亚热带中山常绿阔叶林木本植物幼苗数量动态及其与生境的相关性
2014-08-10施璐璐骆争荣夏家天赵文娟吴友贵丁炳扬
施璐璐, 骆争荣,夏家天,赵文娟, 吴友贵, 丁炳扬,*
(1. 温州大学生命与环境科学学院, 温州 325035;2. 丽水学院生态学院, 丽水 323000;3. 凤阳山-百山祖国家级自然保护区百山祖管理处, 庆元 323800)
亚热带中山常绿阔叶林木本植物幼苗数量动态及其与生境的相关性
施璐璐1, 骆争荣2,夏家天3,赵文娟1, 吴友贵3, 丁炳扬1,*
(1. 温州大学生命与环境科学学院, 温州 325035;2. 丽水学院生态学院, 丽水 323000;3. 凤阳山-百山祖国家级自然保护区百山祖管理处, 庆元 323800)
以百山祖自然保护区5 hm2永久样地150个幼苗监测站木本植物幼苗为研究对象,2008—2011年定期调查样方中幼苗的种类、数量、萌发、死亡等,探究亚热带中山常绿阔叶林幼苗种类组成、数量动态及其与生境的相关性。结果表明:1)百山祖样地在2008年至2011年出现的幼苗属于26科,40属,共53个物种,不同物种萌发时段有异;2)2009年样地幼苗存活比率为7.7%,2010年为-20.8%,2011年则是-0.3%,幼苗存活比率不高,种类和数量呈减少趋势;3)存活幼苗中有明显的优势物种,分别为光亮山矾(Symplocoslucida)、尖连蕊茶(Camelliacuspidata)、浙闽新木姜子(Neolitseaauratavar.undulatula)、尖叶菝葜(Smilaxarisanensis)和短尾柯(Lithocarpusbrevicaudatus),5个物种之和占幼苗总数比例>50%;4)种子的萌发与生境有极显著的相关性,且与生境因子中水分关联最大;5)存活幼苗数与样站坡位、水分、落叶层厚度呈现显著相关性,水和光照是影响幼苗存活的主要因素。
百山祖;幼苗;种类组成;数量动态;生境
种子萌发是植物生长周期的开始,有性生殖的实生苗和营养繁殖的萌生苗是高等植物种群天然更新的实现方式,它们构成了植物种群或群落的幼苗库[1]。在植物生活史中,幼苗建成过程对于植物定居和种群建立来说均是最为脆弱的阶段,也是对生境条件最为敏感的时期[2- 3]。幼苗在不同生境下的生活力和生长速率影响着种群的数量、更新速率[4]以及群落的组成情况,左右着种群和群落的发展,对种群或群落的稳定和更新意义重大[5]。到目前为止,有关森林群落天然更新及木本幼苗的研究涉及很多方面,如种子大小与幼苗生长策略[6],幼苗年际动态[7],不同幼苗光合作用比较[8]等。种子萌发对环境条件的要求反映了该物种自身遗传特性及适应环境的能力[9],因此环境异质性对幼苗的影响也是研究的一大热点,尤其是光环境[10- 11]。彭闪江等提出在不同生境中,光照条件、土壤水分和化学成分等因子的组合严重影响种子和幼苗的命运[12],丁圣彦等也提到光照是林下幼苗更新、存活和生长的重要限制因子[13],此外叶片凋落物也能通过叶片的物理障碍或机械损伤,以及对微气候、营养循环的影响和化学他感作用等来影响种子和幼苗的存活[14],这些研究结果对森林群落的天然更新研究以及植被损坏后的再修复意义重大。
亚热带常绿阔叶林生物多样性丰富,对环境保护、维持全球碳循环的平衡和人类的持续发展等具有重要作用[15]。我国亚热带常绿阔叶林的分布区域达到250万km2,但由于长期受人为干扰,几乎丧失殆尽[16]。因此,对常绿阔叶林的保护是十分重要的,而幼苗期作为维持森林稳态的关键阶段,具有极大的研究和保护意义。百山祖位于浙江南部庆元县境内,其低山地带的森林植被为典型常绿阔叶林,但由于人为因素的干扰,受到严重损害,百山祖北坡的中山区域(1300—1700 m) 是仅存的长久未受明显人为干扰的较大面积的常绿阔叶林,是目前我国亚热带地区罕见的保存完好的中亚热带天然植被,具有重要的保护和研究价值[17]。王伟等通过Torus-translation检测方法分析了百山祖样地中DBH≥1 cm、个体数大于5的89种木本植物与4个小生境的关系。结果表明,生境异质性对亚热带常绿阔叶林中木本植物的生物多样性的维持起着重要作用, 同时大部分物种与生境的相关性会随着生长阶段的变化而发生变化[18]。事实上,树木对生境的适应大多在幼苗更新和定居阶段就已发生,但是该研究并未将对生境更加敏感的幼苗阶段纳入分析。
本文利用百山祖自然保护区2003年建立的5 hm2永久性样地,以2008—2011年木本植物幼苗为调查对象进行研究。主要探讨:1)样地幼苗种类组成及萌发时间分布;2)样地幼苗萌发和死亡的动态; 3)样地幼苗数量动态与生境的相关性。研究结果可为百山祖常绿阔叶林群落的天然更新与物种多样性维持研究积累基础资料,也可为生物多样性的保护提供参考。
1 研究地自然概况
百山祖是“浙江凤阳山—百山祖国家级自然保护区”的重要组成部分,植物多样性高,特有珍稀物种多[19]。百山祖自然保护区位于浙江省庆元县境内,以百山祖为中心的一片山地,地理位置119°7′45″—119°19′20″E, 27°37′45″—27°50′30″N,其主峰百山祖海拔为1856.7 m,是浙江省第2高峰。地貌类型为深切割中山,以侵蚀地貌为主,基岩为侏罗纪火成岩,海拔800 m以下为红壤,800 m 以上以黄壤为主,棕黄壤集中分布在海拔1700 m的百山祖南坡[20]。本地区地带性气候为中亚热带季风气候,由于海拔较高,与庆元县城相比具有山地立体气候特征,年均温低,降水充沛,相对湿度大,雾日多。据百山祖保护站气象观测点(海拔1100 m)的观测资料显示保护区年均温12.8 ℃,年降水量2341.8 mm,相对湿度84%,极端高温32 ℃,极端低温-13 ℃[19]。
2 研究方法
2.1 幼苗样方设置
在百山祖5 hm2监测样地内设置了50 个种子雨和幼苗监测样站,其在样地中的位置见图1。以种子雨收集器为中心,避开靠小道的一面,在种子雨收集器的三面距离2 m处,分别设置1个1 m×1 m的小样方,共150个幼苗小样方(图2),每个小样方的4个角用PVC管标出。
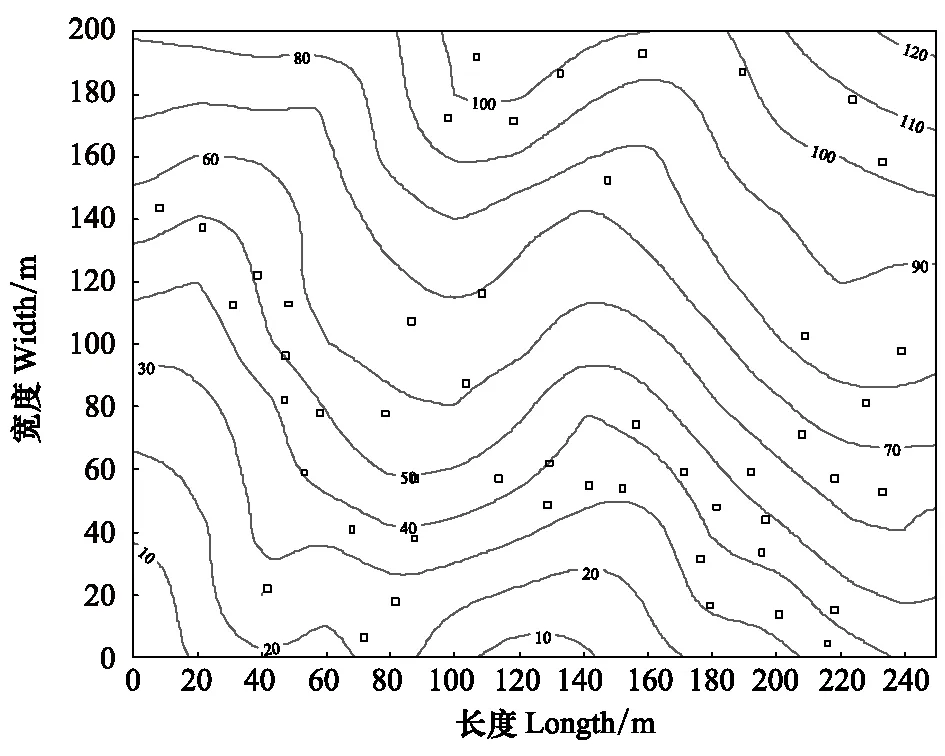
图1 幼苗样站在样地内的分布图Fig.1 Distribution of seedling quadrats in the plot
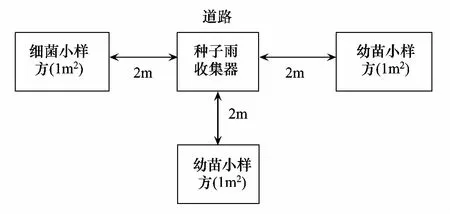
图2 幼苗监测样站内小样方设置示意图Fig.2 The set of seedling polts in seedling monitoring station
2.2 幼苗的调查
于2008年10月进行第1次调查,后每年调查3次,于5月下旬或6月上旬,8月上中旬,10下旬进行。对小样方内所有的木本植物幼苗作逐株调查,并套上塑料脚环进行编号。调查内容包括幼苗的种类、生长状况(出生、死亡)、高度、分枝数、叶数、坐标(在方格绘图纸上标定)等。本文选用2008年10月到2011年10月间的调查数据进行研究。
2.3 数据处理
若在5月份调查时发现新生的幼苗,则视为该物种的种子能在春季萌发,同理,8月份发现视为夏季萌发,10月份为秋季萌发,3次调查均有发现,则视为春夏秋季萌发。
本研究将多个物种的幼苗出生和死亡一并统计。为了区别种群研究中常用的出生率和死亡率,本研究使用出生比率和死亡比率作为表征幼苗群落动态的参数,计算公式如下:
出生比率=本次调查新生幼苗株数/上次调查幼苗总株数
死亡比率=本次调查死亡幼苗株数/上次调查幼苗总株数
幼苗存活比率=出生比率-死亡比率
在探究幼苗与生境的相关性时,由于样地地形变化比较复杂,在监测站中小地形的变化比较大,有时同一监测站的3个幼苗小样方很不一样,为更详细的探究生境各因素与幼苗动态的关系,将生境分解为5个因素:1)坡位(0山谷、1近山谷、2坡面、3近山脊、4山脊);2)坡度(0平坦≤10°,1缓坡10°—25°,2斜坡26°—40°,3陡坡>40°);3)土壤质地(0壤土,以粉粒为主、手感细软;1砂土,以砂粒为主、较粗糙;2多砾石,以石砾为主、很少土壤;3多岩石,大石块或裸露岩石);4)土壤水份(0较干,手抓不能成团;1一般,手抓成团放开能散;2较潮湿,手抓成团放掉不能散;3常年渗水,地表常年渗水);5)落叶层厚薄(0裸露,裸露或四分之一以上面积裸露,1较薄,四分之三以上面积覆盖落叶但厚度不超过1 cm,2中等,整个面积覆盖且厚度超过1 cm但不过超过2 cm,3较厚,厚度超过2 cm)。利用SPSS软件分别将小样方内的幼苗总数(指调查时小样方内的幼苗数总和,下同)、新生数、死亡数与这5个因素进行偏相关性分析(Parial)。用相关系数r来描述两个变量间的密切程度和相关方向,正r值表示正相关,负r值表示负相关,r值绝对值越大,表示相关度越大。
3 结果与分析
3.1 幼苗物种组成及主要萌发时间段
从2008年10月至2011年10月,百山祖样地幼苗监测样站的10次调查数据统计结果见表1。
由表1可得,从2008年10月第1次调查到2011年10月第10次调查在百山祖样地幼苗监测样站共发现53个物种的幼苗。其中秀丽槭、树参、尖叶菝葜等11个树种在1a的3次调查中均有新生幼苗,在春夏秋皆可萌发;扁枝越桔、光亮山矾、黄山木兰等5个物种在5月和8月的调查中发现新生幼苗,其主要萌发季节为春季和夏季;短尾柯、多脉青冈、褐叶青冈等5个树种在8月和10月有新生幼苗,萌发季节为夏季和秋季;黄山松、华东山柳、阔叶槭等13个物种仅在5月份的调查中有新生幼苗,其主要萌发时段为春季;团花山矾、石灰花楸、紫果冬青等8个树种仅在8月份调查中有新生幼苗,主要萌发季节为夏季;猴头杜鹃仅在10月份调查中发现新生幼苗,萌发季节为秋季;而茵芋、东方古柯、厚皮香等10个物种的幼苗在第1次调查时已存在,其后3年未发现新的幼苗,无法判断主要萌发时段。
百山祖样地监测样站内幼苗包括26科,40属,共53个物种,其中43个物种在调查期间有新生幼苗。2009年有34个物种发现新生幼苗,包含21科,28属;2010年新生幼苗包括15科,19属,共21个物种;而2011年新生的幼苗种数为18种,包括9科,16属。在3a间,新生幼苗无论是科属数还是种数都有减少的趋势,而且幼苗种类与百山祖丰富的树种相比明显偏少。
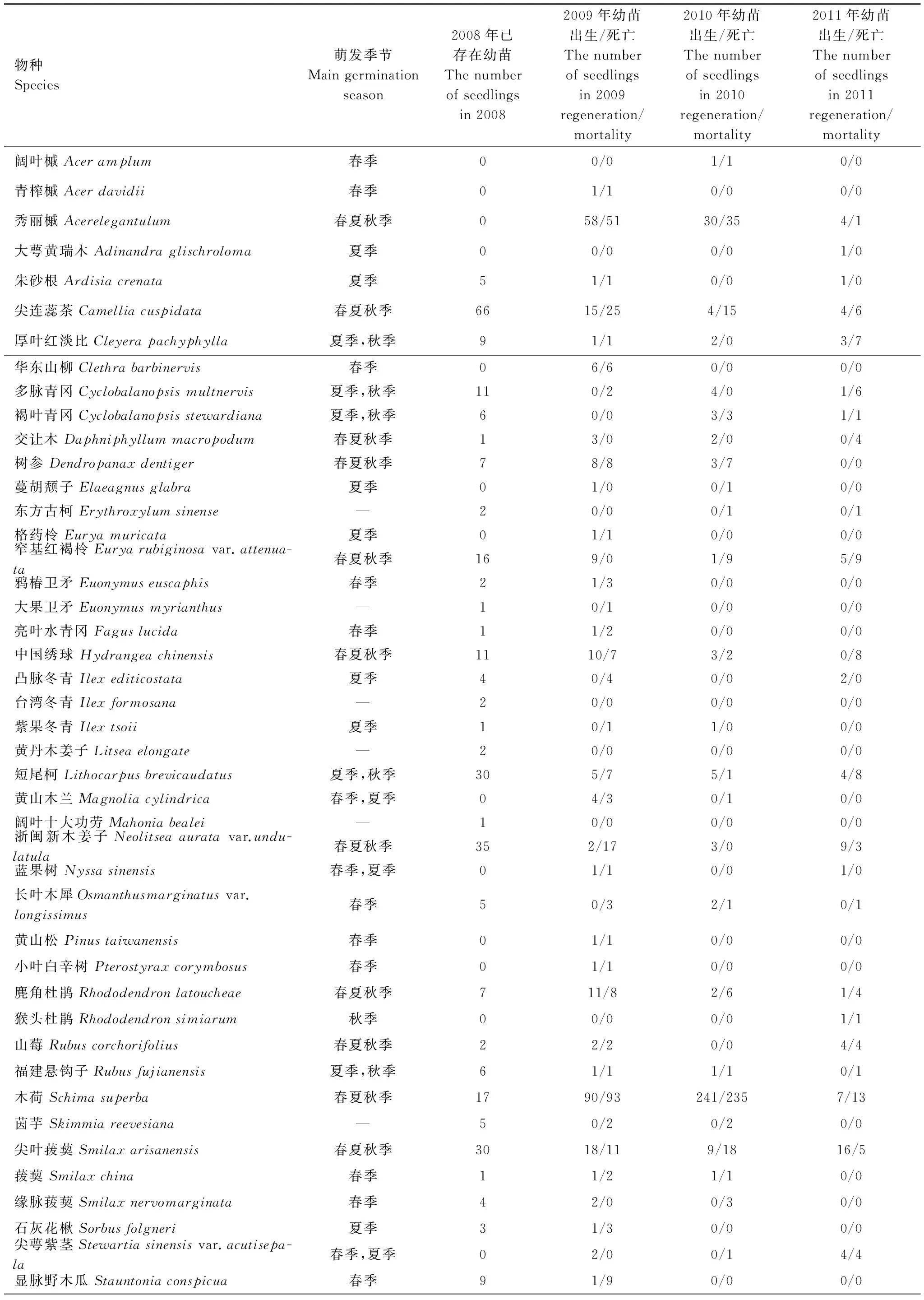
表1 百山祖常绿阔叶林幼苗动态及主要萌发季节
—: 调查期间没有新生幼苗
从图3可以看出,经过3a的动态更新,2008年10月和2011年10月存活的幼苗中,幼苗数量排前5的物种不变,且都占据了总数一半以上,幼苗数量占据前2位的均为光亮山矾和尖连蕊茶。幼苗优势物种除光亮山矾和尖连蕊茶外,还有浙闽新木姜子、尖叶菝葜和短尾柯。
3.2 样地幼苗萌发和死亡的动态
如表2所示,2009年百山祖样地幼苗的3次调查中,幼苗出生比率分别为65.4%、9.9%、3.2%,相应死亡比率为1.9%、7.6%、38.8%。一年周期中,春季幼苗萌发的数量和种类都是最多的,只有少量的幼苗死亡,随着时间的推移,幼苗出生比率不断下降,而死亡比率大幅上升,两者基本抵消,所以对比2008年,2009年的幼苗总数变化不大(每小样方增加不到0.5棵)。由此可得,在百山祖幼苗的存活比率是非常低的,只有极少数的幼苗个体能生存下来。与2009年相比,2010年幼苗各时期的出生比率与之相似,但死亡比率却明显偏大,导致幼苗总数下降。2011年幼苗出生比率和死亡比率都不高,幼苗总数与2010年几乎相同。3a间,幼苗总数下降14.9%,并且维持在360株附近。
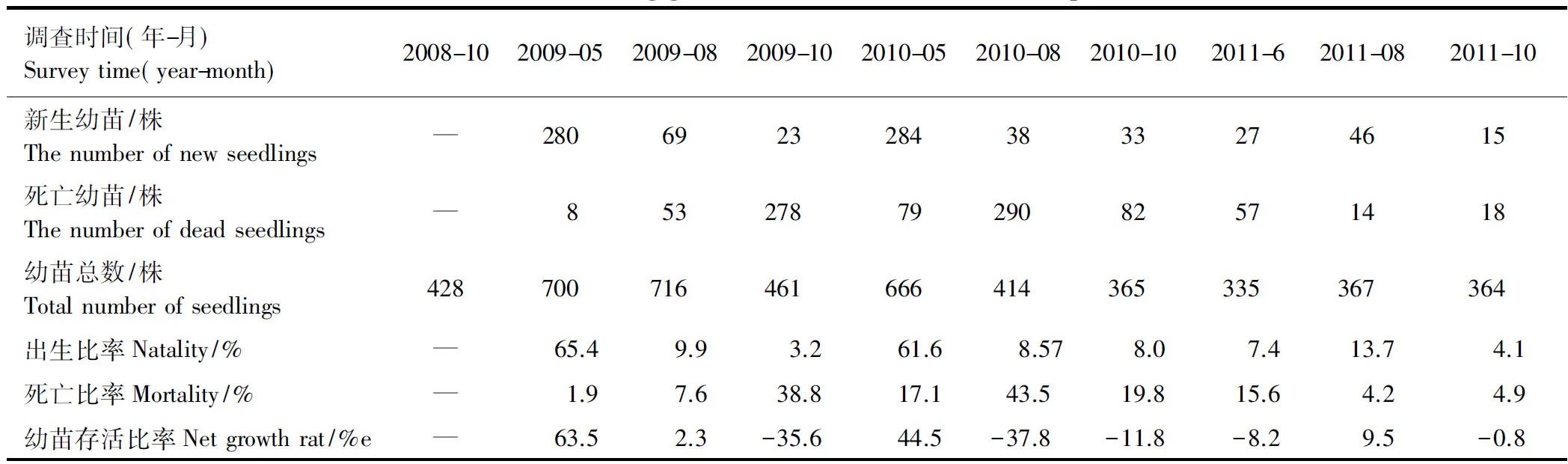
表2 百山祖样地幼苗萌发和死亡情况
—: 2008年第1次调查,尚未能统计该项数据
3.3 小样地幼苗数量动态与生境的相关性
偏相关分析的结果显示幼苗总数与坡位(r=0.329,P=0.000)、土壤水分(r=0.247,P=0.003)存在极显著的正相关性,与落叶层厚薄由薄到厚(r=-0.234,P=0.004)显示极显著的负相关性,而与坡度及土壤质地则无明显相关性。幼苗新生数与5个因素的偏相关性分析结果显示幼苗萌发与土壤水分(r=0.187,P=0.024)存在显著正相关,与其他4个因素无显著相关性。幼苗死亡数与这5个因素都不存在显著的相关性,相对而言,与坡位的相关性较大,沟谷死亡数最高,山脊最低。

表3 偏相关性分析结果
—: 无显著相关性
4 讨论
本研究发现从2008年10月至2011年10月3a间百山祖样地幼苗监测站的幼苗总数减少14.9%,此外幼苗群落无论是科属数还是种数也都呈现减少的趋势。但是3年来,幼苗数量排前五的物种不变,幼苗物种组成并无太大变化,百山祖的幼苗构成表现出明显的优势性和稳定性,各物种幼苗萌发和生存能力之间存在明显差异,因此整个百山祖样地植物群落的物种组成也将基本处于稳定状态。
从幼苗总数在3a间的变化来看,幼苗总数下降主要发生在2010年。百山祖地区当年春季雨季较长,而夏季雨量减少。这种气候异常导致本区域水热条件波动变大,从而引起个体的死亡数增加。然而,由于2009年和2010年大量新生幼苗出现土壤种子库存减少,2011年并没有出现大量幼苗增补。这说明由于某一年气候异常对森林群落造成的影响很难在短时间内恢复。针对百山祖样地的两次群落调查中,相关同事也发现2003年至2008年整个群落中胸径>1 cm的个体也存在数量下降的问题(陈小荣等未发表数据)。根据样地中的枯倒木的年龄推算百山祖样地所在的群落这100多年来鲜受人为的干扰,森林的更新较为缓慢,长久处于一种动态平衡的状态,甚至是近饱和的状态。因此百山祖样地虽然每年都有许多新生幼苗,但存活下来的很少,能长成大树的就更加少。幼苗阶段是森林树种实现更新的最大瓶颈。在四季更替,气候变化的过程中大部分新生幼苗由于不堪忍受苛刻的环境条件而被淘汰。气候和干扰等因素也最易通过影响幼苗更新而影响整个群落的动态。
在幼苗监测站3a共调查到53种植物,但仅有43个物种在调查期间有新的幼苗萌发。幼苗种类与百山祖丰富的树种相比明显偏少,有更新的物种就更少。和其他地区的观测结果类似,本样地的这一调查结果说明了在森林中存在较大的扩散限制[21],离幼苗站较远处的树木并不能将繁殖体扩散至幼苗站。另外,第1次调查时已存在的茵芋、东方古柯、厚皮香等10个物种的幼苗3a未发现新的幼苗。除部分物种依靠根萌更新外,其他物种的更新可能依赖一定的更新生态位和资源的时间波动[22]。
不同物种种子萌发时间的差异也是物种生态位分化的一种表现[23- 24]。通过对观察到的物种种子萌发期的比较,发现43个有新生幼苗的物种的种子萌发时间各不相同。大部分物种种子萌发偏向于春季和夏季。这很大程度上是由于春、夏季水热条件较好,有利于幼苗的存活。同时,很多物种一年中从开始有种子萌发到结束萌发经历的时间较长,大部分物种种子萌发科持续两季以上,有11个树种可在春、夏、秋三季萌发。这说明只要水热条件合适大部分物种可以萌发,种子萌发并无严格的时间限定。密度制约理论认为植物病虫害是限制幼苗更新和植物种群增长最大障碍[25]。本研究结果显示百山祖样地中几个优势种如麂角杜鹃、短尾柯、多脉青冈、木荷、褐叶青冈、厚叶红淡比等都可同时在夏季和秋季萌发。秋季萌发虽然水热条件较差,但是此时病虫害也相应变少。
幼苗更新成功依赖于许多微生境因子,微生境的有利与否,对更新幼苗的发生格局具有决定的作用,成为植物种子能否萌发和生长的决定因素[26]。本研究结果也表明,幼苗更新成功依赖的主要因子是光照和水分。光照是影响幼苗生存的最重要的因素。偏相关分析的结果显示在控制其他变量的前提下,小样方内幼苗总数与土壤水分和坡位成正相关,与落叶层厚度显示负相关,但是由于百山祖的相对湿度比较高,因此坡位最高的裸露的山脊最适合幼苗存活,因为山脊有相对充足的光照,落叶层较薄不易阻挡幼苗根与土壤的接触,同时,由于阳光相对充足,细菌、真菌等微生物不容易滋生。因此,由致病菌引起的幼苗死亡率也相对较低[27]。刘方炎等也指出在森林生态系统中,影响更新幼苗生长的主要因子是温度、光照和水分条件[28]。种子萌发需要较多的水分,环境中具有充足的水分供给干种子吸胀,才能启动种子萌发[11],因此,水分是影响种子萌发的关键因素。本研究结果也显示种子的萌发与水分有显著相关性,而与另外4个因素都无相关性。同时,从时间上看,春季是幼苗出现最多的季节,很大程度上也跟本地区春季多雨有关。
[1] Liu S. Seed Rain and Seedling Regeneration in the Mixed Broad-leaved Korean Pine Forest in Xiaoxing′an Mountains, China [D]. Haerbin: Northeast Forestry University, 2009.
[2] Huang Z Y, Gutterman Y, Hu Z H, Zhang X S. Seed germintion inArtemisiasphaerocephalaII. The influence of environmental factors. Acta Phytoecologica Sinica, 2001, 25(2): 240- 246.
[3] Wang J F, Gao X M, Dong W G. Survival traits of Crofton weed (Eupatoriumadenophorum) seedlings in different habitats. Biodiversity Science, 2008, 16(4): 346- 352.
[4] Clark J S. Why trees migrate so fast: confronting theory with dispersal biology and the paleorecord. The American Naturalist, 1998, 152(2): 204- 224.
[5] Szwagrzy J, Szewczyk J, Bodziarczyk J. Dynamics of seedling banks in beech forest results of a 10 -year study on germination, growth and survival. Forest Ecology and Management, 2001, 141(3): 237- 250.
[6] Wu G L, Du G Z. Relationships between seed size and seedling growth strategy of herbaceous plant: A review. Chinese Journal of Applied Ecology, 2008, 19(1): 191- 197.
[7] Zhang J, Li B H, Bai X J, Yuan Z Q, Wang X G, Ye J, Hao Z Q. Composition and interannual dynamics of tree seedlings in broad-leaved Korean pine (Pinuskoraiensis) mixed forest in Changbai Mountain. Biodiversity Science, 2009, 17(4): 385- 396.
[8] Zhao X Z, Jiang H F, Mao Z J. The studies of the photosynthetic characteristics ofLarixolgensisHenry,LarixkaempferiCarr. AndLarixgmeliniiRupr. seedlings. Bulletin of Botanical Research, 2007, 27(3): 361- 366.
[9] Ding Q, Wang H, Jia G X, Hao Y G. Seed germination and seedling performance ofAmmopiptanthusmongolicus. Journal of Plant Ecology, 2006, 30(4): 633- 639.
[10] Sun Y R, Zhu J J, Yu L Z, Yan Q L, Wang K. Photosynthetic characteristics ofPinuskoraiensisseedlings under different light regimes. Journal of Ecology, 2009, 28(5): 850- 857.
[11] Chen S B, Song A Q, Li Z J. Research advance in response of forest seedling regeneration to light environmental heterogeneity. Chinese Journal of Applied Ecology, 2005, 16(2): 365- 370.
[12] Peng S J, Huang Z L, Peng S L, Ouyang X J, Xu G L. Factors influencing mortality of seed and seedling in plant nature regeneration process. Guihaia, 2004, 24(2): 113- 121.
[13] Ding S Y, Lu X L, Li H M. A comparison of light environmental characteristics for evergreen road-leaved forest communities from different successional stages in Tiantong National Forest Park. Acta Ecologica Sinica, 2005, 25(11): 2862- 2867.
[14] Yang L D, Yang Y, Wang G X, Guo Jian Y. Effects of forest litter on seed germination and seedling growth a review. Chinese Journal of Ecology, 2010, 29(9): 1820- 1826.
[15] Cao F X, Qi C J, Yu X L, Xu Q J, Cao J W, Xu Y F. Conservational significances of evergreen broad-leaved forests in Central China and the strategy of their restoration and rehabilitation. Journal of Central South University of Forestry & Technology, 2010, 30(11): 95- 104.
[16] Zu Y, Zhao G F, Zhang L W, Shen G C, Mi X C, Ren H B, Yu M J, Chen J H, Chen S W, Fang T, Ma K P. Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest. Journal of Plant Ecology, 2008, 32(2): 262- 273.
[17] Xu M, Luo Z R, Yu M J, Ding B Y, Wu Y G. Floristic composition and community structure of mid-montane evergreen broad-leaved forest in North Slope of Baishanzu Mountain. Journal of Zhejiang University Agriculture & Life Sciences, 2007, 33(4): 450- 457.
[18] Wang W, Luo Z R, Zhou R F, Xu D M, Ai J G, Ding B Y. Habitat associations of woody plant species in Baishanzu subtropical broad-leaved evergreen forest. Biodiversity Science, 2011, 19(2): 134- 142.
[19] Jin X F, Ding B Y, Zheng C Z, Ye Z L, Chen X R. The floristic analysis of seed plants in Baishanzu Nature Reserve from Zhejiang Province. Acta Botanica Yunnanica, 2004, 26(6): 605- 618.
[20] Yu J H, Yao F P, Chen X R, Zhou R F, Cheng Q B, Ding B Y. An introduction to main vegetation types in the Baishanzu national nature reserve. Journal of Tropical and Subtropical Botany, 2003, 11(2): 93- 98.
[21] Nathan R, Muller-Landau H C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends in Ecology & Evolution, 2000, 15(7): 278- 285.
[22] Shen G C, Yu M J, Hu X S, Mi X C, Ren H B, Sun I F, Ma K P. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity. Ecology, 2009, 90(11): 3033- 3041.
[23] Chesson P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 2000, 31(1): 343- 366.
[24] Wright J. Plant diversity in tropical forests: a review of mechanisms of species coexistence. Oecologia, 2002, 130(1): 1- 14.
[25] Janzen D H. Herbivores and the number of tree species in tropical forests. The American Naturalist, 1970, 104(940): 501- 528.
[26] Chen D M. Analysis on Microsite ofPiceaSchrenkianaNatural Rregeneration and Seedling Spatial Pattern and Dynamic [D]. Wulumuqi: Xinjiang Agricultural University, 2006.
[27] Augspurger C K. Seedling survival of tropical tree species: interactions of dispersal distance, light-gaps and pathogens. Ecology, 1984, 65(6): 1705- 1712.
[28] Liu F Y, Zhang Z X, Wang X Q, Li K, Sun Y Y, Zhang C H. Effects of habitat heterogeneity on early growth ofQuercusfranchetiinatural regeneration seedlings in the Jinsha River dry-hot valley. Chinese Journal of Applied & Environmental Biology, 2011, 17(3): 338- 344.
参考文献:
[1] 刘双. 小兴安岭阔叶红松林种子雨及幼苗更新 [D]. 哈尔滨: 东北林业大学硕士学位论文, 2009.
[2] 黄振英, 胡正海, 张新时. 白沙蒿种子萌发特性研究. Ⅱ 环境因素的影响. 植物生态学报, 2001, 25(2): 240- 246.
[3] 王瑾芳, 高贤明, 党伟光. 不同生境条件下紫茎泽兰实生幼苗的生存特性比较. 生物多样性, 2008, 16(4): 346- 352.
[6] 武高林, 杜国祯. 植物种子大小与幼苗生长策略研究进展. 应用生态学报, 2008, 19(1): 191- 197.
[7] 张健, 李步杭, 白雪娇, 原作强, 王绪高, 叶吉, 郝占庆. 长白山阔叶红松林乔木树种幼苗组成及其年际动态. 生物多样性, 2009, 17(4): 385- 396.
[8] 赵溪竹, 姜海凤, 毛子军. 长白落叶松、日本落叶松和兴安落叶松幼苗光合作用特性比较研究. 植物研究, 2007, 27(3): 361- 366.
[9] 丁琼, 王华, 贾桂霞, 郝玉光. 沙冬青种子萌发及幼苗生长特性. 植物生态学报, 2006, 30(4): 633- 639.
[10] 孙一荣, 朱教君, 于立忠, 闫巧玲, 王凯. 不同光环境对红松幼苗光合生理特征的影响. 生态学杂志, 2009, 28(5): 850- 857.
[11] 陈圣宾, 宋爱琴, 李振基. 森林幼苗更新对光环境异质性的响应研究进展. 应用生态学报, 2005, 16(2): 365- 370.
[12] 彭闪江, 黄忠良, 彭少麟, 欧阳学军, 徐国良. 植物天然更新过程中种子和幼苗死亡的影响因素. 广西植物, 2004, 24(2): 113- 121.
[13] 丁圣彦, 卢训令, 李昊民. 天童国家森林公园常绿阔叶林不同演替阶段群落光环境特征比较. 生态学报, 2005, 25(11): 2862- 2867.
[14] 羊留冬, 杨燕, 王根绪, 郭剑英. 森林凋落物对种子萌发与幼苗生长的影响. 生态学杂志, 2010, 29(9): 1820- 1826.
[15] 曹福祥, 祈承经, 喻勋林, 徐庆军, 曹基武, 徐永福. 中国中部亚热带常绿阔叶林保育的重要性及其恢复重建策略. 中南林业科技大学学报, 2010, 30(11): 95- 104.
[16] 祝燕, 赵谷风, 张俪文, 沈国春, 米湘成, 任海保, 于明坚, 陈建华, 陈声文, 方腾, 马克平. 古田山中亚热带常绿阔叶林动态监测样地-群落组成与结构. 植物生态学报, 2008, 32(2): 262- 273.
[17] 徐敏, 骆争荣, 于明坚, 丁炳扬, 吴友贵. 百山祖北坡中山常绿阔叶林的物种组成和群落结构. 浙江大学学报: 农业与生命科学版, 2007, 33(4): 450- 457.
[18] 王伟, 骆争荣, 周荣飞, 许大明, 哀建国, 丁炳扬. 百山祖常绿阔叶林木本植物的生境相关性分析. 生物多样性, 2011, 19(2): 134- 142.
[19] 金孝峰, 丁炳扬, 郑朝宗, 叶珍林, 陈小荣. 浙江百山祖自然保护区种子植物区系分析. 云南植物研究, 2004, 26(6): 605- 618.
[20] 余久华, 姚丰平, 陈小荣, 周荣飞, 程秋波, 丁炳扬. 百山祖自然保护区主要植被类型概述. 热带亚热带植物学报, 2003, 11(2): 93- 98.
[26] 陈迪马. 天山云杉天然更新微生境及其幼苗格局与动态分析 [D]. 乌鲁木齐: 新疆农业大学, 2006.
[28] 刘方炎, 张志祥, 王小庆, 李昆, 孙永玉, 张春华. 生境异质性对金沙江干热河谷锥连栎天然更新幼苗早期生长的影响. 应用与环境生物学报, 2011, 17(3): 338- 344.
Woody seedling dynamics and the correlation between habitat and regeneration / mortality in a subtropical evergreen broad-leaved forest in China
SHI Lulu1, LUO Zhengrong2, XIA Jiatian3, ZHAO Wenjuan1, WU Yougui3, DING Bingyang1,*
1CollegeofLifeandEnvironmentalScience,WenzhouUniversity,Wenzhou325035,China2CollegeofEcology,LishuiUniversity,Lishui323000,China3ManagementofBaishanzu,Fengyangshan-BaishanzuNationalNatureReserve,Qingyuan323800,China
Seedling recruitment is important for forest regeneration. During the seedling phase, individual plants are sensitive to a wide variety of habitat conditions. Seedling establishment depends not only on the specific species involved, but also on the site specific conditions where the plants grow. However, few studies have examined the ways in which habitat influences the survival and growth of seedlings. In 2003, a single 5 hm2(250 m × 200 m) permanent plot was established in a subtropical evergreen broad-leaved forest in the Baishanzu mountainous region, Zhejiang Province, China (119°3′53″E, 27°40′54″N) to monitor the dynamics of the forest. To explore the dynamics of seedling survival as well as the correlation between the seedling mortality/germination and the habitats of various seedlings in the plot, 50 seed-seedling monitoring stations were established along the inner path of the plot in October 2008. Each seed-seedling monitor station consists of a seed collector (0.75 m × 0.75 m) and 3 seedling quadrats (1 m ×1 m), which were placed 2 m away from the seed collector. The quadrats were surveyed three times each year to tag, identify and quantify the emergence of new germinating seedlings and to track seedling mortality in these quadrats during May, August and October, from 2008 to 2011. During data analysis, we first calculated the germination rate, death rate and survival rate (the difference between the germination and death rates) of all seedlings present during the period. Then, the numbers of seedlings, newly germinated seedlings, and dead seedlings were classified based on slope position, slope degree, soil texture, soil moisture, and depth of leaf litter in the seedling stations by partial correlation. The results show that: 1) the seedlings that geminated between October, 2008 and October, 2011 in the Baishanzu plot belonged to 53 species, 40 genera, 26 families; 2) different species germinated at different times and the duration of the germination period during a year varied among these species with 11 species germinating during spring to autumn, five species germinating in spring and summer, five species germinating in summer and autumn, 13 species germinating only in spring, eight species germinating only in summer, one species germinating only in autumn, while the remaining ten species did not germinate during the study period; 3) the total number of seedlings declined from 428 in October 2008 to 364 in October 2011; also, the number of species present in the quadrats declined from 53 to 45 during this study; 4) the seedling survival rates were 7.7%, -20.8%, and -0.3% in 2009, 2010 and 2011, respectively; 5) dominant species in living seedlings present in the plot wereSymplocoslucida,Camelliacuspidata,Neolitseaauratavar.undulatula,SmilaxarisanensisandLithocarpusbrevicaudatus; these five species included > 50% of the total number of plants present in the quadrats; 6) the number of germinating seedlings in a particular seedling quadrat was correlated with the habitat type, which was itself determined by water availability at the site; 7) the number of surviving seedlings in a quadrat was significantly correlated to slope position, moisture content, and depth of leaf litter of the seedling quadrat. These results suggest that species identity, water availability and light were the major factors affecting seedling germination and survival. Species exhibit differences in regeneration niche during the seedling stage of growth.
Baishanzu; seedling; species composition; quantity dynamic; habitats
中国科学院知识创新工程重要方向项目(KZCX2-YW- 430); 浙江省大学生科技创新项目(2009R424027)
2013- 02- 15; 网络出版日期:2014- 03- 17
10.5846/stxb201302150268
*通讯作者Corresponding author.E-mail: dby@wzu.edu.cn
施璐璐, 骆争荣,夏家天,赵文娟, 吴友贵, 丁炳扬.亚热带中山常绿阔叶林木本植物幼苗数量动态及其与生境的相关性.生态学报,2014,34(22):6510- 6518.
Shi L L, Luo Z R, Xia J T, Zhao W J, Wu Y G, Ding B Y.Woody seedling dynamics and the correlation between habitat and regeneration / mortality in a subtropical evergreen broad-leaved forest in China.Acta Ecologica Sinica,2014,34(22):6510- 6518.
