新生隐球菌感染小鼠肺泡巨噬细胞相关细胞因子与疾病病程相关性研究
2014-09-11雷文知桑军军潘炜华廖万清
雷文知 桑军军 潘炜华 廖万清
(上海长征医院皮肤病与真菌病研究所全军真菌病重点实验室第二军医大学附属长征医院皮肤科,上海 200003)
新生隐球菌是临床上重要的致病真菌之一,它具有显著的嗜神经性,能导致严重的隐球菌性脑膜炎,症状重、死亡率高。其感染途径主要经呼吸道吸入气化担孢子(有性繁殖体)或干燥的酵母细胞而侵入肺部,肺组织是新生隐球菌感染人体后最先受影响的器官,而肺泡巨噬细胞是机体对抗隐球菌感染的第一道防线的重要组成部分。隐球菌被吸入后,首先被吞噬效应因子 (包括巨噬细胞、树突状细胞和嗜中性粒细胞等)识别并吞噬[1],其中单核/巨噬细胞是最主要的效应细胞之一,巨噬细胞和隐球菌的相互作用将决定感染的程度。巨噬细胞一方面通过内吞和氧化杀伤以及分泌胞外抗菌酶等非特异杀伤隐球菌[2],另一方面通过释放细胞因子调节自身及其他免疫细胞的抗真菌作用。巨噬细胞在受到外界病原刺激时,可以向肺组织迁移并释放多种细胞因子 IL-6、IL-8、TGF-β、TNF-α 等,而这些细胞因子也可以反过来趋化中性粒细胞、嗜酸性粒细胞及T淋巴细胞等向感染部位迁移,并对巨噬细胞的功能产生影响。为进一步研究巨噬细胞源性细胞因子在隐球菌致病中所起的作用,本文建立了新生隐球菌吸入感染小鼠模型,并对感染后不同时期小鼠肺泡巨噬细胞上相关细胞因子的表达水平与小鼠肺组织变化进行相关性分析。
1 材料和方法
1.1 动物模型的建立
①实验动物:近交型雌性BALB/c小鼠购于中国科学院,一共144只,体重在16~22 g,随机分为8组,每组18只,对照组为未感染新生隐球菌小鼠,其余7组均为感染组,感染后处死时间分别是1 d、4 d、7 d、11 d、14 d、18 d 和 21 d。②菌种及菌悬液制备:菌种来源于上海长征医院皮肤真菌保藏实验室的标准型H99新生隐球菌菌株,菌株20 mL YPD培养基中培养至饱和态,PBS清洗,1 200 r/min离心3 min,反复3次,无菌双蒸水重悬,菌体计数调定终浓度为5×106CFU/mL,配好的菌悬液仅限当天使用。③小鼠吸入感染新生隐球菌模型:感染组小鼠经水合氯醛麻醉后,迅速以移液器移出20 μL菌悬液经小鼠单侧鼻孔接种感染 (1×105CFU/鼠)(见图1A)。
1.2 肺泡巨噬细胞的分离和纯化
①CO2窒息法处死BALB/c小鼠,酒精消毒后将小鼠置于操作台上仰卧固定,无菌条件下消毒颈部,剪开颈部皮肤,仔细分离气管。②取1 mL注射器抽取37℃ PBS 1.0 mL(含1%EDTA),缓慢推入(约1 min)气管,轻揉肺部,停留1 min后再缓慢回抽,得回收液约0.8 mL(首次肺内残留约0.2 mL,以后各次注入量与回收量相等,即为1.0 mL),重复灌洗10次(见图1B)。③离心肺泡灌洗液收集细胞,用 PBS洗涤3次,用含10%胎牛血清的DMEM培养液8 mL重悬后至25 mL培养瓶中,充分混匀后在37℃、5%CO2、饱和湿度的培养箱内孵育2 h,使巨噬细胞贴壁。④摇晃培养瓶,悬浮未黏附细胞,吸出细胞悬液。PBS反复冲洗,除去未贴壁细胞,加入10 mL 1%EDTA,以使贴壁细胞脱落。30 min后回收贴壁细胞。⑤DMEM培养液洗涤巨噬细胞3次,用含10%胎牛血清的DMEM培养液悬浮细胞,即得纯化后的肺泡巨噬细胞。
1.3 组织病理学检查
经灌洗后的肺组织常规PAS染色,光镜下观察肺组织病理变化、新生隐球菌生长和巨噬细胞吞噬隐球菌状态、炎症细胞浸润情况。
1.4 RT-PCR 检测
常规提取肺泡巨噬细胞RNA后,逆转录为cDNA,用RT-PCR技术检测巨噬细胞源性细胞因子 (IL-6、IL-8、TGF-β、TNF-α)的 mRNA 表达,PCR引物信息详见表1(引物由上海生工有限生物公司合成)。

表1 RT-PCR引物信息表Tab.1 Primer information of RT-PCR
1.5 统计学分析
统计学处理采用SPSS 13.0。各组间的比较采用独立样本t检验,P<0.05为有统计学差异。
2 结 果
2.1 新生隐球菌吸入感染小鼠模型肺组织病理学变化
新生隐球菌吸入感染小鼠模型肺部组织病理学变化见图2。对照组(未感染)小鼠肺组织病理(见图2a,2b);感染后第1天,肺组织病理未见明显感染灶 (见图2c,2d);感染后第4天,可见散在分布的隐球菌 (见图2e,2f);感染后第7天,可见肉芽肿形成(见图2g,2h);为感染后第11天,可见大量炎性细胞浸润,坏死灶 (见图2i,2j);感染后第14天,可见完整的肉芽肿,肉芽肿内见大量隐球菌 (见图2k,2l);感染后第18天,隐球菌分布至全肺(见图2m,2n);感染后第21天,可见肺组织大量坏死,隐球菌弥漫至全肺 (见图2o,2p)。
2.2小鼠肺泡巨噬细胞相关细胞因子mRNA的表达
小鼠肺泡巨噬细胞相关细胞因子mRNA在不同感染时期的表达见表2、图3。TGF-β在巨噬细胞中的表达量在感染隐球菌后第4天开始升高,与未感染小鼠相比具有统计学差异,P<0.05;随着感染时间推移,TGF-β的表达量进一步升高,至第14天出现峰值,至最后一次观察时间,TGF-β的表达量逐渐下降。IL-6在感染后第14天升高与对照组相比有显著差异,之后逐渐下降。IL-8一直处于稳定高表达状态,但与未感染小鼠相比无显著性差异。TNF-α波动较大,在第7天和第14天表达水平降低,第11天、18天和21天表达升高。
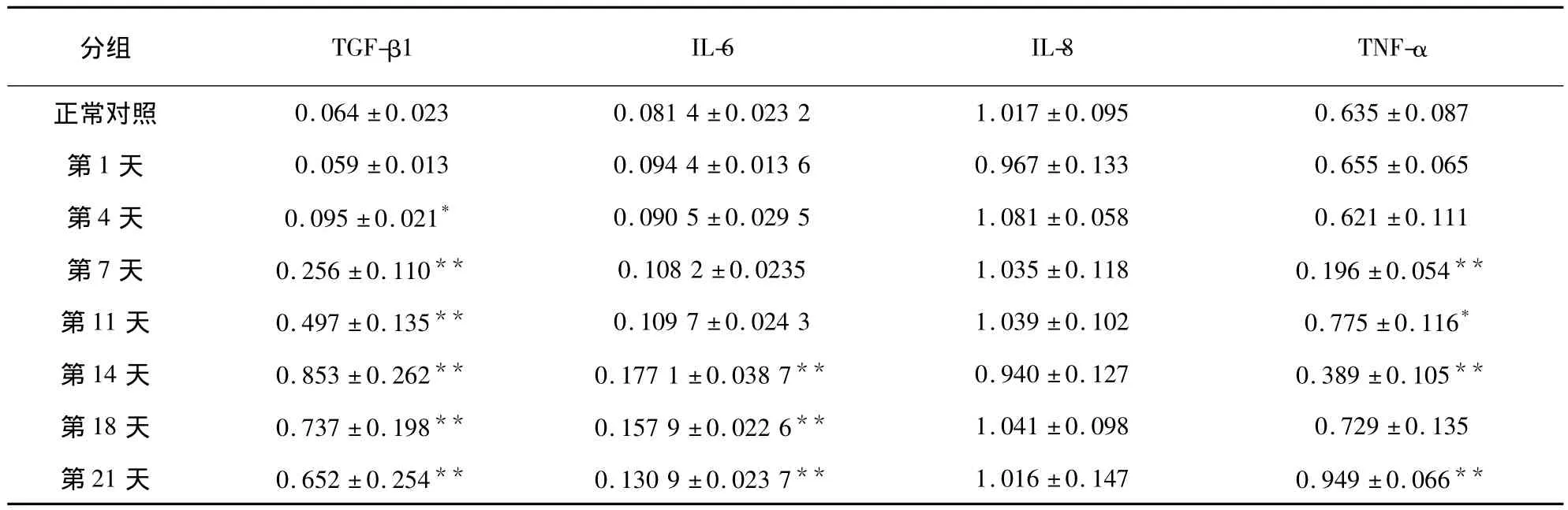
表2 小鼠肺泡巨噬细胞相关细胞因子mRNA在不同感染时期的表达Tab.2 The mRNA expression of mice alveolar macrophage related cytokines at different infection time points
3 讨 论
现今较为成熟的隐球菌感染模型采用的动物有豚鼠、兔、小鼠和大鼠,其中小鼠的近交系特性好,饲养方便,无需免疫抑制处理,所以本实验选择了小鼠建立隐球菌感染模型。根据既往文献报道小鼠感染隐球菌毒性测定,选用感染菌量1×105CFU/鼠,在该菌量侵入条件下,雌性BALB/c小鼠表现出较好的菌株耐受性,未出现急性中毒死亡现象。同时,为了更好的还原人体感染隐球菌的途径,本实验采用气管吸入新生隐球菌的方法建立模型。感染新生隐球菌后小鼠的死亡时间通常为3~4周,本实验选择的观察病程时间点为感染后第1、4、7、11、18、21 天。巨噬细胞是机体抵御隐球菌感染的重要非特异性免疫机制,补体系统以及细胞免疫都可以刺激增强巨噬细胞的吞噬作用起到抗隐球菌感染的作用[3],但隐球菌也同样具备多种毒力因子包括荚膜、黑色素、尿素酶、磷脂酶等能抵御或逃避宿主的免疫,比如巨噬细胞对隐球菌的吞噬作用与隐球菌的荚膜大小呈负相关[4]。研究如何提高巨噬细胞的吞噬和杀伤作用,对有效抵御隐球菌的感染,避免隐球菌向中枢神经系统扩散具有重要意义。
巨噬细胞与新生隐球菌相互作用产生细胞因子是调节宿主免疫的重要方面。IL-6、IL-8、TGF-β、TNF-α等细胞因子是巨噬细胞生产的重要免疫调节因子。本实验发现,在感染后第1天,肺部病理未见明显损害,肺泡灌洗液内可见散在体积较小新生隐球菌,此时RT-PCR检测肺泡巨噬细胞上IL-6、IL-8、TGF-β、TNF-α 的表达与未感染小鼠相比均未产生明显变化。感染后第4天,在小鼠肺组织病理中发现有散在分布的隐球菌,肺泡灌洗液中发现隐球菌体积增大,出芽增多,此时TGF-β的表达水平出现上升。至第7天可见肉芽肿形成,肉芽肿形成是因为随着新生隐球菌菌体的增大,单个巨噬细胞已不能完成对其的吞噬,需要多个巨噬细胞联合发挥抗隐球菌效应,此时TGF-β的水平明显上升,推测其在巨噬细胞早期杀伤隐球菌效应中起促进作用。至感染第11天,肺组织病理可见大量炎性细胞浸润,此时TGF-β的表达水平进一步升高,TGF-β有很强的趋化单核细胞的能力,我们推测炎性细胞的大量浸润与TGF-β的升高有关。到感染第14天,肺组织病理发现许多完整的肉芽肿,肉芽肿内有大量新生隐球菌,此时TGF-β的表达量也达到峰值,之后缓慢下降,考虑TGF-β的下降与此时巨噬细胞形成的肉芽肿不能完全杀灭新生隐球菌,反而为新生隐球菌的生长和繁殖提供更为有利的胞内生长环境有关,更多的隐球菌菌体在巨噬细胞中增殖后逃逸出来,逐步分布至全肺,导致肺组织大量坏死,小鼠濒临死亡。
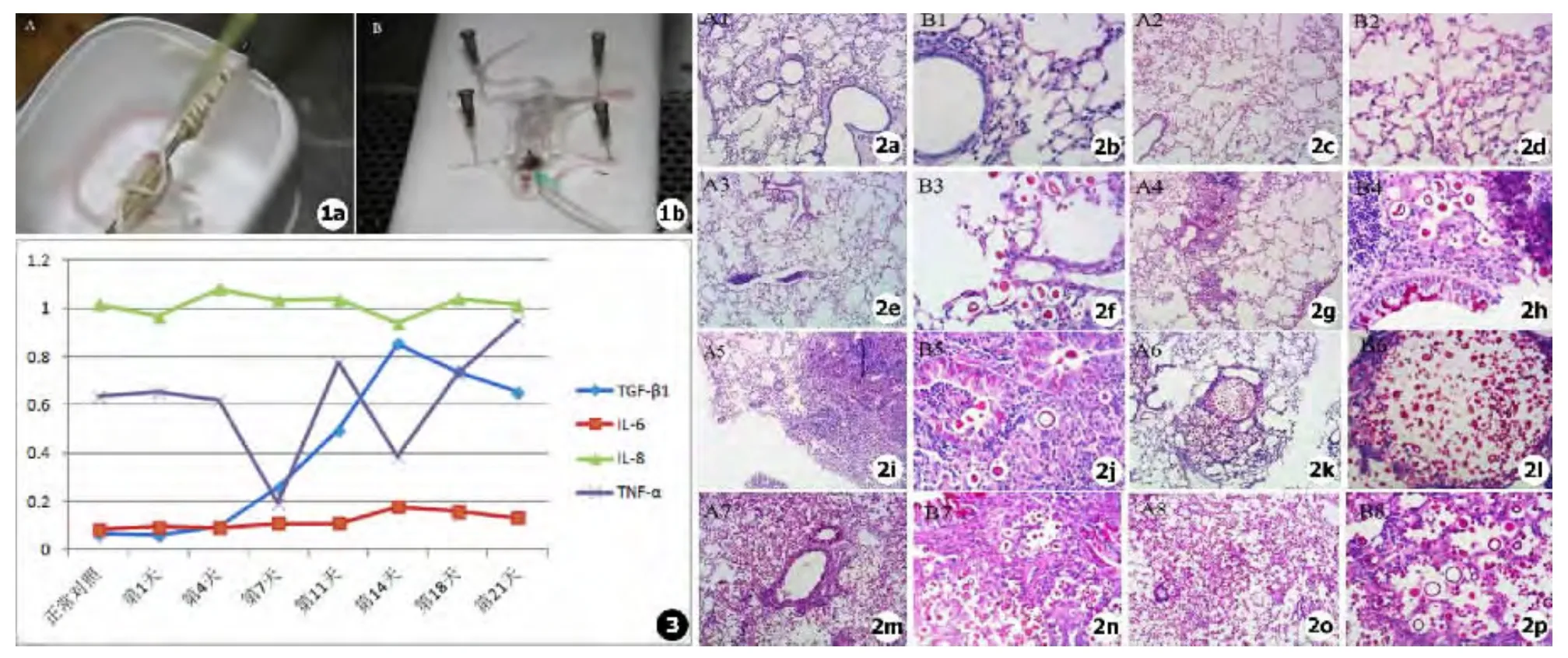
图1 a.小鼠吸入感染新生隐球菌模型,b.肺泡灌洗提取巨噬细胞 图2 小鼠吸入感染H99后,肺组织隐球菌感染病理学变化 (a,c,e,g,i,k,m,o.PAS 染色,×100;b,d,f,h,j,l,n,p.PAS 染色,×400) 图3 小鼠肺泡巨噬细胞上不同感染时期相关细胞因子 mRNA 的表达折线图Fig.1 a.Mouse model of Cryptococcus neoformans inhalation infection,b.Extraction macrophages by alveolar lavage Fig.2 Lung tissue pathology of mouse after Cryptococcus neoformans infection(a,c,e,g,i,k,m,o.× 100,PAS;b,d,f,h,j,l,n,p.× 400,PAS)Fig.3 Broken line graph of mRNA expression of mouse alveolar macrophage related cytokines at different infection time points
IL-6的表达在感染前期呈缓慢上升趋势,到感染后第14天也达到峰值,然后缓慢下降,IL-6是Th17细胞的分化共刺激因子,在隐球菌肺部感染模型中发现其可以通过刺激Th17的分化及分泌发挥保护性抵抗隐球菌感染的作用[5],本实验中IL-6的缓慢升高可能与感染后期刺激细胞免疫逐渐发挥作用有关,但当疾病病程发展到无法控制的状态,肺部满布隐球菌的时候,无论是天然免疫还是细胞免疫的功能都受到了隐球菌毒力效应的抑制。本实验发现TNF-α在巨噬细胞上的表达波动很大,在感染后第7天出现表达明显下降,之后又迅速上升,到第14天又出现下降,再继以上升,尽管有报道称在隐球菌脑膜炎患者中持续升高的TNF-α对生存率有明显促进作用[6],但在本实验中未发现TNF-α与疾病病程有显著相关性,另一项隐球菌感染小鼠模型也没有发现TNF-α 的保护作用[7]。
综上所述,巨噬细胞中TGF-β参与了机体对抗新生隐球菌感染的天然免疫过程并与疾病病程呈相关性,当TGF-β表达量下调时机体的感染状态已无法控制,对于TGF-β的具体功能还需进一步研究,这也为疾病的治疗靶点选择提供了新的方向。
[1]Shao X,Mednick A,Alvarez M,et al.An innate immune system cell is a major determinant of species-related susceptibility differences to fungal pneumonia[J].J Immunol,2005,175(5):3244-3251.
[2]Ie S,Quiniones B,Kovitz K.Cryptococcus neoformans[J].J La State Med Soc,1998,150(12):640-641.
[3]Voelz K,May RC.Cryptococcal interactions with the host immune system[J].Eukaryot Cell,2010,9(6):835-846.
[4]BolanosB, MitchellTG. PhagocytosisofCryptococcusneoformansby rat alveolar macrophages[J].J Med Vet Mycol,1989,27(4):203-217.
[5]Wozniak KL,Kolls JK,Wormley FL Jr.Depletion of neutrophils in a protective model of pulmonary cryptococcosis results in increased IL-17A production by gammadelta T cells[J].BMC Immunol,2012,13:65.
[6]Jarvis JN,Casazza JP,Stone HH,et al.The phenotype of the Cryptococcus-specific CD4+memory T-cell response is associated with disease severity and outcome in HIV-associated cryptococcal meningitis[J].J Infect Dis,2013,207(12):1817-1828.
[7]Wozniak KL,Ravi S,Macias S,et al.Insights into the mechanisms of protective immunity againstCryptococcus neoformansinfection using a mouse model of pulmonary cryptococcosis[J].PLoS One,2009,4(9):e6854.
