复合性状转基因水稻B2A68在南京地区杂草化的风险评估
2014-08-13戴伟民肖国樱宋小玲
黄 鹞,王 建,戴伟民,强 胜,肖国樱,宋小玲
(1.南京农业大学杂草研究室,江苏南京 210095;2.中国科学院亚热带农业生态研究所,湖南长沙 410125)
复合性状转基因作物指同一植株中含有2个或2个以上的转基因性状或转化体的作物[1]。复合性状转基因作物具有明显的优势,在拥有更广阔的农艺性状以适应非适宜环境的同时,还可以提高产量[2-3]。在所有复合性状中,抗除草剂性状与抗虫性状是主要的复合性状,成为将来转基因作物发展的一个重要方向。
植物杂草化(weediness)是指原本自然分布的栽培植物,在新的人工生境中能自然繁殖其种群而转变为杂草的演化过程。转基因作物由于插入抗虫、抗病、抗除草剂等基因,具有了更强的适应性,在生长势、越冬性、种子产量和种子越冬能力等方面可能比非转基因植株强,入侵性提高,从而占据其他植物的栖息地,影响生物多样性,破坏自然种群平衡。尽管从目前的研究来看,单一性状的转基因水稻在生存竞争能力、自生苗以及越冬能力等方面与非转基因受体以及当地常规水稻相比无明显区别,且杂草化的风险较小[4]。但由于复合性状的转基因作物转入了2个外源基因,外源基因引起的转基因作物生存竞争能力的改变可能比单一性状的转基因更为复杂。
现有的报道都是针对单一性状转基因作物的生存竞争能力,如宋小玲等对孟山都公司的转基因大豆进行了杂草化研究,发现抗草甘膦转基因大豆在江苏省南京地区演化为杂草的可能性较小[5];崔荣荣等对抗草铵膦转基因水稻明恢86B进行了杂草化评估,发现明恢86B演化为杂草的可能性不大[4];陈小文等比较了转Cyr1Ac基因的抗虫玉米及其受体在荒地中的生存竞争情况,发现该转基因玉米品种演化为超级杂草的可能性很小[6];张兴华等对转双价抗虫棉进行了生存能力竞争研究,发现抗虫棉的生存竞争能力强于其非转基因受体[7]。但鲜有复合性状转基因作物的相关报道。
本研究以转Cry2Aa#和转Bar基因的抗虫抗除草剂转基因水稻B2A68为供试材料[8],探索复合性状转基因水稻的杂草化潜力,以期为该复合性状转基因水稻的安全性评估提供科学依据。
1 材料与方法
1.1 材料
转基因水稻B2A68:由湖南杂交水稻研究中心提供,该水稻携带单拷贝的抗虫基因Cry2Aa#与抗除草剂草铵膦的Bar基因,受体水稻为早稻D68。
转基因水稻Bar68-1:转入抗除草剂草铵膦Bar基因,其亲本受体同上,由香港中文大学、湖南杂交水稻研究中心与中国科学院亚热带农业生态研究所合作培育。
当地常规稻:宁粳46,粳型。
1.2 方法
1.2.1 试验基地概况 试验在南京农业大学杂草研究室转基因水稻安全性评估试验基地进行。土壤pH值大于6小于7,偏酸性,肥力良好、排灌情况良好。试验地栽培管理按当地常规栽培方式播种后,不加以其他管理措施。试验期间无影响整个试验结果的恶劣气候因素,试验地设专人看管,设围拦以防止人为或动物破坏。需要收获的转基因水稻材料单独收获、单脱、单藏,由专业技术人员专人运输和保管。检测试验完毕后,除需要保留的材料外,剩余的转基因试验材料一律烧毁。试验地四周有 2 m 高的围栏,100 m范围内无水稻种植,符合国家相关规定。
1.2.2 转基因水稻生存竞争力试验
1.2.2.1 试验设计 处理1:直播。采用3种种植密度即高密度(300粒/m2)、中密度(150粒/m2)、低密度(75粒/m2),小区面积为4 m2(2 m×2 m)。分4期播种,适宜期2期,分别为2012年6月19日(Ⅰ)和2012年6月29日(Ⅱ);非适宜期2期,分别为2012年7月9日(Ⅲ)和2012年7月20日(Ⅳ)。每处理4次重复,采用随机区组排列。
处理2:移栽。用25~30 d秧苗单株移栽,株距20 cm,行距20 cm。小区面积6.8 m2(2 m×3.4 m),各处理随机排列。
1.2.2.2 调查内容 杂草调查:采用对角线5点取样法,直播方式播种的样方面积为0.1 m2,移栽方式的样方面积为1 m2。在播种后30、60 d调查记录每小区杂草种类、株数,按杂草垂直投影面积占小区面积的比例估算出杂草的相对覆盖率。
竞争力调查:直播方式在播种后10 d、移栽方式在水稻移栽后15 d调查记录每小区的水稻株数及成苗率,以后每隔15 d每小区随机调查10株长势最好的水稻,测量其主茎株高、分蘖数。
1.2.3 转基因水稻繁殖能力
1.2.3.1 试验设计 本试验的田间试验设计同“1.2.2”部分,在生存竞争力试验田中进行。
1.2.3.2 调查内容 花粉活力检测:每小区随机取10株水稻,每株水稻取3朵小花。采用常规I2-KI染色法进行检测。
生殖生长指标统计:水稻成熟后每小区采收10株长势最好的水稻,统计每株主穗的穗长、每穗饱粒数、每穗总粒数、落粒数、地上部生物量,并计算出结实率(结实率=每穗饱粒数/每穗总粒数×100%)和落粒率,落粒率的检测及计算参照崔荣荣等的方法[4],落粒率水平参照应存山的方法[9]。
1.2.4 自生苗和再生苗 在生存竞争力试验的同一块田中进行。除繁殖力考察中每小区采收10株水稻外,其余水稻均不收割,让其自然落在土壤中,田块冬季闲置,不进行任何管理。在收完需考察的水稻后,每隔20 d调查一次试验小区内的自生苗和再生苗情况,共调查2次,并在翌年当地水稻分蘖期后调查一次自生苗和再生苗。通过考察转基因水稻作物演化成自生苗的比例,评价转基因水稻的自然延续能力。对出现的自生苗取样后带回实验室验证,调查结束后翻耕。
1.2.5 种子生存能力
1.2.5.1 种子萌发试验 水稻种子收回后30 d按GB/T 3543.4—1995《农作物种子检验规程 发芽试验》规定的方法检测发芽率。
1.2.5.2 种子埋藏试验 种子埋藏试验于2013年1月20日进行,按随机区组试验设计,设浅埋(3 cm)和深埋(20 cm)2种埋藏深度。取待测种子25粒装于200目的尼龙网袋中,4次重复,埋藏地点在水稻生存竞争力试验田,并分别于埋后2、4、6个月取出种子进行种子萌发试验,并统计已腐烂数。不能萌发的种子用TTC法检测种子是否有活力。
2 结果与分析
2.1 杂草调查结果
直播稻田播后30 d时第Ⅰ期和第Ⅱ期杂草发生量很大,主要是异型莎草(CyperusdifformisLinn.)、稗草[Echinochloacrusgalli(L.) Beauv.]、水苋菜(AmmanniabacciferaLinn.),占杂草发生量的65%,其次还有少量的鸭舌草[Monochoriavaginalis(Burm. f.)C. Presl]、四叶萍[MarsileaquadrifoliaL.(E)pepper Wort]、水虱草[Fimbristylismiliaceae(L.)Vahl.]、节节菜[Rotalaindica(Willd.) Koehne]。同一水稻品种,高密度播种方式下的杂草量略低于低密度下的杂草量,但总体上来说,B2A68、Bar68-1和常规稻的小区杂草数量没有显著差异。第Ⅰ期、第Ⅱ期总草覆盖率在45%~65%。播后 60 d 时,千金子大量发生,占总草的50%以上,千金子与水稻的高度比约为1.6 ∶1,总草覆盖率达到80%。
直播第Ⅲ期和第Ⅳ期,各小区杂草发生量差异不显著,播后30 d杂草发生量较第Ⅰ期和第Ⅱ期少,主要杂草为千金子和异型莎草,占总草的44%左右,其次还有少量的水虱草、节节菜等,总草覆盖率为56%~60%。播后60 d时,千金子成为主要的杂草,占总草的60%以上,总草覆盖率约为70%。
移栽水稻田移栽后30 d杂草与直播水稻相比发生量不大,主要杂草为异型莎草,覆盖率约为20%,下层有少量的空心莲子草[Alternantheraphiloxeroides(Mart.) Griseb.]和节节菜。
2.2 转基因水稻生存竞争力
2.2.1 出苗率 由表1可知,在第Ⅰ期,相同水稻在不同密度下的出苗率无显著差异,相同密度下不同水稻的出苗率也无显著差异。
在第Ⅱ期,B2A68低密度种植出苗率显著低于中密度,但与高密度处理下的出苗率无显著差异;Bar68-1在不同密度处理下的出苗率均没有显著差异;而常规稻3种种植密度之间的出苗率存在很大差异,其中中密度的出苗率显著高于高密度、低密度。在相同密度处理下,中密度时3种水稻的出苗率无显著差异,高密度、低密度时常规稻的出苗率显著低于2种转基因水稻。
第Ⅲ期和第Ⅳ期的情况与第Ⅰ期相同,即相同水稻在不同密度下的出苗率无显著差异,相同密度下不同水稻的出苗率也无显著差异。种植密度和播种时间对同一水稻品种的出苗率影响不大。
从出苗率的整体情况考察,B2A68没有表现出比Bar68-1更高的出苗率,2种转基因水稻的出苗率相当,多在70%以上;与常规稻相比,供试的2种转基因水稻在大多数情况下没有表现出更高的出苗率,但在第Ⅱ期高密度、低密度时2种转基因水稻的出苗率显著高于常规水稻的出苗率。
2.2.2 株高 直播第Ⅰ、Ⅱ、Ⅲ、Ⅳ期的株高见图1。25~70 d的株高调查结果显示,同一密度处理下B2A68和Bar68-1多高于常规稻,且2种转基因水稻之间的株高差异不显著。但是从4期的最终株高来看,3种水稻的株高差异不明显。研究结果还表明,相同水稻在不同种植密度下的株高没有显著差异,说明种植密度对株高影响不大。从数值上相比,第Ⅳ期的3种水稻的最终株高均低于前3期的相同品种的水稻,而前3期水稻的最终株高没有显著差异。
在移栽条件下,移栽后25~40 d,B2A68和Bar68-1的株高显著高于常规稻,但2种转基因水稻间的株高差异不显著;55 d至最终,常规稻株高显著高于2种转基因水稻(图2),说明在草压较小的情况下,2种转基因水稻的长势比常规稻弱。

表1 直播方式种植的3种水稻播种后10 d的出苗率(平均数±标准误)

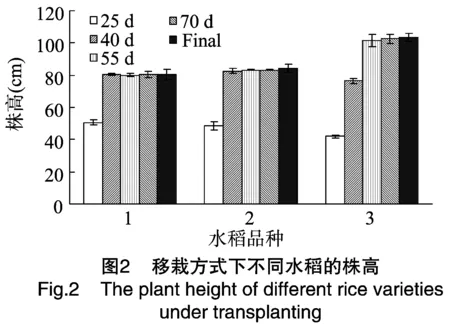
2.2.3 分蘖 由图3可见,第Ⅰ、Ⅱ期,在播后 25~55 d,由于杂草竞争条件的存在,3种水稻的分蘖数都呈现出先增后减的趋势。相同水稻在不同密度处理下分蘖数差异不大,相同密度处理下不同水稻的分蘖数差异也不大。第Ⅲ、第Ⅳ期由于处在水稻生长发育的非适宜期,各种水稻的分蘖数都很少。第Ⅳ期不同处理下的3种水稻分蘖数均为0。
在移栽条件下,3种水稻的分蘖数没有显著差别(图3)。移栽后70 d调查的结果显示,3种水稻的分蘖数量均在9~11个之间。
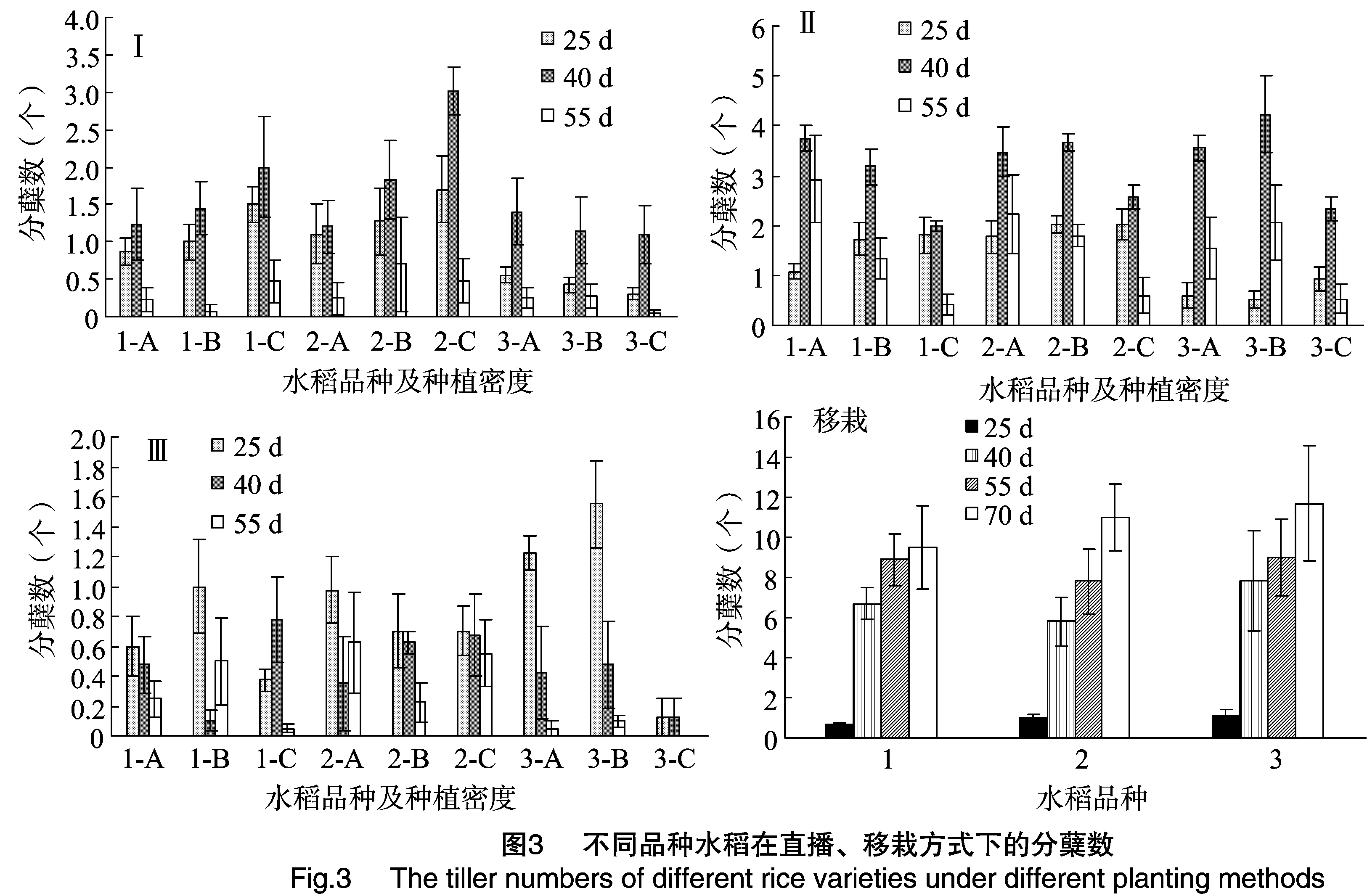
从分蘖数量上分析,在直播适宜期和非适宜期,在相同密度处理以及移栽条件下,3种水稻的分蘖数没有显著差异。
2.3 繁殖力调查
2.3.1 花粉活力 通过观察和统计,3种水稻花粉活力在97%~99%之间,无显著差异,花粉活力相当。
2.3.2 生育期比较 通过观察和统计,3种水稻在4个播期下都能够完成生活史,B2A68和Bar68-1的花期基本一致,常规稻的花期比2种抗性水稻晚:第Ⅰ、Ⅱ期晚10~14 d,第Ⅲ、Ⅳ期晚6~10 d(表2)。
2.3.3 生殖指标分析 由表3可知,从整体来看,相同水稻在不同密度下的生殖指标没有太大差异。而在相同密度处理下,直播适宜期和非适宜期的2种转基因水稻所考量的穗长和每穗饱粒数多显著高于常规稻,2种转基因水稻之间差异不显著;2种转基因水稻与常规水稻相比,结实率和地上部生物量无显著差异。从数值上看,适宜期的各项指标(除结实率)整体上比非适宜的要高,说明B2A68和Bar68-1在不良气候条件下生长竞争能力变弱。与当地常规稻相比,在直播条件下,无论是适宜期还是非适宜期,2种转基因水稻的生殖指标(除结实率)都要高于常规水稻。就生殖生长指标分析,2种转基因水稻没有显著差异,说明转基因水稻没有因为导入了2种外源基因而生长竞争力就变强。

表2 B2A68、Bar68-1、常规稻在4种播期下的花期(2012)
由表4可知,常规水稻的穗长显著小于2种转基因水稻,但每穗饱粒数显著大于2种转基因水稻,2种转基因水稻之间无显著差异;常规水稻和转基因水稻的结实率和地上部生物量无显著差异。这说明在生殖能力方面2种转基因水稻弱于常规水稻。

表3 供试水稻在直播条件下的生殖指标

表4 供试水稻在移栽条件下生殖指标
2.3.4 落粒性 B2A68的落粒率为13.33%,Bar68-1的落粒率为12.80%,常规稻的落粒率为7.02%。在外力条件下,2种转基因水稻的落粒率显著高于常规稻。但参照应存山的报道[9],3种水稻的落粒性都处于中等水平。
2.3.5 自生苗情况 在收回需要考种的水稻之后对田间的自生苗进行调查,调查发现各小区中均无自生苗出现。
2.3.6 种子生存能力
2.3.6.1 种子萌发率 水稻种子收获后30 d,从其萌发情况(图4、图5)来看,不论在直播还是移栽条件下,2种转基因水稻的萌发情况一致,7 d时的萌发率均在80%以上。不同处理下的常规稻3 d时萌发率有所差异,但是7 d时萌发率接近100%。2种转基因水稻的萌发相对不齐,全部萌发所需时间更长,到11 d时仍有少数种子不能萌发。常规稻没有休眠的种子,而2种转基因水稻未能萌发的种子用TTC检测后发现只有不到0.01%的种子保持活力,处于休眠状态。
2.3.6.1 种子活力保存能力 埋藏前,3种水稻种子活力均在90%以上,但是经过埋藏后,不论是浅埋还是深埋,3种水稻种子活力都有一定程度的下降。其中经过2个月浅埋后的B2A68和Bar68-1的种子活力迅速降低到20%左右,深埋处理下2种转基因水稻种子活力迅速降低到13%左右;而常规稻在浅埋和深埋处理下2个月种子活力没有迅速降低,均保持在80%以上。4~6个月,3种水稻种子活力下降得相对缓慢。2~6个月的浅埋和深埋处理下,2种转基因水稻的种子活力无显著差异,但显著低于常规稻,说明2种转基因水稻种子在土壤中保持活力的能力弱于常规稻(图6、图7)。
3 结论与讨论
杂草是能够在人工生境中自然繁衍其种族的植物,最大特点是适应性、持续性、危害性[10]。当一个或多个外源基因导入水稻后有可能提高水稻的生存竞争能力,而竞争力是判断植物是否具有杂草性的主要因子之一。植物的竞争力强弱受到一系列影响因子的影响,如种子萌发特性、株高、生物量等[11]。本试验中供试的复合性状转基因水稻B2A68在出苗率上与单一性状转基因水稻Bar-68是一致的,并没有表现出更高的出苗率。而在有相似杂草竞争的环境下,B2A68与Bar-68在株高、分蘖、花粉活力、穗长、主穗饱粒数、结实率、落粒性、土壤中种子活力保存能力等方面都没有显著差异,说明B2A68并没有因为导入2种外源基因而与单抗水稻Bar-68产生显著不同,没有表现出更强的竞争能力,演化为杂草的可能性也没有因此变大。2种转基因水稻与当地种植的常规稻宁粳46相比,在相似的草压下都没有更强的侵略性。本试验结果表明,以早籼稻D68为受体的转入抗虫抗除草剂的复合性状转基因抗水稻B2A68与单一性状转基因水稻Bar-68在整个生活史中竞争能力相似,没有明显的杂草化倾向,而且与当地常规稻相比,复合性状转基因水稻B2A68生存竞争优势也不突出,该水稻在南京地区演化为杂草的可能性不大。




休眠性和落粒性是评价作物杂草化潜力较为重要的2个特点,休眠性可以让杂草避开非适宜期,这一特征与植物成为入侵能力强的杂草密切相关[12]。由于外源基因插入到水稻基因组的位置是随机的,所以造成的影响是复杂和未知的。水稻休眠是由多个数量性状位点(QTL)所控制,这样的位点在水稻染色体上都有分布,在第12条上分布得最为广泛[13-16]。外源基因的插入有可能造成水稻休眠性增强或降低,但是目前尚没有这类报道,本试验中的转基因水稻没有因为插入外源基因而使水稻休眠性发生明显的变化。落粒性是杂草的一种繁衍种族的机制,也是作物与杂草最大的区别之一。在水稻中迄今已定位5个落粒基因(sh-1、sh-2、sh-3、sh-4 和sh-h),与落粒相关的主效QTL也研究得很多[17]。在评价转基因作物的杂草化中,落粒性是一项重要的指标,本研究中2种外源基因与1种外源基因导入水稻后对水稻落粒性的影响并没有显著差异,说明本试验水稻外源抗虫抗除草剂基因并没有触发水稻的落粒性。
目前复合性状应用较多的植物是玉米和棉花,但是水稻是世界上主要的粮食作物之一,针对复合性状的转基因水稻的杂草化环境风险制定一份标准是十分必要的。虽然复合性状业已成为发展最快的基因技术之一,但是种植了复合性状转基因作物的国家和地区对其监管的要求、内容却各不一样。如美国、加拿大、巴西依据单个转化事件管理复合性状转基因作物。欧盟对复合性状转基因作物的管理较为严格,评价时主要考虑插入基因的完整性和表型的稳定性及复合性状转基因之间可能的相互作用,从选择合适的参照物、成分的比较分析、毒性、过敏性和营养价值评价以及环境评价等方面评价其相互作用带来的影响[1,18-20]。但我国尚未出台复合性状转基因作物杂草化环境风险的评价标准。在转基因技术快速发展的今天,需要加强对复合性状转基因作物的研究和管理。本试验的开展可以为复合性状转基因水稻安全性评估标准的制订提供一定的试验依据。本试验比较的是复合性状和单一性状转基因水稻的杂草化风险,但是为了完整地制定管理标准,还需在试验中设置非转基因受体水稻空白对照进行研究。
参考文献:
[1]刘培磊,李 宁,程金根. 不同国家和地区复合性状转基因作物安全评价管理的比较[J]. 农业科技管理,2008,27(3):23-26.
[2]Halpin C. Gene stacking in transgenic plants-the challenge for 21st century plant biotechnology[J]. Plant Biotechnology Journal,2005,3(2):141-155.
[3]ISAAA. Pocket K No. 42:Stacked traits in biotech crops[EB/OL]. [2014-02-03]. http://www.isaaa.org/resources/publications/pocketk/42/.
[4]崔荣荣,韦 颖,孟攀潘,等. 抗草铵膦转基因水稻明恢86B杂草化潜力评价[J]. 中国水稻科学,2012,26(4):467-475.
[5]宋小玲,强 胜,彭于发. 抗草甘膦转基因大豆[Glycinemac(L.) Merri]杂草性评价的实验实例[J]. 中国农业科学,2009,42(1):145-153.
[6]陈小文,李吉崇,郭玉海,等. 抗虫转基因玉米荒地生存竞争力评价[J]. 杂草科学,2012,30(1):31-34.
[7]张兴华,田绍仁,张天玉,等. 转双价双Bt基因(Cry1Ac+Cry2Ab)抗虫棉栽培地生存竞争能力研究[J]. 现代农业科技,2012(3):78-79,84.
[8]翁绿水,蒋利平,肖国樱. 抗虫抗除草剂转基因水稻恢复系B2A68的培育[J]. 杂交水稻,2013,28(1):63-67.
[9]应存山. 中国稻种资源[M]. 北京:中国农业科学技术出版社,1993:164-185,530-533.
[10]强 胜. 杂草学[M]. 北京:中国农业出版社,2001:1-2.
[11]杜 峰,梁宗锁,胡莉娟. 植物竞争研究综述[J]. 生态学杂志,2004,23(4):157-163.
[12]李 博. 植物竞争:作物与杂草相互作用的实验研究[M]. 北京:高等教育出版社,2001:3-4.
[13]卢丙越. 水稻品种N22强休眠性的QTL定位及遗传解析[D]. 南京:南京农业大学,2011.
[14]江 玲,张文伟,翟虎渠,等. 水稻种子休眠性基因座的定位和分析[J]. 中国农业科学,2005,38(4):650-656.
[15]Ye H,Foleyb M E,Gua X You. New seed dormancy loci detected from weedy rice-derived advanced populations with major QTL alleles removed from the background[J]. Plant Science,2010,179(6):612-619.
[16]Guo L B,Zhu L H,Xu Y B,et al. QTL analysis of seed dormancy in rice (OryzasativaL.)[J]. Euphytica,2004,140:155-162.
[17]杨绍华,杨 玲,李建君,等. 水稻落粒性的研究进展[J]. 天津农业科学,2011,17(3):112-114.
[18]Davies H,Sweet J. Risk assessment of plants containing genetic modification events combined by crossing[EB/OL]. (2006-06-08)[2013-12-10]. http://www.efsa.europa.eu.
[19]Datta K,Baisakh N,Thet K M,et al. Pyramiding transgenes for multiple resistance in rice against bacterial blight,yellow stem borer and sheath blight[J]. Theor Appl Genet,2002,106(1):1-8.
[20]EFSA. Guidance document of the scientific panel on genetically modified organisms for the risk assessment of genetically modified plants containing stacked transformation events[EB/OL]. (2007-05-16)[2013-12-10]. http://www.efsa.europa.eu.
