天花粉蛋白基因导入欧美杨1081)
2014-09-18孙婷婷
孙婷婷 邹 莉
(东北林业大学,哈尔滨,150040)
许继飞
(内蒙古大学)
于 洋
(东北林业大学)
欧美杨 108(Populus euramericana“114/690”)引种于意大利,为美洲黑杨与欧洲黑杨的人工杂种无性系,该品种在耐旱性、耐寒性、生长速度、抗病虫害能力及成材质量等方面都非常优越,被列为我国华北和西北重点推广树种。但是,随着欧美杨的广泛栽培,病虫害的发生越来越严重,因此,其抗性育种显得尤为迫切。采用基因工程技术开展转基因杨树的研究,是培育抗逆、抗虫和抗病杨树新品种的有效途径[1]。天花粉蛋白(Trichosanthin,TCS)是从葫芦科药用植物栝楼(Trichosanthes kirilowii Maxim)块根中分离得到的单链核糖体失活蛋白(Ribosome Inactivating Protein,RIP),近年来的研究结果表明,天花粉蛋白和植物的防御反应相关,它对真菌、病毒和昆虫有直接或间接的抑制和杀灭作用[2]。利用基因工程手段,将具有抗病和抗虫特性的天花粉蛋白基因转入欧美杨108,增强其抗逆性,使欧美杨108免遭病虫危害,具有无污染,不杀伤天敌,保护生物多样性,维持生态平衡等特点,且意义重大。
目前,阔叶树种主要采用根癌农杆菌介导进行遗传转化[3]。本研究以欧美杨108无菌苗为材料,采用农杆菌介导法,进行抗病虫天花粉蛋白(Trichosanthin,TCS)基因的遗传转化研究。抗性体经PCR和PCR-Southern检测,初步证实TCS基因已经整合到欧美杨108的基因组中。
1 材料与方法
植物材料:欧美杨 108(Populus euramericana“114/690”)组培苗由林木遗传育种国家重点实验室(东北林业大学)提供。
菌株和质粒:宿主菌 Escherichia coli DH5α,Agrobacterium tumefeciens LBA4404均为北京大学保藏菌种。pT1-3为北京大学构建含有TCS启动子及结构基因(提取自葫芦科药用植物栝楼(Trichosanthes kirilowii Maxim)块根中)的质粒,其中启动子大小约为 1140 bp,结构基因大小约为 760 bp。pCambia1301为北京大学保藏的质粒。pUCm-T质粒载体购自上海生工公司。
试剂:各种限制性内切酶、质粒提取试剂盒、DNA片段和PCR产物回收试剂盒为Promega公司产品;T4 DNA连接酶和 λDNA/HindⅢ+EcoRⅠMarker为华美生物工程公司产品;Taq DNA聚合酶购自TaKaRa公司,其他常规试剂均为国产分析纯。引物合成和测序由上海生工公司完成。
培养基:分化培养基,MS+0.5 mg·L-16-BA+0.1 mg·L-1NAA;预培养基,MS+0.5 mg·L-16-BA+0.1 mg·L-1NAA;共培养基,MS+0.5 mg·L-16-BA+0.1 mg·L-1NAA+200 μmol·L-1AS(乙酰丁香酮);预筛选培养基,MS+0.5 mg·L-16-BA+0.1 mg·L-1NAA+400 mg·L-1Carb(羧卞青霉素);筛选培养基,MS+0.5mg·L-16-BA+0.1mg·L-1NAA+0.2mg·L-1Hyg(潮霉素)+400 mg·L-1Carb(羧卞青霉素);生根培养基,WPM(木本植物用培养基)+0.4 mg·L-1IBA+0.1 mg·L-1Hyg(潮霉素)+600 mg·L-1Cefo(头孢噻肟钠)。
植物表达载体构建:用限制性内切酶SacⅠ和BamHⅠ双酶切质粒pT1-3得到TCS启动子及结构基因,再回收目的片段;同样使用限制性内切酶SacⅠ和BamHⅠ双酶切质粒pCambia1301后得到载体片段并将其回收,将两者用T4 DNA连接酶16℃过夜连接,连接产物转化大肠杆菌DH5α感受态细胞。经PCR鉴定的阳性菌液,提取质粒,进行SacⅠ和BamHⅠ双酶切鉴定和双向测序。鉴定正确的质粒命名为 pC-tPro-TCS[4]。
农杆菌介导转化欧美杨108:将构建好的植物表达载体用液氨冻融法导入农杆菌中。采用叶盘法转化欧美杨108,从平板上挑取经PCR验证的阳性单菌落放入5 mL含有相应抗生素(50 mg·L-1Rif、40 mg·L-1Strep、50 mg·L-1Kan)的 LB 液体培养基中,28℃,190 rpm振荡培养过夜;取1 mL菌液加入20 mL无抗生素的LB液体培养基中,振荡培养6~8 h,在无菌条件下用无菌水稀释菌液至OD600(光密度)为0.4左右;在无菌条件下,取生长良好的无菌苗叶片,将叶片横斜切成2~3段,叶背向下在黑暗条件下预培养2 d;放入农杆菌侵染液中侵染15 min,在这一过程中轻轻振荡,取出叶片段,用无菌滤纸吸干;将叶片转入共培养基中于培养室中暗培养2d[5]。叶片枯化后进行数据记录,数据用 SPSS和DPS软件分析。
转化效率=(形成抗性芽(或抗性愈伤)的叶片数/总的叶片数)×100%。
抗性植株的分子生物学检测:抗性植株的PCR扩增检测。采用CTAB法[6]提取植物材料总DNA。以质粒DNA作为阳性对照,以未转化的欧美杨108 DNA作为阴性对照,抗性苗进行PCR检测,根据TCS结构基因设计引物为 5'-GGGAAGCTTTGCCATATTGTTTCGATTC-3'和 5'-GGGCATATGGATGTTAGCTTCCGGTTA-3';PCR反应的总体积 25 μL,PCR反应条件为94℃预变性4 min,94℃变性1 min,55℃退火1 min,72℃延伸 1 min,30 个循环,再72℃延伸10 min。扩增产物用1%琼脂糖凝胶电泳检测。
抗性植株的PCR-Southern杂交:采用地高辛标定的探针进行分子杂交。分子杂交试剂盒为DIG High Prime DNA Labeling and Detection Starter KitⅠ,Cat.No.11745832910。将 PCR 扩增产物电泳后印迹于硝酸纤维素膜上,以阳性质粒扩增回收产物为模板合成探针,进行PCR-Southern杂交检测。
2 结果与分析
2.1 植物表达载体pC-tPro-TCS鉴定
质粒pCambia 1301为11.8 kb,在多克隆位点上有SacⅠ和BamHⅠ酶切位点,SacⅠ和BamHⅠ之间有10 bp。质粒pT1-3的SacⅠ与BamHⅠ位点之间约为96 bp,TCS启动子及结构基因长为1.9 kb,所以构建的重组质粒 pC-tPro-TCS大小约为13.7 kb。提取重组质粒pC-tPro-TCS,进行KpnⅠ和BglⅡ双酶切鉴定,得到约2.0 kb和11.7 kb两个片段,所得酶切产物片段大小与预期相符。此外,双向测序结果也表明,克隆的TCS启动子及结构基因序列是正确的。
2.2 植物表达载体转化农杆菌
将构建好的植物表达载体经冻融法转化农杆菌LBA4404,在附加卡那霉素、利福平和链霉素的LB培养基上筛选。挑取阳性克隆,一部分摇菌、提取质粒进行PCR扩增,另一部分进行菌落直接PCR鉴定,两者均特异的扩增出约760 bp大小的TCS结构基因;除此之外,提取质粒进行KpnⅠ和BglⅡ双酶切鉴定,所得酶切产物大小与预期相符,结果表明,已成功地将含有天花粉蛋白(TCS)基因的pC-tPro-TCS植物表达载体导入了根癌农杆菌LBA4404。
2.3 欧美杨108转化抗性再生植株的获得
欧美杨108叶片经农杆菌侵染和共培养后,在含有2 mg·L-1Hyg(潮霉素)的筛选培养基中进行筛选,2个月后,移入生根培养基中诱导生根,获得再生植株(图1、图2)。本研究转化420个外植体,共获得18个抗性愈伤和抗性芽,转化效率为4.3%。
2.4 抗性植株的PCR扩增检测
取抗性植株基因组DNA做模板,以农杆菌质粒DNA作为阳性对照,以未转化植株为阴性对照,用TCS结构基因的特异性引物进行PCR扩增。结果如图3所示,未转化植株的基因组未出现PCR特异扩增片断,抗性植株与质粒DNA的扩增条带相同,大小均为760 bp。因此,初步证明了目的基因TCS已经整合到受体欧美杨108的基因组中。

图1 选择培养基上抗性芽的获得

图2 转基因苗
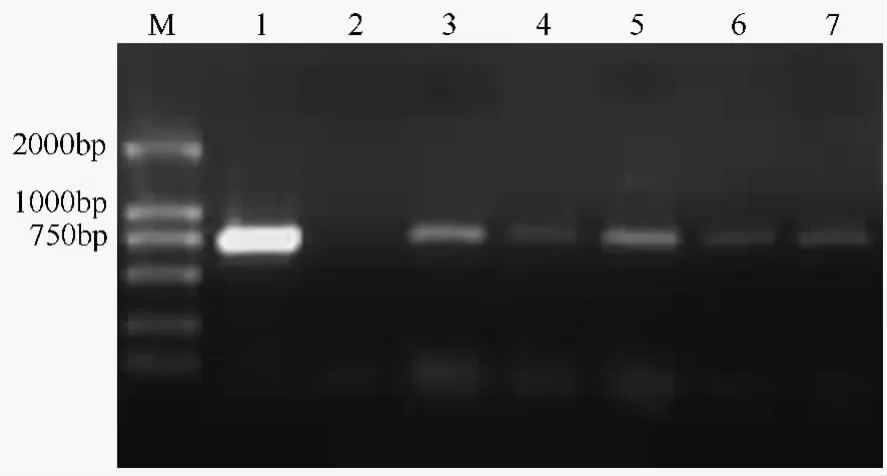
图3 Hyg抗性再生植株的PCR检测电泳分析
2.5 抗性植株的PCR-Southern杂交
为了进一步证明PCR扩增产物是目的基因片段,将PCR扩增产物电泳后印迹于硝酸纤维素膜上,以阳性质粒扩增回收产物为模板合成探针,进行PCR-Southern杂交检测,结果表明,转入TCS基因植株和对照一样,均显示出较强的杂交信号。结果见图4。在检测过的4个抗性植株中,全部呈阳性。进一步证明了TCS基因已整合到了欧美杨108基因组中。
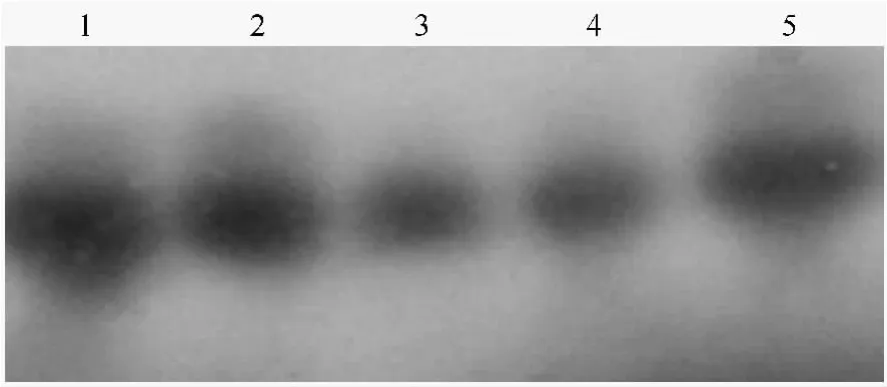
图4 Hyg抗性再生植株的PCR-Southern检测
3 结束语
近年来,天花粉蛋白(Trichosanthin,TCS)基因被转入不同植物中,徐琼芳等[7]将天花粉蛋白基因导入小麦中,发现转天花粉蛋白基因的小麦对黄矮病的抗性得到显著提高。Yan et al.[8]、Ming et al.[9]将天花粉蛋白基因与35S启动子融合,通过改进的基因枪技术,转入到Zhonghua 8中,获得了天花粉蛋白稳定表达的阳性植株,同时,还发现天花粉蛋白作为外源基因的引入并没有对水稻的生长发育产生影响。周长梅[10]采用根癌农杆菌介导法首次将天花粉蛋白基因导入木本植物葡萄中,并获得了PCR阳性植株。2011年刘静等[11]将 TCS基因转化泡桐,通过抗性试验表明,转TCS基因泡桐对丛枝病具有一定的抗性。到目前为止,还未见有利用农杆菌介导法将TCS基因转入欧美杨108的研究报道。
本研究构建了含有TCS启动子和结构基因的植物表达载体,对构建的植物表达载体pC-tPro-TCS的鉴定时,采用的是KpnⅠ和BglⅡ双酶切,而不是SacⅠ和BamHⅠ双酶切鉴定,KpnⅠ和BglⅡ分别位于SacⅠ的上游和BamHⅠ的下游,这样酶切和测序的结果更为准确。将构建好的植物表达载体通过农杆菌介导法成功导入欧美杨108植株中,抗性体经PCR和PCR-Southern检测可以初步证实外源基因整合到基因组中[12-14],而对于TCS基因是否真正整合到欧美杨108的基因组中则需要进一步试验,例如是否有报告基因蛋白表达的染色等[15]。
[1]闫新甫.转基因植物[M].2 版.北京:科学出版社,2003:426.
[2]汪猷.天花粉蛋白[M].北京:科学出版社,1990.
[3]姜金仲,李云,薛诺稳,等.林木遗传转化及其潜在生态风险研究进展[J].西北林学院学报,2012,27(1):103-108,123.
[4]邹莉.天花粉蛋白基因表达载体的构建及转化杨树的研究[D].哈尔滨:东北林业大学,2005.
[5]许继飞.欧美杨108再生体系的优化及转天花粉蛋白基因的研究[D].哈尔滨:东北林业大学,2006.
[6]Mccouch S R,Chen X,Panaud O,et al.Microsatellite marker development,mapping and applications in rice genetics and breeding[J].Plant Molecular Biology,1997,35(1/2):89-99.
[7]徐琼芳,李连城,马有志,等.用天花粉蛋白基因转化小麦获得转基因植株[J].遗传,2001,23(2):135-137.
[8]Yan Yuanxin,An Chengcai,Li Li,et al.The sense and antisense expression of gibberellin 20-oxidase gene(rga5)in rice and its effects on GA1 level and agronomic traits[J].Chinese Science Bulletin,2003,48(5):443-448.
[9]Ming Xiaotian,Yuan Huayi,Wang Lijiang,et al.Agrobacteriummediated transformation of rice with help of bombardment[J].Acta Botanica Sinica,2001,43(1):72-76.
[10]周长梅.农杆菌介导法将天花粉蛋白基因导入葡萄的研究[D].哈尔滨:东北农业大学,2002.
[11]刘静,黄艳艳,翁曼丽,等.TCS基因转化泡桐及抗病能力[J].林业科学,2011,47(5):171-176.
[12]张书利,刘丽君,唐晓飞,等.利用农杆菌介导法将BrCS基因导入大豆的研究[J].大豆科学,2011,30(4):569-573.
[13]金华,尚敏克,姜国斌.导入抗逆基因的转双抗虫基因741杨的获得[J].东北林业大学学报,2012,40(1):28-31.
[14]徐书举,杨晓钦,张仁和,等.不同玉米自交系中导入SAMS基因的转化与检测[J].西北农业学报,2012,21(3):59-62.
[15]梁海永,刘兴菊,苏彦苹,等.RolB基因对三倍体毛白杨的转化研究[J].西北林学院学报,2011,26(2):86-90.
