短花针茅荒漠草原物种数空间变化特点及其与取样方法的关系
2014-09-20贾利娟桑雪颖李慧娟吕世杰苏金梅刘红梅
贾利娟,桑雪颖,李慧娟,吕世杰,苏金梅,乌 兰,刘红梅
(1.内蒙古苏尼特右旗草原工作站,内蒙古 赛罕塔拉 011200;2.内蒙古农业大学理学院 内蒙古 呼和浩特 010018;3.内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;4.内蒙古林业科学研究院,内蒙古 呼和浩特 010010)
短花针茅荒漠草原物种数空间变化特点及其与取样方法的关系
贾利娟1,桑雪颖2,李慧娟2,吕世杰2,苏金梅2,乌 兰1,刘红梅3,4
(1.内蒙古苏尼特右旗草原工作站,内蒙古赛罕塔拉011200;2.内蒙古农业大学理学院内蒙古呼和浩特010018;3.内蒙古农业大学生态环境学院,内蒙古呼和浩特010019;4.内蒙古林业科学研究院,内蒙古呼和浩特010010)
以短花针茅荒漠草原物种数为研究对象,采用样线法和样地法两种取样方法,对短花针茅荒漠草原物种数空间分布特点及其与不同取样方法的关系进行初步探讨,结果表明:短花针茅荒漠草原物种数在不同样方间的变化为4~17种/m2。样线法取样时平均物种数为9~10种/m2,空间变化的结构比随样线长度的增加整体呈减小的变化趋势,分形维数在样线长度为1500m时最小,采用样线法取样时样线长度应该在1500m~2000m比较适宜。样地法取样时平均物种数为11~12种/m2,样地法取样时结构比随样地面积的增加呈先增加后稳定的变化趋势,分形维数随样地面积的增加而逐渐增大,样地取样面积在6400m2~8100m2比较适宜。样地法取样会在两个方向上同时获得荒漠草原物种数空间分布信息,更能够全面地反映荒漠草原物种数空间分布特点。
荒漠草原;短花针茅;空间异质性;取样方法
1 引言
探索植物群落空间异质性的性质,分析植物群落空间异质性的原因和潜在的生态学效应,有助于从空间透视的角度,更清楚地揭示植物群落格局与生态学过程及其机制。因此,20世纪90年代以来,生态学中的空间异质性研究成为倍受关注的一个极具潜力的理论问题〔1,2〕。Li和Reynolds认为,异质性可根据两个组分来定义,即所研究的系统属性及其复杂性或变异性,这种观点强调空间结构特征的可观察性及其可定量分析性,以及空间尺度依赖性〔3-5〕。邬建国在讨论空间异质性与缀块性时指出,空间异质性是某种生态学变量在空间上的不均匀性及复杂程度,是空间缀块性和空间梯度的综合反映。缀块性强调缀块的种类组成特性及其空间分布与配置的关系,而空间梯度则指沿某一方向景观特征有规律地逐渐变化的空间特征〔6〕。国内学者从种群和群落的角度对马尾松、野生甘草、泡泡刺灌丛、小叶锦鸡儿群落等进行了空间异质性分析〔7-10〕,国外研究者对这方面的研究较多,如探索植物群落斑块特征(如斑块大小、质量、密度、高度、生物量等)对动物采食选择的影响〔11-14〕。
本文采用样线法和样地法两种取样方法,以短花针茅荒漠草原植物群落物种数为研究对象,利用地统计学软件GS+计算结构比和分形维数,探讨短花针茅荒漠草原物种数的一维和二维空间变化特征,并对两种取样方法下的短花针茅荒漠草原植物群落物种数空间特征进行比对,为选用合理的取样方法,详细了解短花针茅荒漠草原植物群落物种数空间变化情况提供基本的理论依据和数据支持。
2 试验设计和研究方法
2.1试验设计
试验区位于内蒙古高原荒漠草原亚带南侧呈条状分布的短花针茅草原的东南部,地理位置在锡林郭勒盟苏尼特右旗朱日和镇附近的都呼木苏木,E112°47′16.9″,N42°16′26.2″。在试验区内采用两种取样方法并对样方内物种数进行统计,详情如下。
1)样线法
在样地随机一点由西向东间隔10m取样,样方面积1m×1m,样方数和样线长度如表1所示,样线长度和样方数均以等差数列方式增长,样线长度以an=a1+(n-1)×500幅度增加(其中a1=500,n=1,2,3,4,5,6),样方数以bn=b1+(n-1)×50幅度增加(其中b1=51,n=1,2,3,4,5,6)。
2)样地法
随机选择一点,定义为平面坐标原点,然后按机械取样法每隔10米进行网格取样,样方面积1m×1m,样方数和样地面积见表1,样地面积增加幅度以an=a1×q2方式增加(其中a1=100,q=6,7,8,9,10),样方数以bn=q2幅度增加(其中q=7,8,9,10,11)。

表1 取样方法
2.2研究方法
利用GS+软件,采用地统计学的半方差函数分析和分形维数分析方法,计算结构比和分维数〔15,16〕。
3 结果与分析
3.1物种数的变化
3.1.1 不同样线长度物种数变化
不同样线长度样方内物种数的变化见表2所示,当样线长度为500m时,样方内平均物种数最大,为11.75(P<0.05),当样线长度增加到1000m时,样方内平均物种数为10.35,其显著小于样线长度为500m时样方内平均物种数(P<0.05),当样线长度继续增大时,样方内平均物种数尽管显著小于线长度为1000m时的物种数(P<0.05),但随着样线长度的继续增加,样方内平均物种数已不再发生显著性变化。这表明当样线长度为1500m、样方数为151个时,样方内平均物种数已趋于稳定,即短花针茅荒漠草原物种数大约集中在9~10种/m2。
标准差表示的是样方间物种数与平均物种数的差异程度,由下表可知,在500~1500m之间,样方间物种数与平均物种数的差异程度在逐渐增大,在1500~3000m之间,样方间物种数与平均物种数的差异程度在逐渐减小。表明在1500m样线内,样方间的物种数变化较为强烈。标准误随着样线长度的增加逐渐减小,表明不同样线长度测定的物种数越来越接近平均物种数。变异系数的变化趋势与标准差相同,表明单位均值上的物种数变化也呈现出先增大后减小的变化趋势,且在500m样线内,样方间的物种数变化最为强烈。
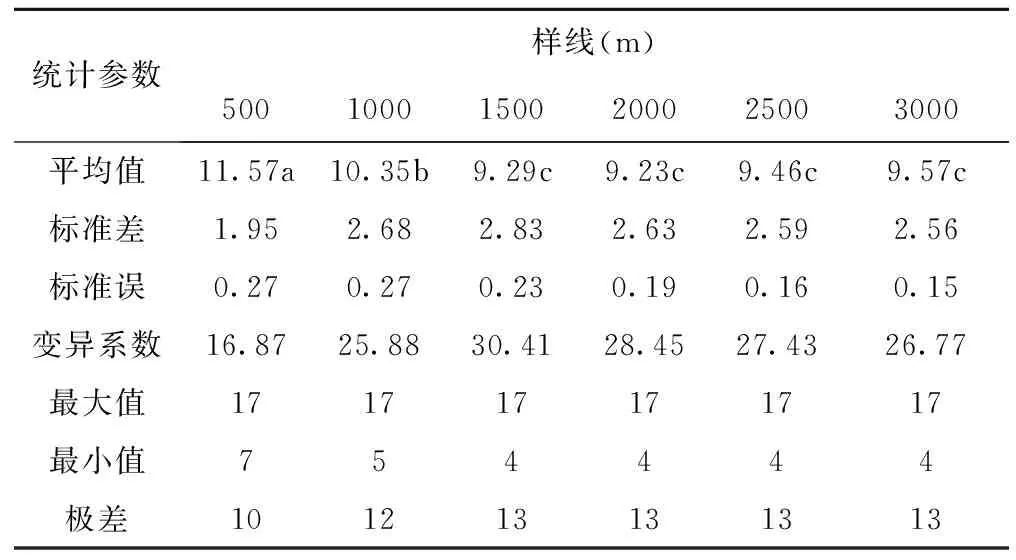
表2 不同样线长度物种数变化
3.1.2 不同样地面积物种数变化
不同样地面积样方内物种数变化见表3。尽管取样面积在不断增大,但样方内平均物种数始终在11~12种/m2之间。标准差、标准误和变异系数均呈现出随样地面积的增大而减小的变化趋势。这表明,随着样地面积的增大,样方间物种数的变动越来越与平均物种数接近。
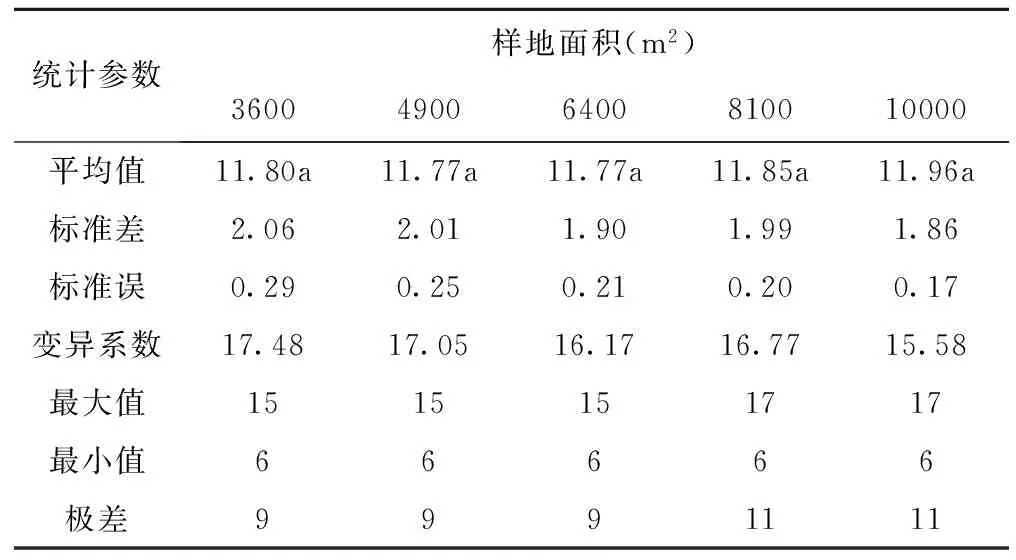
表3 不同样地面积物种数变化
3.2结构比变化
3.2.1 不同样线长度物种数空间变化的结构比
结构比反映的是结构因素影响的空间变异占最大空间变异的比例,最大空间变异由结构因素影响的空间变异和随机因素影响的空间变异共同决定,因此,结构比也可以反映随机因素影响的空间变异占最大空间变异的比例。
表4为不同样线长度物种数空间变化的结构比。从表中可以看到,随着取样面积的增加,结构比大体呈现出逐渐减小的变化趋势。在500~1500m之间,随着样线长度的增加,物种数空间变化的结构比存在波动,当样线长度大于1500m,物种数空间变化的结构比逐渐减小。这表明,在500~1500m之间,随机性因素引起的物种数空间变异占最大空间变异的比例波动较大,导致结构比波动较大;而在样线长度大于1500时,随机因素引起的物种数空间变异占最大空间变异的比例逐渐增大,结构比逐渐减小。表明在短距离内,短花针茅荒漠草原物种数分布主要以结构性因素起决定作用,而在样线长度较大时,随机性因素对物种数分布的影响逐渐增强。
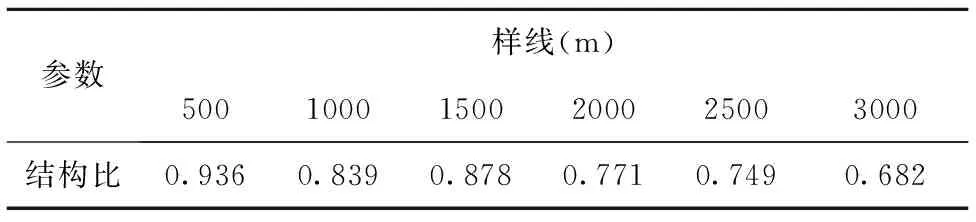
表4 不同样线长度物种数空间变化的结构比
3.2.2 不同样地面积物种数空间变化的结构比
不同样地面积物种数空间变化的结构比在小样地范围内(3600m2)具有较低的结构比(见表5),表明在进行空间异质性分析时,样地面积较小可能会导致随机取样时随机因素在总变异中的比例加大。当取样面积在4900m2~10000m2之间时,物种数空间变化的结构比在0.915~0.948之间波动,波动幅度为0.033,这说明当取样面积大于4900m2时,物种数空间变化的结构比会维持在一个相对稳定的区间内。也进一步说明,在短花针茅荒漠草原进行物种数空间变化分析时,取样面积达到4900m2时已经能够代表短花针茅荒漠草原物种数空间变化情况。
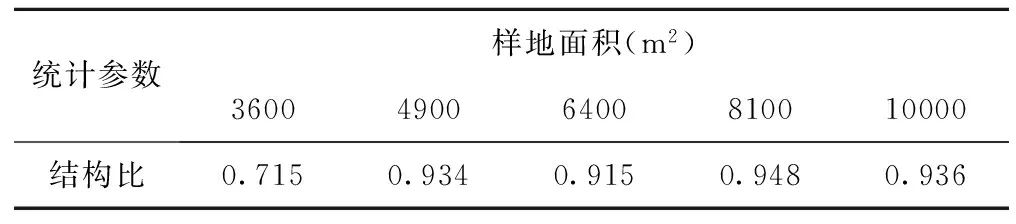
表5 不同样地面积物种数空间变化的结构比
3.3分形维数变化
3.3.1 不同样线长度物种数空间变化的分形维数
不同样线长度物种数空间变化的分形维数计算结果见表6。在样线长度为500~1500m时,分形维数会急剧降低,在1500~3000m样线长度范围内,物种数空间变化的分形维数逐渐增大。表明样线长度不同,分形维数结果会有差异。这种差异反映的是短花针茅荒漠草原物种数空间变化强烈程度,反映的是自相关变量空间分布格局的复杂程度;分维数高,意味着物种数空间分布格局简单,空间依赖性越强,空间结构性好;分维数低意味着空间分布格局相对复杂,随机因素引起的异质性占有较大的比重。因此,在样线长度为500~1500m时,物种数空间变化格局越来越复杂,空间依赖性越来越小,空间结构性越来越差;在1500~3000m样线长度范围内,物种数空间变化格局由复杂向简单化方向发展,空间依赖性逐渐增强,随机因素引起的异质性逐渐减弱。
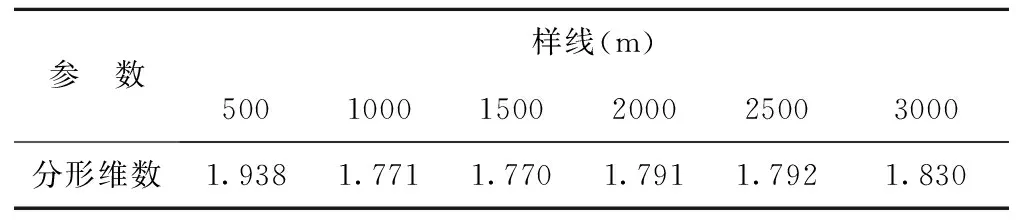
表6 不同样线长度物种数空间变化的分形维数
3.3.2 不同样地面积物种数空间变化的分形维数
不同样地面积物种数空间变化的分形维数在3600m2~4900m2时迅速增大,在4900m2~10000 m2范围内始终处于增加状态,但增加幅度比较小(见表7)。表明样地面积小,随机因素对物种数空间变化的影响较大,随着样地面积的逐渐增大,结构性因素对荒漠草原物种数的影响越来越大。
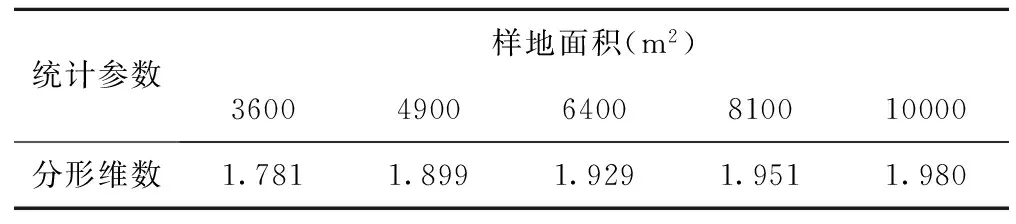
表7 不同样地面积物种数空间变化的分形维数
4 讨论
无论是样线法取样还是样地法取样,在进行荒漠草原物种数空间变化分析时,均能够不同程度的反映荒漠草原物种数空间变化特点。但由于受样线长度和样地面积的影响,荒漠草原物种数空间变化会有很大的差异,这种差异研究者应该在试验设计的时候就应该考虑到。区组试验设计能够对单向倾向性影响进行控制〔17,18〕,而拉丁方试验设计能够对双向倾向性影响进行控制〔18〕,样线法取样和样地法取样尽管不涉及试验控制问题,但由于草地水平结构的空间异质性是平面结构(一定范围内),不是线性变化。所以在试验区内探讨空间异质性时,采用样地法更为合适;当探讨某一梯度异质性问题,且梯度变化空间较大,样地法取样受人力、物力、财力限制时,可采用样线法。
当样线长度偏小,物种数空间异质性分布的参数——结构比及分形维数计算结果会偏大,致使物种数空间分布受结构性因素影响占最大空间变异的比例加大,导致研究者直观地认为物种数空间分布过程中结构性因素占有重要地位,从而对研究荒漠草原物种数空间分布规律产生不利的影响;样线长度过长,会导致结构比偏小,导致研究者直观地认为物种数空间分布过程中随机因素影响较大。
当样地面积偏小,物种数空间异质性分布的参数——结构比及分形维数计算结果会偏小,致使物种数空间分布受结构性因素影响占最大空间变异的比例缩小,导致研究者错误地认为物种数空间分布过程中随机性因素占有重要地位,从而对研究荒漠草原物种数空间分布也产生不利的影响。当样地面积过大,结构比及分形维数计算结果会偏大。因此,无论是样线法还是样地法,取样距离或者取样面积均不宜过小,样线法的取样距离或者样地法取样面积过小,可能会产生不同的结论。
从本文分析结果看,采用样线法取样,当样方间距为10m时,样线长度应该在1500m~2000m比较适宜;当采用样地法,样方间距为10m时,样地面积在6400m2~8100 m2比较适宜。从分析结果中也可以看到,样线法取样计算的结构比和分形维数呈线性变化比较明显,而样地法取样计算的结构比和分形维数在样地面积大于某一数值时,其会呈现波动或小幅度增加状态。原因是样线法取样只能得到一个方向上荒漠草原物种数空间分布特征(即一维空间分布特征),而样地法取样会在两个方向上同时获得荒漠草原物种数空间分布信息(即二维空间分布特征)。因此,就取样方法而言,样地法取样更能够全面地反映荒漠草原物种数空间分布特点。
由于空间异质性研究需要空间尺度进行限定,当研究尺度存在变化,荒漠草原物种数空间分布的特点也会发生相应的变化,对空间异质性的分析结果也就产生差异。所以当进行空间异质性分析时,小尺度小到什么程度,大尺度大到什么程度,研究者应该根据自己的研究目的和研究内容以及人力、物力和财力等综合考虑决定。因此,先确定研究尺度,再研究变量空间变化特点会达到事半功倍的效果。
5 结论
5.1 荒漠草原物种数在不同样方间的变化为4~17种/m2,样线法取样时平均物种数为9~10种/m2,样地法取样时平均物种数为11~12种/m2。
5.2 样线法取样时荒漠草原物种数空间变化的结构比随样线长度的增加整体呈减小的变化趋势,样地法取样时结构比随样地面积的增加呈先增加后稳定的变化趋势。
5.3 荒漠草原物种数空间变化的分形维数在样线长度为1500m时最小。样地法取样时随样地面积的增加而逐渐增大。
5.4 研究荒漠草原物种数空间分布特点采用样线法时,样线长度应该在1500m~2000m比较适宜;当采用样地法,样地面积在6400m2~8100 m2比较适宜。
〔1〕王政权.地统计学及在生态学中的应用〔M〕.北京:科学出版社. 1999.
〔2〕王政权,王庆成,李哈滨.红松老龄林主要树种的空间异质性特征与比较的定量研究〔J〕.植物生态学报,2000,24:718-723
〔3〕Li H. and Reynolds J. F. On definition and quantification of heterogeneity. Oikos, 1995, 73: 280-284.
〔4〕Li H, Reynolds JF. 1994. A simulation experiment to quantify spatial heterogeneity in categorical maps. Ecology, 75: 2446-2455
〔5〕韩有志.林分空间异质性与水曲柳的更新格局和过程〔D〕.东北林业大学博士学位论文.2001:89-102
〔6〕邬建国.景观生态学一格局,过程,尺度与等级〔M〕. 北京,高等教育出版社,2000.
〔7〕陈美高.马尾松种源胸径生长的空间异质性〔J〕.福建林学院学报,2005,25(4):333-337
〔8〕沈海亮,王季槐,李明.宁夏野生甘草分布空间异质性及分布格局研究〔J〕.草业科学,2007,24(7):18-22
〔9〕刘冰,赵文智.荒漠绿洲过渡带泡泡刺灌丛沙堆形态特征及其空间异质性〔J〕.应用生态学报,2007,18(12):2814-2820
〔10〕蒋德明,李明,押田敏雄等.封育对科尔沁沙地小叶锦鸡儿群落植被特征及空间异质性的影响〔J〕.生态学杂志,2009,28(11):2159-2164
〔11〕Wallis DeVries,M F,Daleboudt C.Foraging strategy of cattle in patchy grassland〔J〕. Oecologia, 1994, 100:98-106
〔12〕Wilmshurst J F,Fryxell J M,Hudsonb R J.Forage quality and patch choice by wapiti(Cervuselaphus)〔J〕.Behav Ecol,1995,6:209-217
〔13〕Hjalten J,Danell K,Lundberg P.Herbivore avoidance by association:vole and hare utilization of woody plants〔J〕.Oikos,1993,68:125-131
〔14〕Searle K R,Hobbs N T,Shipley L A.Should I stay or should I go?patch departure decisions by herbivores at multiple scales〔J〕.Oikos,2005,111:417-424
〔15〕刘红梅,卫智军,杨静,等. 不同放牧制度对荒漠草原表层土壤氮素空间异质性的影响〔J〕. 中国草地学报,2011, 33(2): 51-56+74.
〔16〕刘红梅, 卫智军, 杨静, 等. 不同放牧制度对荒漠草原短花针茅空间异质性的影响〔J〕. 干旱区资源与环境, 2011, 25(8): 138-143.
〔17〕李喜春, 姜丽娜, 邵云, 等. 生物统计学(第五版)〔M〕. 北京: 科学出版社, 2014.
〔18〕盖钧镒. 试验统计方法〔M〕. 北京: 中国农业出版社, 2000.
S812
A
2095—5952(2014)04—0030—05
2014-11-06
内蒙古自治区科技计划项目“不同类型草原区围封草地利用模式研究”;内蒙古农业大学创新基金“短花针茅荒漠草原植物群落地上地下现存量比较与模拟分析”。
贾利娟(1988-),女,内蒙古苏尼特右旗人,助理畜牧师,从事草原保护建设及技术推广工作。
吕世杰(1978-),男,汉族,研究方向为生物统计与数量生态,E-mail:lshj123@163.com。
