碱蓬金属硫蛋白基因对碳酸盐胁迫的应答
2014-08-25金淑梅
金淑梅,孙 丹,王 吉
(东北油田盐碱植被恢复与重建教育部重点实验室 东北林业大学,黑龙江 哈尔滨 150040)
碱蓬金属硫蛋白基因对碳酸盐胁迫的应答
金淑梅,孙 丹,王 吉
(东北油田盐碱植被恢复与重建教育部重点实验室 东北林业大学,黑龙江 哈尔滨 150040)
从碱蓬(Suaedaglauca)中分离了一个Ⅱ型金属硫蛋白基因(SgMT),该基因读码框为234 bp,编码78个氨基酸。利用qRT-PCR发现SgMT基因在种子和叶中高度表达。在不同浓度的Na2CO3和NaHCO3盐胁迫条件下,碱蓬种子中SgMT基因的表达量也在逐渐增大。不同pH值对SgMT基因的表达量无显著影响。将SgMT导入酵母细胞,在添加12 mmol·L-1Na2CO3和26 mmol·L-1NaHCO3的酵母培养基中,含有转SgMT基因的酵母生长明显好于对照酵母的生长。在不同pH值的酵母培养基上,转SgMT基因和未转基因酵母的生长没有明显的区别。这些结果说明在碳酸盐胁迫过程中,金属硫蛋白基因在植物提高环境逆境胁迫过程中起着一定的作用。
碱蓬;金属硫蛋白;盐胁迫;pH值
碱蓬(Suaedaglauca)为碱蓬属一年生草本,作为改良盐碱地的“先锋植物”[1],具有很强的耐盐碱能力,分布在松嫩平原上的碱蓬能在碳酸盐含量较高的碱斑上正常生长,在重度盐碱地区上形成的大面积碱蓬群落是土壤严重碱化的标志[2]。
以NaCl和Na2SO4为主的盐化土壤和以Na2CO3、NaHCO3为主的碱化土壤总称为盐碱土壤[3]。黑龙江省松嫩平原的草地上存在不同程度以Na2CO3和NaHCO3为主的碱性盐碱土[4]。盐碱对植物种子萌发具有毒害作用,白三叶(Trifoliumrepens)种子对NaCl、Na2CO3的耐受浓度阈值分别为300和100 mmol·L-1[5]。碱茅(Puccinelliadistans)种子在340 mmol·L-1NaCl胁迫下不能发芽[6]。关于碱蓬属种子萌发也有相关报道,翅碱蓬(S.pterantha)种子能耐受NaCl胁迫,但当NaCl溶液的浓度达到600 mmol·L-1时,种子几乎不发芽[7]。在800 mmol·L-1的NaCl溶液中,囊果碱蓬(S.physophora)只有极少数种子萌发[8],目前,有关盐碱胁迫对碱蓬影响的报道较少,当用100 mmol·L-1Na2CO3或300 mmol·L-1NaHCO3溶液处理土壤时,碱蓬种子萌发会被显著抑制[9-10]。利用CaCl2等外源物质浸种,能提高盐地碱蓬种子的发芽率和发芽速度[11]。魏磊等[12]利用400 mmol·L-1NaCl或200 mmol·L-1NaHCO3处理碱蓬幼苗,处理组的净光合速率值与未加盐处理相比分别减少了73.7%和76.0%。
金属硫蛋白(Metallothioneins,MTs)最初是从马的肾脏中提取出来的一类富含半胱氨酸[13]、能结合重金属离子的低分子量蛋白质[14]。东北油田盐碱植被恢复与重建实验室从碱蓬植株中克隆出一个金属硫蛋白基因,发现该基因与盐碱逆境有一定关系,为了检测碱蓬金属硫蛋白基因在种子萌发时与盐碱胁迫的应答关系,选取了Na2CO3和NaHCO3碳酸盐逆境、不同酸碱度对SgMT基因在转录水平上的表达特性进行了研究,并利用酵母细胞,进一步研究了SgMT基因与盐碱的关系。有关盐碱和酸碱度胁迫对碱蓬种子萌发时金属硫蛋白表达的研究还未见报道。在分子水平上,植物抵御盐碱胁迫有多种可能的机制[15],碱蓬是高度耐盐碱植物,其金属硫蛋白基因在碱蓬抗逆性中是否发挥作用呢?目前对金属硫蛋白基因研究主要集中在与金属的关系上[16],缺少对金属硫蛋白基因在NaHCO3和Na2CO3等碱性盐胁迫下表达的研究,而黑龙江省多数为含有一定碱性盐的苏打盐碱土,因此,研究金属硫蛋白基因在碱性盐胁迫的表达更有意义。鉴于此,本研究主要对在不同盐逆境胁迫下的金属硫蛋白基因的表达特性进行研究,以期为阐明SgMT基因提高植物盐碱抗性提供理论依据。
1 材料和方法
1.1材料
碱蓬植株和种子采自黑龙江省安达市重度盐碱化地区。
1.2方法
1.2.1基因的RT-PCR克隆 根据GenBank上EST片段(登录号:HS411207)查询核苷酸序列,通过同源性比对,与海蓬子(Salicorniaeuropaea)的Ⅱ型金属硫蛋白序列(JF780913)的近3’端片断存在83%的相似性,根据海蓬子金属硫蛋白基因的核苷酸序列,设计RT-PCR引物,SgMTF:5′-ATGTCTTGCTGTGGTGGT-3′和SgMTR:5′-TCATTTGCAGTGCAAGGG-3′。采用Trizol试剂盒(Invitrogen)提取新鲜碱蓬植株总RNA;按照M-MLV cDNA合成试剂盒(宝生物)的操作说明,把总RNA反转录成cDNA,以其为扩增模板,PCR反应条件:95 ℃ 5 min,95 ℃ 30 s,50 ℃ 30 s,72 ℃ 2 min,35个循环,72 ℃延伸10 min;PCR产物片段回收、连接T载体后测序。
1.2.2SgMT基因在碱蓬各个器官中的表达情况 采集新鲜碱蓬根、茎、叶、花、种子,用Trizol法提取各器官的RNA,反转录成cDNA,根据碱蓬金属硫蛋白基因和碱蓬肌动蛋白Actin(EU429457)序列[17]设计引物。序列如下:
SgMTF:5′-ATGTCTTGCTGTGGTGGT-3′
SgMTR:5′-TCATTTGCAGTGCAAGGG-3′
ActinF:5′-GTGGTCGTACAACAGTA-3′
ActinR:5′-GACCCTCCAATCCAGACA-3′
利用SYBR Green Ⅰ试剂盒(Tiangen)进行实时荧光定量qRT-PCR检测,每个样品都做3个平行试验。经优化后的PCR反应体系为20.0 μL(2×Sybr Green mix 10.0 μL;10 μmol·L-1上、下游引物各0.5 μL;cDNA模板1.0 μL,ddH2O 8.0 μL);反应条件为:94 ℃预变性5 min,94 ℃ 15 s,60 ℃ 1 min,40个循环。每个样品进行3次独立重复实验。根据2-△△Ct的方法计算SgMT基因的表达水平。得到的各器官中碱蓬金属硫蛋白表达量,利用SPSS软件中的方差分析进行各组间的比较。
1.2.3SgMT基因在逆境处理后的碱蓬种子中的表达 每个处理分别选取30 粒种子,3次重复,在Na2CO3(50、100、150、200 mmol·L-1)、NaHCO3(100、200、300、400 mmol·L-1)、酸碱度(pH值6、7、8、9、10)逆境处理3 d后,提取种子RNA。种子总RNA反转录为cDNA,利用qRT-PCR分析SgMT基因在逆境处理后的碱蓬种子中的表达情况,最后得到的相对表达量利用SPSS软件进行方差分析。
1.2.4SgMT基因在酵母细胞中的表达 酵母重组表达载体的构建:为构建SgMT基因的酵母表达载体,选用了在酿酒酵母(Saccharomycescerevisiae)中能诱导表达重组蛋白的pYES2载体,该载体中的半乳糖激酶启动子(GAL1)能够在酿酒酵母中被半乳糖高水平地诱导目的蛋白表达,同时能够被葡萄糖抑制表达。首先设计在基因读码框上游添加BamHⅠ和下游添加XhoⅠ酶切位点的特异性引物,PCR获得增设酶切位点的SgMT基因读码框片段,用BamHⅠ/XhoⅠ双酶切pMD18-T-(BamHⅠ)SgMT(XhoⅠ)连接T载体的质粒,同时用BamHⅠ/XhoⅠ双酶切pYES2质粒DNA,相应地回收SgMT基因目的片段和pYES2载体,经T4连接酶连接两者,测序正确后命名为pYES2-SgMT。pYES2(对照)和pYES2-SgMT(过量表达SgMT基因)两种质粒分别转化酵母细胞INScⅠ中,用于抗性分析。
在酵母中的抗性分析:挑取含有pYES2和pYES2-SgMT的酵母单克隆于YPD(Yeast Extract 10 g·L-1、Peptone 20 g·L-1、Glucose 20 g·L-1)液体培养基中,30 ℃培养过夜。次日用分光光度计分别测菌液的OD600值,调整菌液OD600值为0.8,取样至离心管,离心弃上清后,加入灭菌水500 μL。将菌液用H2O分别稀释至10-1,10-2,10-3,10-4,10-55个浓度。每个浓度吸取5 μL菌液分别点在未处理的,和含12 mmol·L-1Na2CO3、26 mmol·L-1NaHCO3(通过梯度试验后选取的最佳浓度)及不同酸碱度(pH值6、7、8、9、10)的YPG(Yeast Extract 10 g·L-1、Peptone 20 g·L-1、Galactose 20 g·L-1)固体培养基上,观察两种酵母的生长情况。30 ℃培养2~3 d后进行观测照相。
2 结果与分析
2.1SgMT基因的克隆
将提取碱蓬植株的总RNA反转录成cDNA,以cDNA为模板,利用特异设计的引物进行PCR扩增反应,电泳后在200 bp左右有明显的条带,回收PCR产物连接到pMD18-T载体上,命名为pMD18-T-SgMT,SgMT基因编码区测序为234 bp,利用Primer premier 5软件,发现编码78个氨基酸。通过NCBI氨基酸序列同源性比对,结果显示碱蓬金属硫蛋白(SgMT)与海蓬子Ⅱ型金属硫蛋白(SbMT ofSalicorniabrachiata)(JF780913)有很高的相似度,其同源性达到了83%(图1)。在数据库中得到的其他相关蛋白是巴拉圭茶(IpMT ofIlexparaguariensis)(AFP93964.1)和花生的Ⅱ型金属硫蛋白(AhMT ofArachishypogaea)(ABA08415.1),与他们的氨基酸序列都有80%的同源性。同时得到碱蓬金属硫蛋白的氨基酸序列与二色补血草Ⅱ型金属硫蛋白(LbMT ofLimoniumbicolor)(ABL10085.1)和蝇子草Ⅱ型金属硫蛋白(SnMT ofSileneniceensis)(ADP92404.1)分别达到了78%和75%(图1)。

图1 碱蓬金属硫蛋白的同源性比对Fig.1 Homology alignment analysis of SgMT
2.2SgMT基因在碱蓬各个器官中的表达特性
SgMT基因在碱蓬不同器官中的表达情况为,在种子中表达量最高,其次是叶、茎、花、根,且种子和叶与其他器官间表达量差异显著(P<0.05)(图2)。
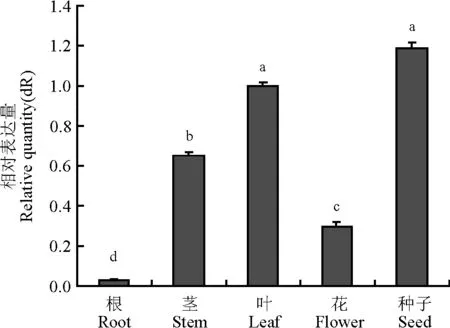
图2 不同碱蓬器官中金属硫蛋白表达的定量比较Fig.2 Quantitative comparison of MT gene expression in Suaeda glauca organs by qRT-PCR
2.3不同盐胁迫和pH值下SgMT基因表达特性
碱蓬种子金属硫蛋白基因SgMT在Na2CO3、NaHCO3和不同pH逆境中均受到了不同程度的诱导,在未处理条件下,SgMT基因的表达量最低,基因表达量随着各种逆境浓度的增大而增大。金属硫蛋白的表达量在碱性盐各个处理浓度下比未处理的条件下都明显增加(图3A,3B)。不同pH值处理和未处理的条件下基因的表达增加不显著(P>0.05)(图3C)。
2.4SgMT基因在酵母中表达及抗逆性
为了解析SgMT基因在酵母中是否具有抗逆性,以转pYES2的酵母菌株为对照,对转pYES2-SgMT的酵母在逆境条件下的生长进行观察(图4),在不做处理的培养基上(CK),转pYES2-SgMT与转pYES2的酵母的生长相似。但在12 mmol·L-1Na2CO3和26 mmol·L-1NaHCO3盐胁迫下,转pYES2-SgMT的酵母菌株生长明显好于转pYES2的酵母菌株。说明携带SgMT基因的酵母菌株在盐胁迫下表达提高,增强INVSc1菌株(pYES2-SgMT)耐盐能力。在不同pH逆境胁迫下,转pYES2-SgMT与转pYES2的酵母菌株的生长没有明显的区别。

图3 不同Na2CO3, NaHCO3, pH值浓度下碱蓬种子中金属硫蛋白表达的定量比较Fig.3 Quantitative comparison of MT gene expression in Suaeda glauca seed under Na2CO3, NaHCO3, and pH value stress by qRT-PCR

图4 过量表达pYES2-SgMT基因酵母抗逆性分析Fig.4 Resistance analysis of yeast overexpressed pYES2-SgMT recombinant plasmid
3 讨论
3.1SgMT基因的克隆
Ⅱ型金属硫蛋白,半胱氨酸排列一般为Cys-Cys,而且Cys-Cys位于氨基酸序列的第3、4位,碱蓬中克隆得到的金属硫蛋白蛋白序列为MSCCGGNCGCGAGCKCGNGCGGCKMFPDFSESTSASTELISGVAPQRSYMGGSEMGVAAENDGCKCGDNCQCNPCTCK,第3、4位就是两个半胱氨酸,并且通过同源序列比对,与其他植物的Ⅱ型金属硫蛋白同源性比较大,因此断定试验中克隆的基因为碱蓬Ⅱ型金属硫蛋白基因。
3.2SgMT基因表达的组织特异性
运用灵敏度很高的qRT-PCR检测不同处理的SgMT基因表达情况,每个处理中3次重复的检测结果都很接近,说明用qRT-PCR检测该基因表达差异的可信度是很高的。本研究中的基因在叶中的表达量明显高于根部表达量,这与相关研究中Ⅱ型金属硫蛋白基因主要在植物叶片中的表达量较高,在根部表达量较少的结论相符[18-19]。盐碱是影响种子萌发的显著因素,盐碱环境下的种子萌发又是盐生植物生长的关键和敏感阶段[20],SgMT基因在种子中的表达量最大,有可能帮助植物缓解在种子萌发期受到的盐碱胁迫伤害,提高植物在逆境下的发芽率。
3.3盐胁迫对碱蓬种子中SgMT基因表达特性的影响
研究结果表明,SgMT基因在转录水平上分别不同程度地受到了碱性盐NaHCO3、Na2CO3逆境的诱导。高pH值对碱蓬种子中基因的表达没有显著影响。由此推测SgMT基因对碱性盐胁迫有一定的应激反应,这种反应不是由高pH值所引起的。
3.4过量表达碱蓬SgMT基因的酵母在盐胁迫下的表达特性
酵母和植物都属于真核生物,代谢途径比较相似,对翻译的外源蛋白质能够进行折叠加工和修饰。酵母比植物生长周期短,因此试验周期快,本研究通过过量表达碱蓬SgMT基因的酵母细胞在碱性盐和不用酸碱度胁迫条件下的生长状况,来验证基因的功能。转SgMT基因酵母在添加12 mmol·L-1Na2CO3和26 mmol·L-1NaHCO3的酵母培养基上的生长明显好于未转SgMT基因酵母,表明SgMT基因在酵母体内表达后,减缓了逆境对组织和细胞伤害作用,从而提高了酵母在逆境条件下的生长。在不同pH值的培养基中,转SgMT基因酵母与未转SgMT基因的酵母生长状态没有明显的区别,这说明SgMT基因不能通过强化细胞内pH调控能力来提高抗盐碱能力。SgMT基因具有良好的抗盐碱能力是否由于SgMT基因的表达量增加,产生不同的金属硫蛋白基因调控链,带动其他逆境胁迫应答基因的表达,从而提高转SgMT基因酵母的抗逆性,尚待更深入的研究。
本研究对碱性盐逆境下碱蓬种子中金属硫蛋白基因的表达进行了研究,并利用酵母表达体系进一步观察碱蓬金属硫蛋白基因的表达,初步揭示碱蓬高抗盐碱性的分子机制,对充分利用碱蓬的抗逆基因资源和植物分子育种提供更多帮助。
[1]张立宾,徐化凌,赵庚星.碱蓬的耐盐能力及其对滨海盐渍土的改良效果[J].土壤,2007,39(2):310-313.
[2]魏磊.盐碱胁迫碱蓬幼苗的蛋白质组学研究[D].哈尔滨:东北林业大学,2012.
[3]黎立群.盐渍土基础知识[M].北京:科学出版社,1986.
[4]柳参奎,张欣欣,金洙哲.中国东北盐碱地植物原色图鉴[M].哈尔滨:东北林业大学出版社,2006.
[5]卢艳敏,苏长青,李会芬.不同盐胁迫对白三叶种子萌发及幼苗生长的影响[J].草业学报,2013,22(4):123-129.
[6]张海南,周青平,颜红波,梁国玲,刘文辉.盐胁迫对5种碱茅材料种子萌发的影响[J].草业科学,2013,30(11):1767-1770.
[7]刘兴.NaCl溶液对翅碱蓬种子萌发的影响[J].农业科技与装备,2012(11):1-5.
[8]王雷,田长彦,张道远,周智彬.光照、温度和盐分对囊果碱蓬种子萌发的影响[J].干旱区地理,2005,28(5):670-674.
[9]代莉慧,蔡禄,周耀龙,贾晋.NaCl和Na2CO3胁迫对内蒙古河套灌区盐地碱蓬种子萌芽生理指标的影响[J].种子,2013,32(7):14-17.
[10]赵楠,芦艳,左进城,鲁周民.碱胁迫对碱蓬种子萌发的影响[J].北方园艺,2012,36(1):45-47.
[11]何丽丹,刘广明,杨劲松,吕真真,李金彪.外源物质浸种对NaCl胁迫下盐地碱蓬发芽的影响[J].草业科学,2013,30(6):860-867.
[12]魏磊,庞秋颖,张爱琴,国静,阎秀峰.盐碱胁迫对碱蓬幼苗光合特性的影响[J].东北林业大学学报,2012,40(1):32-35.
[13]Margoshes M,Vallee B L.A cadmium protein from equine imaging hyperintensity in Alzheimer’s disease:Correlation with kidney cortex[J].Journal of the American Chemical Society,1957,79:2.
[14]全先庆,张洪涛,单雷,毕玉平.植物金属硫蛋白及其重金属解毒机制研究进展[J].遗传,2006,28(3):375-382.
[15]Tester M,Davenport R.Na+tolerance and Na+transport in higher plants[J].Annals of Botany,2003,91:503-527.
[16]Hsieh H M,Liu W K,Huang P.A novel stress-inducible metallothionein-like gene from rice[J].Plant Molecular Biology,1995,28(3):381-389.
[17]马清,周向睿,伍国强,王锁民.盐生植物碱蓬Actin基因片段的克隆及序列分析[J].生物技术,2009,19(1):1-3.
[18]Zhou J,Goldsbrough P B.Functional homologs of fungal metallothionein genes fromArabidopsis[J].The Plant Cell Online,1994,6(6):875-884.
[19]Huang G Y,Wang Y S.Expression and characterization analysis of type 2 metallothionein from grey mangrove species (Avicenniamarina) in response to metal stress[J].Aquatic Toxicology,2010,99:86-92.
[20]Khan M A,Sheith K H.Effect s of different levels of salinity on seed germination and growth ofCapsicumAnnuum[J].Biologia,1996,22:15-16.
(责任编辑 张瑾)
2014年10月国内市场主要畜产品与饲料价格分析
10月国内主要畜产品和饲料价格波动中呈现下降态势。
一、猪肉、鸡肉和鸡蛋价格下降,牛肉和羊肉价格稳中有升
10月国内猪肉、鸡肉和鸡蛋平均价格分别为19.40、15.05和10.86元·kg-1,环比分别下降2.8%、5.2%和1.6%。从区域分析,猪肉价格东部分别高于中、西部0.1%和0.1%;鸡肉价格东部分别高于西、中部2.4%和7.7%;鸡蛋价格西部分别高于中、东部2.6%和3.8%。牛肉平均价格54.47元·kg-1,未有明显波动,东部分别高于中、西部0.3%和0.4%。羊肉平均价格53.29元·kg-1,环比上涨0.2%,东部分别高于中、西部2.8%和4.3%。
二、玉米、大豆、豆粕和棉粕价格不同幅度下降
10月国内玉米、大豆、豆粕和棉粕平均价格为2 510、4 263、3 688和2 433元·t-1,环比分别下降6.2%、1.7%、0.3%和14.3%。从区域分析,玉米价格东部分别高于西、中部4.7%和4.9%;大豆价格东部分别高于西、中部1.5%和2.9%;豆粕价格西部分别高于中、东部3.3%和8.4%;棉粕价格东部分别高于中、西部0.6%和3.9%。
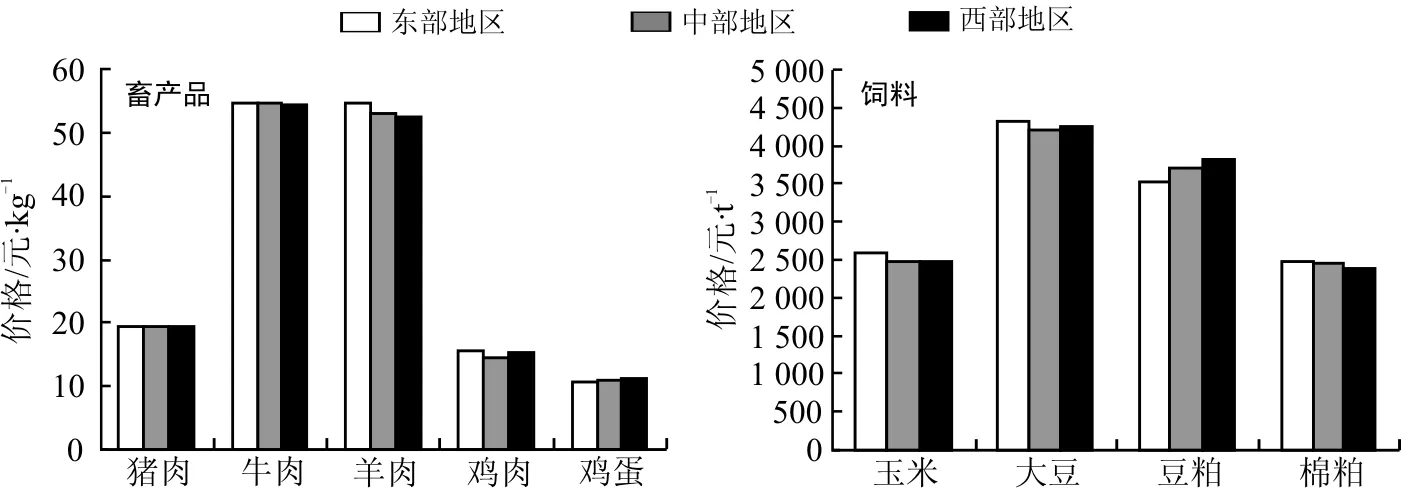
图1 2014年10月国内市场主要畜产品与饲料价格
数据来源:猪肉http://www.chinapig.cn/index.htm;牛肉、羊肉、鸡肉、鸡蛋http://www.21food.cn/;玉米、豆粕http://www.zhue.com.cn/;大豆、棉粕http://www.chinafeed.org.cn/
(兰州大学草地农业科技学院 王化 整理)
Metallothioneingene(SgMT)fromSuaedaglaucaexpressionundersalinestress
JIN Shu-mei, SUN Dan, WANG Ji
(Key Laboratory of Saline-alkali Vegetation Ecology Restoration in Oil Field (SAVER), Ministry of Education, Northeast Forestry University, Harbin 150040, China)
A type Ⅱ metallothionein gene (SgMT) gene was isolated fromSuaedaglaucawhich had 234 bp open reading frame and encoded 78 amino acids.SgMTwas highly expressed in seeds and leaves which were tested by qRT-PCR.SgMTgenes up-regulated under different concentrations of Na2CO3and NaHCO3salt stress and did not change under high pH conditions.The recombinant yeast cells withSgMTgrew better than control cells under 12 mmol·L-1Na2CO3and 26 mmol·L-1NaHCO3salt stress and had no difference under different pH value.These results suggested thatSgMTgene played an important role in plant adaptation to environmental stresses.
Suaedaglauca; metallothionein; saline stress; pH value
JIN Shu-mei E-mail:jinshumei1972@163.com
2014-04-09 接受日期:2014-07-15
哈尔滨市青年基金(2008RFQXN007);国家自然科学基金 (31070616);黑龙江省自然科学基金(C200714);黑龙江省自然基金(留学及海外基金LC2013C10)
金淑梅(1972-),女,黑龙江明水人,副教授,博士,主要从事生物化学与分子生物学方向研究。 E-mail:jinshumei1972@163.com
Q945.78
:A
:1001-0629(2014)11-20820-06
10.11829j.issn.1001-0629.2014-0161
