大兴安岭5种类型低质林土壤呼吸日变化及影响因素1)
2014-09-18宋启亮董希斌
宋启亮 董希斌
(东北林业大学,哈尔滨,150040)
森林生态系统作为陆地生态系统中重要的碳汇与碳源,其在调控生态系统碳平衡方面有着不可替代的作用。森林土壤是森林生态系统中最大的碳库,固碳作用明显,而森林土壤呼吸作为森林土壤碳输出的主要途径,显著影响着空气中CO2的浓度,成为全球碳循环研究的热点[1-2]。森林土壤呼吸主要包含植被根系呼吸、土壤微生物呼吸、土壤动物呼吸以及土壤中含碳物质的化学氧化过程[3-4]。不同类型森林生态系统中各组分对土壤呼吸的贡献程度有很大差异,这是由于土壤呼吸不仅受森林植被类型、根系生物量、土壤微生物种群数量等生物因子的影响,而且还受气温、降雨、枯落物、土壤理化性质、土壤温度、土壤湿度等非生物因子的影响[5-7]。土壤呼吸速率还存在时间上的差异性,而土壤温度是造成土壤呼吸速率日变化的主要限制因素[8]。研究森林土壤呼吸的变化及不同因子对土壤呼吸的影响,并建立各因子与土壤呼吸间的关系模型,可以为准确预测气候变化提供基础,同时也能预测森林土壤肥力及森林生态系统的生产力[9-10]。
大兴安岭林区作为我国重要的林业生产基地,在经过多年的过度开发及自然灾害破坏后,形成了大量生态功能脆弱的低质林。本研究即以大兴安岭林区针阔混交低质林、山杨(Populus davidiana)低质林、蒙古栎(Quercus mongolica)低质林、白桦(Betula platyphylla)低质林和阔叶混交低质林5种类型低质林为研究对象,采用LI-8150多通道土壤呼吸自动测量系统测定不同类型低质林土壤呼吸速率的日变化,并测定观测点的土壤温度、湿度、理化性质和枯落物,建立土壤呼吸速率与土壤温度、湿度的单因素关系模型,以及土壤呼吸速率与土壤温湿度的双因素复合模型,分析土壤理化性质和枯落物对土壤呼吸速率的影响,为大兴安岭地区不同类型低质林生态系统的研究提供理论参考。
1 研究区概况
研究区设置在加格达奇林业局跃进林场和翠峰林场,低山丘陵地带,平均海拔500 m左右,平均坡度在10°以下。林地土壤以暗棕壤为主,厚度10~25 cm。无霜期80~110 d,年平均降水量500 mm以上,属寒温带大陆性季风气候,冬长夏短,冬季寒冷,夏季炎热多雨。在跃进林场51林班选取针阔混交低质林;在跃进林场54林班选取山杨低质林;在翠峰林场174林班选取蒙古栎低质林、白桦低质林和阔叶混交低质林3种典型低质林类型进行研究,5种类型低质林的立地条件和林分概况见选取表1。

表1 样地概况
2 研究方法
在大兴安岭林区加格达奇林业局跃进林场和翠峰林场选取最典型的5种类型低质林进行研究,分别在5种森林群落中设置3个20 m×20 m的典型样地,共15块样地。在每个样地按“Z”型设置5个观测点,于2013年6月底进行土壤呼吸及土壤、枯落物因子的测定。
土壤呼吸测定:采用LI-8150多通道(本研究应用5通道)土壤呼吸自动测量系统测定土壤呼吸速率,提前24 h将PVC土壤环踩入土壤中,高出地面2~3 cm,并保持土壤环内枯落物的原始自然状态。每个观测点连续观测24 h,以0.5 h为一个测量周期,全天重复测量48次。采用LI-8150系统配套的E型热电偶土壤温度探头和EC-5土壤水分传感器测量观测点10 cm深处的土壤温度和湿度。
土壤理化性质和枯落物的测定:土壤呼吸测量完成后,将土壤环内枯落物按未分解和半分解层全部收集,然后用容积为100 cm3的环刀取样,用于分析土壤的物理性质,同时取1 kg土壤剖面0~10 cm的土壤带回实验室,在实验室做自然风干处理,研细过筛,采用森林土壤分析方法测定土壤的化学性质。
应用 LI-8150配套的 File Viewer v3.0.0软件将测得的土壤呼吸速率、土壤温度、土壤湿度等数据导出,利用Excel软件对数据进行统计整理,绘制各类型低质林样地土壤呼吸速率日变化曲线、土壤呼吸速率随10 cm深处土壤温度及湿度的变化曲线。应用SPSS17.0进行数据分析,对各样地土壤呼吸速率进行ANOVA分析;计算各类型低质林样地土壤呼吸速率的平均值和标准差;并在0.05显著水平进行LSD检验;运用线性和非线性回归模型分析土壤呼吸速率与土壤温湿度之间的关系;采用Spearman相关性检验,对土壤呼吸速率与土壤理化性质、枯落物相关指标的相关性进行分析。
3 结果与分析
3.1 土壤呼吸速率日变化及差异分析
不同类型低质林土壤呼吸速率日变化见图1,各个类型低质林样地的土壤呼吸速率日变化整体上呈现单峰趋势,但各个时间段土壤呼吸速率有一定的波动。针阔混交低质林样地土壤呼吸速率最大值8.54 μmol·m-2·s-1出现在 13:00,最小值 5.44 μmol·m-2·s-1出现在02:00;山杨低质林样地土壤呼吸速率最大值 6.84 μmol·m-2·s-1出现在 12:00,最小值 4.49 μmol·m-2·s-1出现在 01:00;蒙古栎低质林样地土壤呼吸速率最大值5.56 μmol·m-2·s-1出现在 14:00,最小值 3.28 μmol·m-2·s-1出现在3:00;白桦低质林样地土壤呼吸速率最大值7.54 μmol·m-2·s-1出现在 12:00,最小值 4.45 μmol·m-2·s-1出现在23:00;阔叶混交低质林样地土壤呼吸速率最大值 8.11 μmol·m-2·s-1出现在 13:00,最小值 5.25 μmol·m-2·s-1出现在 23:00。综合分析发现一天当中土壤呼吸速率一般在12:00—14:00最高,在23:00—03:00最低,各类型低质林样地白天土壤呼吸速率均高于夜间土壤呼吸速率,这与土壤温度、土壤湿度等条件有关。
经过方差分析发现,不同类型低质林样地土壤呼吸速率存在显著性差异(见表2),由表3可知,5种类型低质林样地土壤呼吸速率日平均值从低到高依次为:蒙古栎低质林、白桦低质林、山杨低质林、阔叶混交低质林、针阔混交低质林。LSD检验表明,蒙古栎低质林土壤呼吸速率日平均值最低,且与其他类型低质林差异显著(P<0.05),白桦低质林与山杨低质林之间差异不显著,阔叶混交低质林和针阔混交低质林土壤呼吸速率日平均值之间差异同样不显著。

图1 不同类型低质林样地土壤呼吸日变化
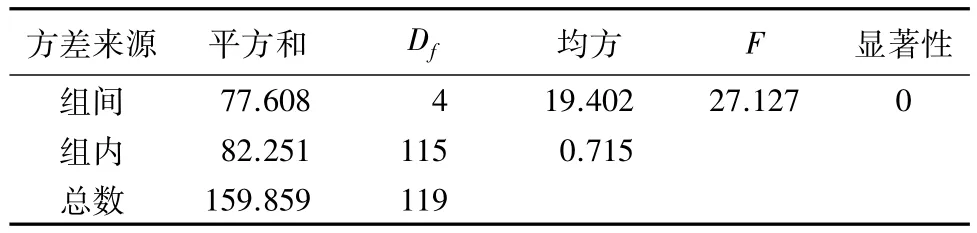
表2 5种类型低质林土壤呼吸速率方差分析

表3 5种类型低质林土壤呼吸速率平均值
3.2 土壤呼吸速率与土壤温湿度的关系
土壤温度是影响土壤呼吸的最主要因素之一,几乎影响着土壤呼吸过程的各个方面,土壤温度与土壤呼吸之间的关系已经存在很多经验模型,其中Van’t Hoff提出的经验指数模型,对于处在一定温度范围内的生态系统来说,是普遍适用的,应用最为广泛。5种类型低质林样地的土壤呼吸速率都随土壤温度的升高而上升,二者之间的关系见图2,拟合后发现土壤呼吸速率与10 cm深处土壤温度间适合指数模型Rs=αeβT,式中:Rs为实验测量的土壤呼吸速率(μmol·m-2·s-1);T为距地表下10 cm处的土壤温度(℃);α为0℃时的土壤呼吸速率;β为温度响应系数。一般用Q10来描述土壤呼吸过程对土壤温度的敏感性,它表示温度每升高10℃土壤呼吸速率增加的倍数,Q10的表达式为:Q10=e10β。

图2 不同类型低质林样地土壤呼吸速率和土壤温度的关系
用散点图描绘5种不同类型低质林样地的土壤呼吸和土壤温度的关系(见图2),发现土壤呼吸速率随着土壤温度的升高而逐渐升高,对其进行回归分析(见表4),发现二者之间呈现显著的指数关系(R2为 0.73~0.82),其中阔叶混交低质林土壤呼吸速率和土壤温度之间的关系最显著。各类型低质林样地 Q10值在 2.06~2.87之间,其中白桦低质林的Q10值2.87最大,说明白桦低质林林下土壤呼吸速率对距地表下10 cm的土壤温度敏感性最强。

表4 土壤呼吸速率和土壤温度的关系方程
用散点图描绘5种不同类型低质林样地的土壤呼吸和土壤湿度的关系(见图3),发现各样地土壤呼吸速率随着土壤湿度的升高而逐渐上升,但当土壤湿度升高到一定值后土壤呼吸速率不再升高,反而随着土壤湿度的升高而呈现下降的趋势,不同类型低质林土壤湿度的阈值不同。因此,本研究采用二次多项式模型对土壤呼吸速率和土壤湿度的关系进行模拟,模型表达式为:Rs=aW2+bW+c,式中,Rs为土壤呼吸速率(μmol·m-2·s-1);W为10 cm深处土壤湿度(%);a、b、c为待定参数。模拟结果见表5,由结果可知,不同类型低质林土壤呼吸速率和土壤湿度呈显著的二次多项式关系(R2为0.61~0.85),其中蒙古栎低质林土壤呼吸速率和土壤湿度之间的相关性最高。
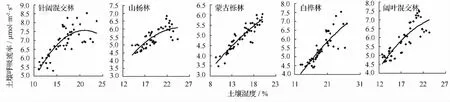
图3 不同类型低质林土壤呼吸速率和土壤湿度的关系

表5 土壤呼吸速率和土壤湿度的关系方程
采用线性和非线性复合模型分析土壤呼吸速率与10 cm深处土壤温湿度的复合关系(表6)。结果发现,与单因子模型相比,除针阔混交低质林和山杨低质林应用模型R=aTW+b拟合效果不如单因子(温度)模型外,其余各类型低质林不同双因子复合模型的决定系数R2值都高于单因子模型,R2值为0.65~0.905,表明用双因子复合模型对土壤呼吸速率的预测更准确。从拟合的效果看,不同类型低质林的最佳复合模型并不相同,针阔混交低质林和蒙古栎低质林应用关系模型Rs=aTbWc,模拟效果最佳,决定系数分别为 0.79、0.887;山杨低质林、白桦低质林和阔叶混交低质林应用关系模型Rs=aebTWc,模拟效果最佳,决定系数分别为 0.803、0.815、0.905。因此,为了更加准确地预测土壤呼吸速率,应该比较多个模型,从中选取最优的一个。

表6 土壤呼吸速率与土壤温度及土壤湿度的复合关系方程
3.3 土壤呼吸速率影响因子的相关性分析
不同类型低质林样地土壤呼吸速率与土壤理化性质、枯落物蓄积量等指标做Spearman相关性检验,从表7可知,不同类型低质林样地土壤呼吸速率与土壤密度呈显著的负相关性(P<0.05);而与土壤孔隙度呈显著的正相关性(P<0.05);山杨低质林、蒙古栎低质林和阔叶混交低质林样地土壤pH值与土壤呼吸速率呈显著的正相关性;各类型低质林样地土壤呼吸速率与土壤有机质质量分数均呈显著的正相关性;除山杨低质林样地土壤呼吸速率与土壤全N质量分数呈显著的正相关性外,其他类型低质林样地土壤呼吸速率与土壤全量养分(全N、全P、全K)质量分数均未通过Spearman相关性检验(P>0.05);白桦低质林和阔叶混交低质林样地土壤呼吸速率与土壤水解N质量分数相关性显著,其他类型样地土壤呼吸速率与土壤有效养分(水解N、有效P、速效K)质量分数相关性不显著;各类型低质林样地土壤呼吸速率与未分解层枯落物蓄积量相关性不显著,蒙古栎低质林、白桦低质林和阔叶混交低质林样地半分解层枯落物的蓄积量与土壤碳通量有显著的正相关性。

表7 土壤呼吸速率与各指标的相关性
4 结论与讨论
本文研究结果表明:不同类型低质林样地白天土壤呼吸速率均高于夜间土壤呼吸速率,一天当中土壤呼吸速率最高值一般出现在12:00—14:00,而最低值在23:00—03:00,这与土壤温度和湿度对植物生理作用过程和微生物活性的影响有关[11]。5种类型低质林样地土壤呼吸速率日平均值依次为:针阔混交低质林、阔叶混交低质林、山杨低质林、白桦低质林、蒙古栎低质林,蒙古栎低质林土壤呼吸速率(4.62 μmol·m-2·s-1)最低,且与其他类型低质林差异显著(P<0.05),白桦低质林(5.41 μmol·m-2·s-1)与山杨低质林(5.47 μmol·m-2·s-1)之间,阔叶混交低质林(6.41 μmol·m-2·s-1)与针阔混交低质林(6.86 μmol·m-2·s-1)之间差异不显著,这与低质林植被类型产生的呼吸作用、根生物量、土壤微生物量、枯落物及土壤有机质含量的不同有关,同时,林内小型动物分布的差别也是造成这一现象的原因[12]。
土壤温度不但可以影响酶的活性和土壤生物,而且可以通过底物供应对土壤呼吸产生影响[13]。大多数研究认为土壤呼吸速率与土壤温度正相关,本研究经拟合发现大兴安岭5种类型低质林样地土壤呼吸速率与土壤温度的关系都适合指数模型(R2为 0.73~0.82)。Q10值是反映土壤呼吸速率对土壤温度敏感性的指标[14],目前不同的研究Q10值差异较大[7,14-16],这是由于 Q10值随时间、地理位置、土壤以及生态系统类型的变化而发生较大的改变[13,17],全球Q10的中间值为2.4[18]。本文得到的针阔混交低质林、山杨低质林、蒙古栎低质林、白桦低质林、阔叶混交低质林在植物生长季的Q10值分别为2.66、2.06、2.10、2.87 和 2.43,由于大兴安岭林区位于北半球高纬度地区,各类型低质林土壤呼吸速率对温度的敏感性较高,不同类型低质林Q10值不同是受植物根系、土壤微生物的种类和数量的影响,而且各种环境因子调控着底物的供应状况[19]。
土壤湿度对土壤呼吸的影响机理较为复杂[20],其不仅直接影响植物根系和微生物的生理过程,而且也间接的影响着底物和氧气的扩散,不同研究发现土壤湿度和土壤呼吸的关系并不相同,有研究认为土壤水分对土壤呼吸的影响较小[21-22],而有研究认为二者之间存在显著的相关性[10,23]。这主要是由于研究区域、立地条件、植被类型等多种因素不同所造成的。本研究发现大兴安岭地区不同类型低质林样地土壤呼吸速率和土壤湿度(8%~27%)呈显著的二次曲线关系(R2为 0.61~0.85),土壤呼吸速率在干燥条件下较低,与土壤湿度正相关,在中等土壤湿度水平时最大,超过阈值后,土壤呼吸速率与土壤湿度负相关[10],当湿度很高、厌氧条件占优势,导致好氧微生物的活性受到抑制时又下降,最适的含水量是接近田间持水量,这时大孔隙空间大部分充满空气,利于氧气扩散,小孔隙空间大部分充满水,利于可溶性底物的扩散[24]。不同植被类型样地土壤湿度对土壤呼吸速率的影响程度不同。
不同研究中使用不同形式的方程来描述土壤温湿度对土壤呼吸速率的影响[25-26],研究发现土壤温湿度双因子复合模型对土壤呼吸速率的预测更准确[10,27-28],这是由于土壤温湿度交互作用影响着土壤呼吸过程[21]。本研究中,与单因子模型相比,除针阔混交低质林和山杨低质林应用模型R=aTW+b拟合效果不如温度模型外,其余各类型低质林土壤呼吸速率的线性和非线性双因子复合模型的决定系数R2值均有所提高,土壤温湿度能够共同解释各类型低质林土壤呼吸速率的65%~90.5%。不同类型低质林的最佳复合模型并不相同,所以为了更准确的预测土壤呼吸速率,应该对各种复合模型进行对比,从而得到最佳的土壤呼吸速率与土壤温湿度拟合模型。
土壤理化性质和枯落物的蓄积量也是影响土壤呼吸速率的重要因子,这是由于土壤理化性质和枯落物的差异往往会影响植被根系的分布和生长量、土壤微生物的数量及活性,从而影响土壤呼吸速率[29],相关研究认为土壤密度、pH 值、有机质、全N、全P、水解N、有效P等因素都会影响土壤呼吸速率。土壤物理性质越好,越有利于土壤中CO2的传输和扩散,从而提高土壤的呼吸速率;枯落物是土壤呼吸在地上部分最主要的碳源,枯落物分解后一部分以CO2的形式归还到大气中,另外一部分以腐殖质的形式返回到土壤中,不同类型低质林的树种及植物群落不同,造成枯落物数量、质量以及分解速率各不相同,从而间接影响了各类型低质林的土壤呼吸速率[30];土壤pH值通过影响土壤化学反应和土壤酶活性来改变土壤呼吸的速率;土壤有机质作为土壤微生物呼吸作用的碳源,同时也是林木根系生长重要的物质基础,其质量分数直接影响着微生物群落的生物量和活性,并显著影响林木跟系的生长,因此土壤有机质质量分数显著影响着森林土壤呼吸速率[10];土壤氮、磷、钾是植物生长所需的重要营养元素,可以保证林木的生长,加快林木的呼吸作用,从而间接影响土壤呼吸速率。本研究表明,土壤呼吸速率与土壤总孔隙度、有机质含量存在显著的正相关性,这与多数研究结论相同[31-33],另外土壤呼吸速率与土壤pH值、氮质量分数及半分解枯落物蓄积量相关性较高。
本文分析了大兴安岭林区5种不同类型低质林土壤呼吸速率的日变化走势,得到了土壤呼吸速率与温湿度的关系方程,同时分析了枯落物、土壤理化性质对土壤呼吸速率的影响。但由于土壤呼吸速率的时空变化性以及不同因素对土壤呼吸速率的影响都十分复杂,所以大兴安岭地区低质林土壤呼吸变化及不同因子对土壤呼吸的影响机制仍需深入研究。
[1]武传胜,沙丽清,张一平.哀牢山中山湿性常绿阔叶林凋落物对土壤呼吸及其温度敏感性的影响[J].东北林业大学学报,2012,40(6):37-40.
[2]雷海清,傅懋毅,李正才,等.不同海拔高度毛竹林土壤呼吸变化及其影响因子[J].东北林业大学学报,2013,41(2):56-59.
[3]栾军伟,向成华,骆宗诗,等.森林土壤呼吸研究进展[J].应用生态学报,2006,17(12):2451-2456.
[4]孟春,王立海,沈微.小兴安岭针阔混交林择伐6a后林地土壤呼吸速率空间变异性[J].东北林业大学学报,2011,39(3):72-75.
[5]刘颖,韩士杰.长白山典型森林生态系统土壤呼吸的季节动态[J].东北林业大学学报,2012,40(11):107-110.
[6]Concilio A,Ma S,Li Q,et al.Soil respiration response to prescribed burning and thinning in mixed-conifer and hardwood forests[J].Canadian journal of forest research,2005,35(7):1581-1591.
[7]严俊霞,秦作栋,李洪建,等.土高原地区柠条人工林土壤呼吸[J].林业科学,2010,46(3):1-8.
[8]Bajracharya R M,Lal R,Kimble J M.Diurnal and seasonal CO2-C flux from soil as related to Erosion Phases in central Ohio[J].Soil Science Society of America Journal,2000,64(1):286-293.
[9]杨阔,王传宽,焦振.东北东部5种温带森林的春季土壤呼吸[J].生态学报,2010,30(12):3155-3162.
[10]康永祥,夏国威,刘建军,等.秦岭小陇山锐齿栎林皆伐迹地土壤呼吸特征[J].应用生态学报,2014,25(2):342-350.
[11]冯朝阳,吕世海,高吉喜,等.华北山地不同植被类型土壤呼吸特征研究[J].北京林业大学学报,2008,30(2):20-26.
[12]孟春,王立海,王俭,等.5种人工林非主要生长季节土壤呼吸对土壤温度昼夜变化的响应[J].东北林业大学学报,2009,37(11):77-79,91.
[13]杨庆朋,徐明,刘洪升,等.土壤呼吸温度敏感性的影响因素和不确定性[J].生态学报,2011,31(8):2301-2311.
[14]李勇,刘继明,秦世立,等.大兴安岭3种林分夏季土壤呼吸的日变化[J].东北林业大学学报,2011,39(10):65-66,80.
[15]Bååth E,Wallander H.Soil and rhizosphere microorganisms have the same Q10for respiration in a model system[J].Global Change Biology,2003,9(12):1788-1791.
[16]纪浩,董希斌,李芝茹.大兴安岭低质林诱导改造后土壤呼吸影响因子[J].东北林业大学学报,2012,40(4):97-100.
[17]Curiel Yuste J,Janssens I A,Carrara A,et al.Annual Q10of soil respiration reflects plant phenological patterns as well as temperature sensitivity[J].Global Change Biology,2004,10(2):161-169.
[18]Raich J W,Schlesinger W H.The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate[J].Chemical and Physical Meteorology,1992,44(2):81-99.
[19]Davidson E A,Janssens I A,Luo Y Q.On the variability of respiration in terrestrial ecosystems:moving beyond Q10[J].Global Change Biology,2006,12(2):154-164.
[20]于雷,王玉杰,王云琦,等.重庆缙云山针阔混交林地土壤呼吸速率及温度敏感性特征分析[J].环境科学研究,2014,27(1):57-63.
[21]汪金松,范娟,赵秀海,等.太岳山油松人工林土壤呼吸组分及其影响因子[J].林业科学,2013,49(2):1-7.
[22]赵娜,孟平,张劲松,等.华北低丘山地不同土地利用条件下的土壤呼吸比较[J].林业科学,2014,50(2):1-7.
[23]王铭,刘兴土,李秀军,等.松嫩平原西部草甸草原典型植物群落土壤呼吸动态及影响因素[J].应用生态学报,2014,25(1):45-52.
[24]Luo Y Q,Zhou X H.土壤呼吸与环境[M].姜丽芬,曲来叶,周玉梅,等,译.北京:高等教育出版社,2007.
[25]Gulledge J,Schimel J P.Controls on soil carbon dioxide and methane fluxes in a variety of taiga forest stands in interior Alaska[J].Ecosystems,2000,3(3):269-282.
[26]Lavigne M B,Foster R J,Goodine G.Seasonal and annual changes in soil respiration in relation to soil temperature,water potential and trenching[J].Tree Physiology,2004,24(4):415-424.
[27]Rayment M B,Jarvis P G.Temporal and spatial variation of soil CO2efflux in a Canadian boreal forest[J].Soil Biology and Biochemistry,2000,32(1):35-45.
[28]Kang S,Doh S,Lee D,et al.Topographic and climatic controls on soil respiration in six temperate mixed-hardwood forest slopes,Korea[J].Global Change Biology,2003,9(10):1427-1437.
[29]Tripathi S,Kumari S,Chakraborty A,et al.Microbial biomass and its activities in salt-affected coastal soils[J].Biology and fertility of soils,2006,42(3):273-277.
[30]Borken W,Beese F.Soil respiration in pure and mixed stands of European beech and Norway spruce following removal of organic horizons[J].Canadian Journal of Forest Research,2005,35(11):2756-2764.
[31]陈书涛,刘巧辉,胡正华,等.不同土地利用方式下土壤呼吸空间变异的影响因素[J].环境科学,2013,34(3):1017-1025.
[32]Rodeghiero M,Cescatti A.Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps[J].Global Change Biology,2005,11(7):1024-1041.
[33]Merbold L,Ziegler W,Mukelabai M M,et al.Spatial and temporal variation of CO2efflux along a disturbance gradient in a miombo woodland in Western Zambia[J].Biogeosciences Discussions,2010,7(4):5757-5800.
