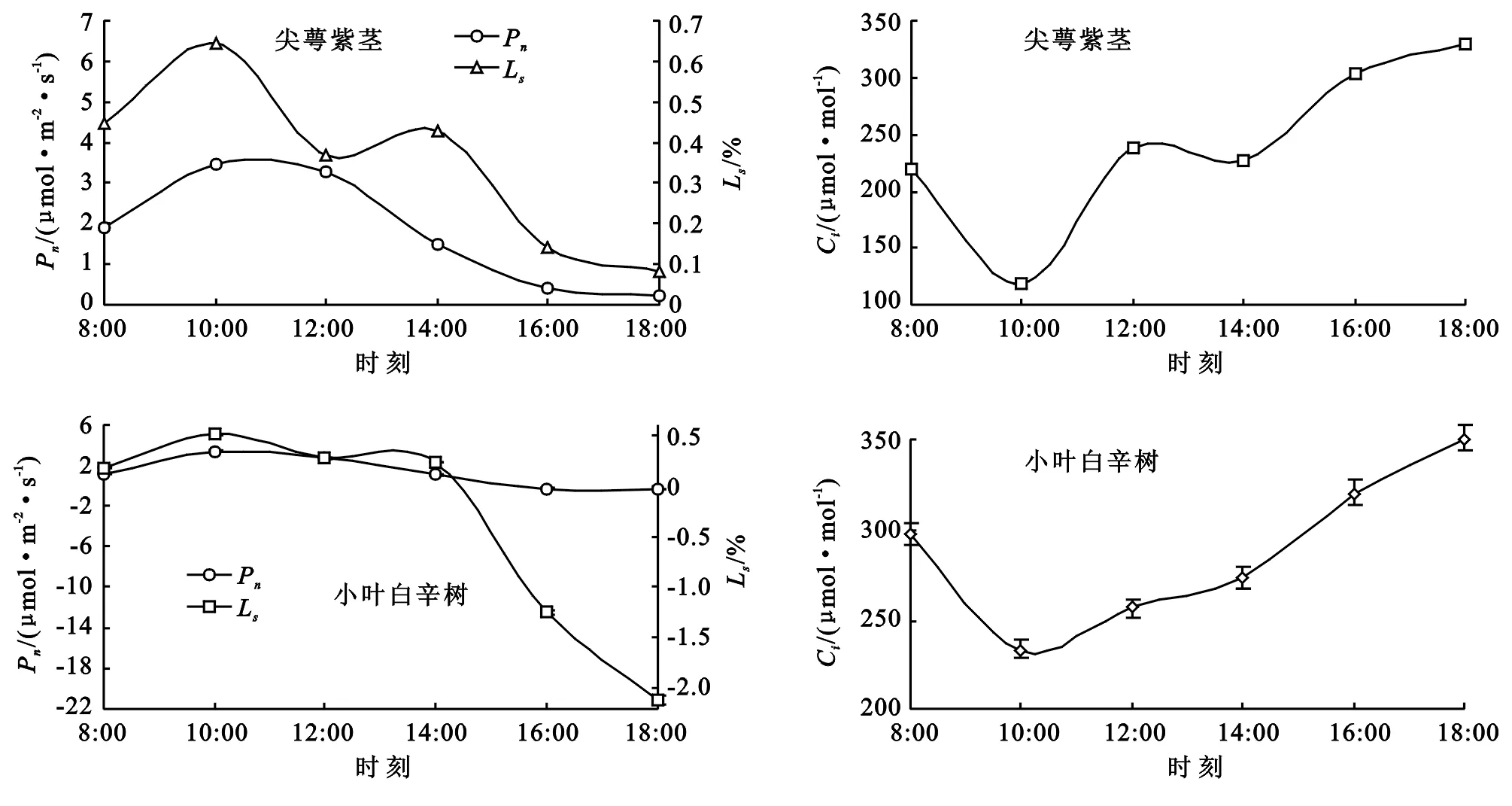
图2 净光合速率(Pn)、胞间CO2摩尔分数(Ci)和气孔限制值(Ls)日变化
3.1.2 蒸腾速率(Tr)与相关因子日变化 据图3所示,在8:00—10:00间,很可能因为环境温度突然有较大幅度的升高,小叶白辛树的气孔发生应激性的暂时关闭[20],导致其Tr从0.85 mmol/(m2·s)降至0.66 mmol/(m2·s),但随后植物自身的调节机制使其恢复常态。小叶白辛树在此处为林冠较下层木,有研究表明[21],林冠上层的叶片受到强光照射,气孔导度下降后恢复需要较长时间,处于林冠下层的植物叶片则可以保持相对较高的气孔导度,即使下降也可较快恢复。尖萼紫茎的Tr在这一时段则呈平稳上升趋势,随后尖萼紫茎与小叶白辛树的Tr不同程度上升,在12:00分别达到最大值1.12 mmol/(m2·s),1.30 mmol/(m2·s)。随后2树种Tr呈不同幅度下降趋势,在16:00—18:00间,小叶白辛树下降较为显著,从0.27 mmol/(m2·s)降至0.17 mmol/(m2·s),而尖萼紫茎则趋于平缓,稳定在0.58 mmol/(m2·s)左右。

图3 蒸腾速率(Tr)日变化
气孔作为叶片与外界进行气体交换的最重要通道,可根据外界环境变化自我调节开度大小[22],合理平衡水分散失与CO2获取两者间的关系,从而以较少的失水获得生理活动所需CO2的量。气孔导度(Gs)是气孔对这一过程的调控能力的量化指标,该指标反映了植物叶片气孔传导气体的能力大小。气孔的开闭程度对植物叶片水分散失具有直接影响[23]。叶片水汽压亏缺(VPD)是在同温度下饱和汽压与实际蒸汽压之间的差值[24],是影响植物叶片蒸腾作用的主要因素之一。因此,Tr的大小在很大程度上取决于Gs和VPD两个因子。尖萼紫茎和小叶白辛树Tr,Gs及VPD的日变化值如表2所示,可见两种植物Tr与Gs的日变化趋势接近,而与VPD的日变化趋势则并不相近。在下文的相关性分析中(表3),Tr与Gs呈极显著正相关而与VPD无显著相关性,因此,Gs是Tr的日变化进程的主要影响因子。

表2 蒸腾速率(Tr)、气孔导度(Gs)和水汽压亏缺(VPD)日变化
3.1.3 水分利用效率(WUE)日变化 尖萼紫茎的WUE相较于小叶白辛树变化相对平缓(图4),其高峰在10:00(4.39 μmol/mmol)出现,随后缓慢下降至0.41 μmol/mmol。小叶白辛树的WUE于8:00—12:00间的变化趋势与尖萼紫茎相近,最大值出现于10:00,为5.02 μmol/mmol,14:00后呈现较明显的下降趋势。水分利用效率指单位水量通过叶片蒸腾散失时光合作用所形成的有机物量[25],实际反映了植物消耗水量与干物质产量间关系,是评价植物生长对环境适应性的综合指标。由此证明,在尖萼紫茎在相同条件下积累等量干物质所需消耗水的量较少,即其积累干物质与耗水量之间的平衡比小叶白辛树更易达到,对生境的适应性也较好。
尖萼紫茎和小叶白辛树2种植物的WUE和Ls的日变化都呈现较显著的线性关系(图5),R2分别为0.959 4,0.760 3,且下文表3中2种植物WUE与其Pn,Tr的相关性和Ls与Pn,Tr的相关性显著度非常相似。可以判断,虽然在实际生理活动过程中,各个因子之间存在着复杂的互相作用与反馈机制[26],但Ls是影响研究中植物叶片WUE变化的重要因素。导致这种现象的原因是Ls的变化会造成Pn,Tr的显著变化,而这两个因子对植物的WUE产生直接影响。
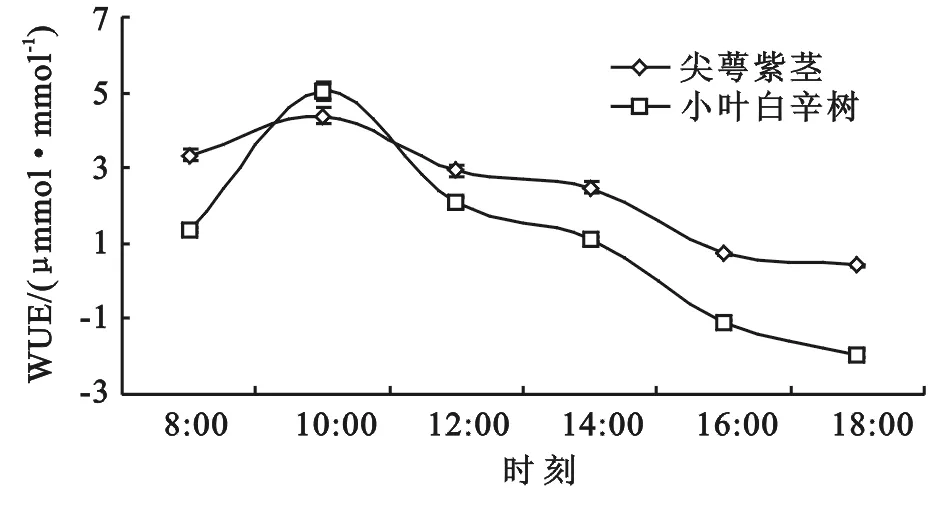
图4 水分利用效率(WUE)日变化
3.2光合蒸腾作用与相关生理生态因子的关系
3.2.1 Pearson相关性分析 如表3所示,通过对尖萼紫茎与小叶白辛树的Pn及各生理因子的Pearson相关性分析可以发现,尖萼紫茎的Pn与Ci呈显著负相关,与WUE则呈显著正相关;小叶白辛树的Pn与Ci呈极显著负相关,与Ls及WUE两个因子呈极显著正相关,与Gs显著正相关。
对各生态因子的Pearson相关性分析中可知,尖萼紫茎的Pn只与PAR呈显著正相关;小叶白辛树的Pn与RH显著负相关,与PAR显著正相关。

图5 水分利用效率(WUE)与气孔限制值(Ls)的关系
据不同树种Tr与各生理生态因子的Pearson相关分析,尖萼紫茎的Tr与Gs,PAR极显著正相关;而与小叶白辛树Tr显著正相关的因子有Ls,WUE和PAR,与其显著负相关的因子是Ci与RH。

表3 光合蒸腾速率与生理生态因子相关性
注:**表示相关显著性水平为0.01;*表示相关显著性水平为0.05;-表示呈负相关。
陆地生态系统的碳水循环间存在耦合性[27],因此,作为陆地生态系统碳循环中最为重要的生物学过程,一般来说,植物光合作用与作为陆地生态系统水循环重要环节之一的蒸腾作用之间也应紧密相关[28]。据表4所示,小叶白辛树光合与蒸腾显著相关,而尖萼紫茎的光合蒸腾作用间无显著相关性,根据对各生理生态因子的相关性分析可以发现,Ci,Ls以及WUE三个因子同时与小叶白辛树Pn,Tr显著相关,而尖萼紫茎的Pn,Tr的主要相关因子并无相同。
由于Ls的变化受Ci影响较大,而WUE则是Pn除以Tr所得商,所以可初步推断小叶白辛树的光合蒸腾作用同时受到气孔因子的调控。尖萼紫茎的Tr显然受气孔影响较大(表3),气孔对其Pn限制则不明显。

表4 净光合速率(Pn)与蒸腾速率(Tr)相关性
注:*表示相关显著性水平为0.05。
3.2.2 逐步回归分析 Pearson相关性分析只是对单因子进行线性分析,而逐步回归分析则是整体地分析各自变量的影响程度,并决定具有代表性的重要影响因子及其对因变量的解释能力[29]。

表5 光合蒸腾特性相关参数的逐步回归分析
式中:Y1为尖萼紫茎Pn;Y2为尖萼紫茎Tr;X1为尖萼紫茎WUE;X2为PAR;X3为尖萼紫茎Gs;X4为PAR;Y3为小叶白辛树Pn;Y4为小叶白辛树Tr;X5为小叶白辛树Ci;X6为PAR;X7为小叶白辛树Gs;X8为RH。
根据逐步回归方程可以看出(表5),就尖萼紫茎而言,各生理生态因子中对其Pn影响最大的生理因子与生态因子分别为WUE,PYR;而对小叶白辛树的Pn影响最大的生理因子为Ci,影响最大的生态因子为PYR。同时,尖萼紫茎的Tr变化分别主要受生理因子Gs以及生态因子PAR的调控;小叶白辛树Tr的主要影响因子则分别为Gs,RH。可以看出,逐步回归分析与Pearson相关性分析的结果不尽相同,尤以影响小叶白辛树蒸腾作用的主要生理生态因子差异最为明显。
4 讨论与结论
一般认为[30],植物协调碳同化与水分耗散之间关系的能力在很大程度上决定了该植物对当地生态环境条件适应性的强弱,因此植物的光合蒸腾作用成为衡量该植物是否适应其生境的重要指标。
(1) 李铭怡[31],肖文发[32]等人的研究表明,辐射强度、空气湿度等环境因子对植物的光合作用日变化进程是否出现“光合午休”现象具有显著影响。在对尖萼紫茎与小叶白辛树光合作用日变化的分析中,2树种的净光合速率变化曲线均呈单峰值,并未出现“光合午休”现象,因此在一定程度上说明以上2种植物对当地环境的应不存在极为不适应的情况[33]。尖萼紫茎与小叶白辛树净光合速率日均值分别为1.796 2 μmol/(m2·s),1.272 0 μmol/(m2·s),且小叶白辛树的净光合速率在一天的变化过程中始终小于尖萼紫茎,并降至负值,说明尖萼紫茎对光能的利用能力要明显强于小叶白辛树。
(2) 尖萼紫茎与小叶白辛树的蒸腾速率日均值分别为0.706 1 mmol/(m2·s),0.698 2 mmol/(m2·s),可见尖萼紫茎的蒸腾速率大于小叶白辛树。水分利用效率是反映植物光合蒸腾特性的一项综合指标[34],2种植物水分利用效率的日均值大小顺序为:尖萼紫茎>小叶白辛树。说明综合而言,尖萼紫茎的碳水平衡能力优于小叶白辛树。
(3) 根据Pearson相关分析,尖萼紫茎净光合速率的主要相关因子是胞间CO2摩尔分数、水分利用效率、气孔限制值及光合有效辐射;小叶白辛树净光合速率的主要相关因子则包括了胞间CO2摩尔分数、气孔限制值、水分利用效率、气孔导度、相对湿度和光合有效辐射。同时,尖萼紫茎蒸腾速率的主要相关因子气孔导度与光合有效辐射,小叶白辛树蒸腾速率的主要相关因子则有胞间CO2摩尔分数、气孔限制值、水分利用效率、空气相对湿度与光合有效辐射5个。可见,制约小叶白辛树的蒸腾速率的因子明显多于尖萼紫茎。逐步回归分析得出的结果则是:对尖萼紫茎净光合速率贡献最大的生理因子为水分利用效率,贡献最大的生态因子则是光合有效辐射;对其蒸腾速率贡献最大的生理因子为气孔导度,贡献最大的生态因子则是光合有效辐射;对小叶白辛树净光合速率贡献最大的生理因子是胞间CO2摩尔分数,贡献最大的生态因子是光合有效辐射;对其蒸腾速率贡献最大的生理因子是气孔导度,贡献最大的生态因子为相对湿度。
尖萼紫茎的光合作用主要受到水分与光照的调控,此处水分利用率的变化主要是由气孔的开闭所导致;蒸腾作用则受气孔和光强调控。小叶白辛树的光合蒸腾作用受到多个因子制约,影响其光合作用的最主要因子是气孔和光照;制约其蒸腾作用的最重要因素是气孔与相对湿度,结合光合有效辐射的变化规律,可以判断是由于光照强度的变化,引致环境中相对湿度大小的改变,最终导致蒸腾速率的变化。本文研究结果显示,尖萼紫茎对百山祖生境的适应性优于小叶白辛树,结合前期对该生境物种组成和群落结构的调查统计[35],可判断尖萼紫茎属百山祖常绿阔叶林的优势树种。
[1] 周国逸.生态系统水热原理及其应用[M].北京:气象出版社,1997.
[2] 雷声坤,王玉杰,王云琦,等.缙云山三种典型阔叶树光合生理特性研究[J].水土保持研究,2012,19(2):189-194,200.
[3] 丁晓纲,何茜,李吉跃,等.毛乌素沙地樟子松和油松人工林光合生理特性[J].水土保持研究,2011,18(1):215-219.
[4] 李召青,张卫强,殷祚云,等.东江中上游4树种水势日变化特征及其与环境因子的关系[J].水土保持研究,2011,18(1):243-247,267.
[5] Habib ur Rahman Khan, Link W, Hocking T J, et al. Evaluation of physiological traits for improving drought tolerance in faba bean(ViciafabaL.)[J]. Plant and Soil,2007,292(1):205-217.
[6] 马玲,赵平,饶兴权,等.乔木蒸腾作用的主要测定方法[J].生态学杂志,2005,24(1):88-96.
[7] 王伟,骆争荣,周荣飞,等.百山祖常绿阔叶林木本植物的生境相关性分析[J].生物多样性,2011,19(2):134-142.
[8] 王群,张金池,田月亮,等.浙江凤阳山天然混交林林分空间结构分析[J].浙江农林大学学报,2012,29(6):875-882.
[9] 赵平,刘惠,孙谷畴.4种植物气孔对水汽压亏缺敏感度的种间差异[J].中山大学学报:自然科学版,2007,46(4):63-68.
[10] 田月亮.凤阳山主要林分类型结构特征及其改土效应[D].南京:南京林业大学,2012.
[11] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[12] 余彦波,刘桐华.植物光效生态研究:Ⅰ.小麦光合午休的原因[J].生态学报,1985,6(4):336-342.
[13] 高辉远,邹琦,程炳嵩.大豆光合日变化过程中气孔限制和非气孔限制的研究[J].西北植物学报,1993,13(2):96-102.
[14] 向仰州,姚斌,尚鹤,等.五氯酚胁迫对转基因杨树光合光响应特性的影响[J].生态环境学报,2009,18(6):2146-2150.
[15] Kuppers M, Wheeler A M, Kuppers B I L, et al. Carbon fixation in eucalyptus in the field:Analysis of diurnal variation in Photosynthetic capacity[J]. Oecologia,1986,70(2):273-282.
[16] Rasehke K, Resemann A. The midday depression of CO2assirnilation in leaves ofArbutusunedoL. diurnal change in photosynthetic capacity related to changes in temperature and humidity[J]. Planta,1986,168(4):546-558.
[17] Bradford K J, Sharkey T D, Farquhar G D. Gas exchange, stomatal behavior, and deltaCvalues of the flacca tomato mutant in relation to abscisic acid[J]. Plant Physiology,1983,72(1):245-50.
[18] 郭春芳,孙云,张木清.土壤水分胁迫对茶树光合作用—光响应特性的影响[J].中国生态农业学报.2008(6):1413-1418.
[19] Kramer P, Kozlowslit T. Physiology of woody plants[M]. NewYork:Academic Press,1979.
[20] 李倩,谭雪莲.旱地植物蒸腾作用研究进展[J].甘肃农业科技,2006(10):18-20.
[21] Michael T, Charles R W, Mark A A. Dynamic light use and protection from excess light in upper canopy and coppice leaves of Nothofagus cunninghamii in an old growth, cool temperate rainforest in Victoria, Australia[J]. New Phytologist,2005,165(1):143-156.
[22] 于贵瑞,王秋凤.植物光合、蒸腾与水分利用的生理生态学[M].北京:科学出版社,2010.
[23] 龚伟,宫渊波,胡庭兴. CO2浓度升高对湿地松针叶蒸腾特性和水分利用效率的影响[J].水土保持学报,2005,19(5):178-182.
[24] 崔大练,马玉心,蔡体久. CO2摩尔分数升高对兴安鹿蹄草光合生理特性的影响[J].东北林业大学学报,2013,41(1):94-99.
[25] 刘玉华,贾志宽,史纪安,等.旱作条件下不同苜蓿品种光合作用的日变化[J].生态学报,2006,26(5):1468-1477.
[26] 徐俊增,彭世彰,丁加丽,等.控制灌溉的水稻气孔限制值变化规律试验研究[J].水利学报,2006,37(4):486-491.
[27] Yu G R, Song X, Wang Q F, et al. Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables[J]. New Phytologist,2008,177(4):927-937.
[28] Berry J A, Beerling D J, Franks P J. Stomatal:key players in the earth system, past and present[J]. Current Opinion in Plant Biology,2010,13(3):233-239.
[29] 付勇,汪立今,柴凤梅,等.多元线性回归和逐步回归分析在白石泉Cu—Ni硫化物矿床研究中的应用[J].地学前缘,2009,16(1):373-379.
[30] 张宁宁,刘普灵,王栓全,等.黄土丘陵区不同经济作物光合作用特性研究[J].水土保持研究,2012,19(3):269-273.
[31] 李铭怡,刘刚,许文年,等.新银合欢在不同土壤水分条件下的适生性研究[J].水土保持研究,2013,20(2):259-266.
[32] 肖文发,徐德应,刘世荣,等.杉木人工林针叶光合与蒸腾作用的时空特征[J].林业科学,2002,38(5):38-46.
[33] 许大全.光合作用“午睡”现象的生态、生理与生化[J].植物生理学通讯,1990,26(6):5-10.
[34] 张启昌,杜凤国,夏富才,等.美国椴光合蒸腾的生理生态[J].北华大学学报:自然科学版,2000,1(5):436-438.
[35] 徐敏,骆争荣,于明坚,等.百山祖北坡中山常绿阔叶林的物种组成和群落结构[J].浙江大学学报:农业与生命科学版,2007,33(4):450-457.
StudyonPhotosynthesis-transpirationCharacteristicsofTwoBroadleafWoodyPlantSpeciesintheForestsofBaishanzu
WU Yan-wen, ZHANG Jin-chi, GU Zhe-yan, LIU Xin, HAN Cheng
(NanjingForestryUniversity,Nanjing210037,China)
Photosynthesis and transpiration play the significant roles in the evaluation of ecological benefits from a forest ecosystem. In order to determine their photosynthesis and transpiration characteristics at canopy level, this paper conducted some measurements with Li-6400 portable photosynthetic apparatus on two typical tree species in the forest ecosystem of Fengyang Mountain. Results show that the main factors influencingPnofStewartiaacutisepalawere WUE and PYR, meanwhile, the main factors influencingPnofPterostyraxcorymbosuswereCiand PYR. In the process of daily variation ofPn, the value ofStewartiaacutisepalawas always higher thanPterostyraxcorymbosus. It can be judged that the photosynthesis ofStewartiaacutisepalawas stronger thanPterostyraxcorymbosus. Additionally, the main factors influencingTrofStewartiaacutisepalawereGsand PAR, while the main factors influencingTrofPterostyraxcorymbosuswereGsand RH. According to daily variation ofTr, the transpiration ofStewartiaacutisepalawas stronger thanPterostyraxcorymbosus. According to the analysis of WUE, the carbon water balance ability ofStewartiaacutisepalawas better thanPterostyraxcorymbosus. Therefore, the adaptability ofStewartiaacutisepalatothis habitat was superior toPterostyraxcorymbosus, and the former was more widely distributed.
Baishanzu; net photosynthetic rate; transpiration rate; physiological factor; ecological factor
2013-11-25
:2013-12-11
国家公益林行业重大专项“典型森林植被对水资源形成过程的调控研究”(201104005)
吴雁雯(1989—),女,江苏常州人,博士,主要从事水土保持和荒漠化防治。E-mail:wywhsl@sina.com
张金池(1962—),男,山东潍坊人,教授,博导,主要从事水土保持和荒漠化防治。E-mail:nfujczhang@sina.com
S718.43
:A
:1005-3409(2014)06-0204-07
