盐分和温度对万寿菊种子发芽的影响
2014-08-10毛子军
刘 敏,厉 悦,毛子军,刘 龙
(1.东北林业大学森林植物生态学教育部重点实验室,黑龙江 哈尔滨 150040;2.齐齐哈尔大学生命科学与农林学院,黑龙江 齐齐哈尔 161006)
盐分和温度对万寿菊种子发芽的影响
刘 敏1,2,厉 悦2,毛子军1,刘 龙2
(1.东北林业大学森林植物生态学教育部重点实验室,黑龙江 哈尔滨 150040;2.齐齐哈尔大学生命科学与农林学院,黑龙江 齐齐哈尔 161006)
为明确万寿菊(Tageteserecta)在种子发芽阶段对盐分、温度及其交互作用的响应,采用5个盐分浓度和3个温度处理分别模拟土壤盐分胁迫条件和气候变暖状况进行研究。结果表明,万寿菊种子能适应低浓度(2 g·L-1)的盐分环境,但中高浓度盐胁迫对种子发芽产生明显的抑制作用。在无盐及低盐(2 g·L-1)环境下,万寿菊种子能适应较宽的温度范围。盐胁迫与温度交互作用显著(P<0.05)。一方面,盐分浓度会影响温度对万寿菊种子发芽和幼苗生长的作用。在无盐及低盐分浓度下,3个温度组的发芽率无显著差异(P>0.05),但在4~6 g·L-1盐分浓度下,高温组万寿菊种子的发芽率和幼苗苗高和根长显著低于另外两个温度组(P<0.05)。另一方面,温度也会影响盐分浓度对万寿菊种子发芽和幼苗生长的作用。在15/25 ℃与20/30 ℃变温环境下,只有较高盐分浓度(≥6 g·L-1)时种子发芽率才显著降低(P<0.05);而在25/35℃的高温变温环境中,盐分浓度≥4 g·L-1时种子发芽率、幼苗苗高和根长就显著下降(P<0.05)。万寿菊种子的发芽指数比发芽率对盐分胁迫的反应更加敏感。
万寿菊;种子发芽;幼苗生长;交互作用;盐害指数
万寿菊(Tageteserecta)为菊科(Asteraceae)植物,一、二年生草本花卉。万寿菊花大、色艳,花期长,观赏价值高,是常用的园林绿化植物;其因具有清热解毒、化痰止咳、补血通经及消肿功效,是我国传统的中药材;此外,万寿菊中的叶黄素,广泛应用于食品着色剂、饲料添加剂领域和功能性保健食品的研发和生产方面。由于其具有很高的经济、药用和观赏价值,被全国各地广泛栽培。
当前,土壤盐渍化是一个世界性的资源问题和生态问题。全球盐碱地面积已达 9.5×108hm2,中国盐渍土总面积约 1×108hm2,其中现代盐渍化土壤约3.7×107hm2,残余盐渍化土壤约4.5×107hm2,潜在盐渍化土壤约1.7×107hm2[1]。盐碱地由于土壤中含盐量高,很多草本花卉在这些地区栽培都会受到不同程度的盐胁迫,甚至无法存活。种子发芽是植物生长最敏感的阶段之一[2],种子在盐胁迫下萌发成苗,是植物在盐碱环境下生存的前提。因此,有必要研究种子在盐胁迫下的萌发生理[3]。
目前,有部分学者对万寿菊种子发芽进行了研究,如模拟酸雨[4]、融雪剂[5]、沼液浸种[6]对万寿菊种子萌发、幼苗生长的影响等;吴志刚等[7]认为,用标准法催芽时,万寿菊种子发芽最适温度是25 ℃,但该试验只考虑了恒温条件下万寿菊种子的发芽情况,没有考虑室外实际的日夜变温情况;另外,盐胁迫试验表明,万寿菊种子发芽期耐盐能力较强[8-9]。有研究认为,盐胁迫下的发芽率与温度高低会有一定的关系[3,10],那么在全球气候变暖的今天,温度的升高对盐碱地区万寿菊种子的发芽是不是有影响呢?本研究根据室外日夜变温情况,利用人工光照培养箱模拟气候变暖和日夜变温,设置3个温度梯度和5个盐分(NaCl)浓度,探讨气温升高和盐分胁迫下万寿菊种子的发芽和幼苗生长情况,以期为气候变暖情况下盐碱地区万寿菊的栽培管理提供理论依据,同时也为我国盐渍化地区的土壤改良和城市绿化及生态重建等提供理论指导。
1 材料与方法
1.1 材料
以万寿菊的迅雷品种为研究对象,种子来源于内蒙古赤峰春禾园艺科研开发有限责任公司。处理前挑选大小一致且颗粒饱满的种子,千粒重为(0.363±0.004)g。用于盐胁迫的盐为化学分析纯NaCl。
1.2 方法
本研究采用双因素(盐分浓度和温度)交叉设计方法。盐分设5个梯度,用蒸馏水分别配制0(CK,蒸馏水)、2、4、6、8 g·L-15个不同NaCl浓度的溶液;温度设置3个梯度,分别为15/25 ℃、20/30 ℃、25/35 ℃。共15个处理,每个处理3次重复。在3个SPX-300-PG人工光照培养箱中进行试验,每个培养箱进行一个温度处理。人工光照培养箱设置如下:将一天分为4段,第1阶段为09:00-15:00,光照强度为5 000 lx,3个培养箱温度分别设置为(25±1) ℃、(30±1) ℃和(35±1) ℃。第2阶段为15:00-19:00,光照强度为2 000 lx,温度设置与第1阶段相同;第3阶段为19:00-04:00,光照强度为0,温度分别设置为(15±1) ℃、(20±1) ℃和(25±1) ℃;第4阶段为04:00-09:00,光照强度为2 000 lx,温度与第3阶段设置相同。
试验前先将万寿菊种子用1 g·L-1的KMnO4溶液浸泡消毒30 min,蒸馏水冲洗3遍,放入大烧杯中置于水温恒定为40 ℃的水浴箱中,温水浸种处理2 h。将培养皿(直径9 cm)洗净、烘干,内置两层滤纸(直径9 cm),每个培养皿加各处理溶液10 mL,然后每皿均匀摆放100粒消毒和温汤浸种处理后的万寿菊种子。每粒种子之间留有一定的间隙,便于发芽和观察计数。盖好盖子,贴好标签,分别放置于设置好的光照培养箱中,进行发芽试验。每天上午定时观察种子发芽情况并记录。发芽标准以胚根突破种皮且下胚轴长度不小于种子自身长度的1/2为基准,直至连续3 d发芽数不再增加时终止发芽试验。试验期间利用倾斜45°培养皿无明显溶液方法补充蒸馏水以保持盐分浓度[3]。试验中霉烂的种子用升汞溶液消毒后放回原处继续观察,严重霉烂的种子将其挑出,避免感染其他种子,并将其记录为未发芽种子。终止发芽试验时,每处理选长势一致的幼苗 10株,测量根长和苗高。
1.3 数据处理
在试验结束后计算种子发芽率、发芽指数和盐害指数。发芽率(GP)=种子最终发芽数/供试种子总数×100%。发芽指数(GI)=∑(Gt/Dt)。式中,Gt为t天的发芽数,Dt为相应的发芽天数。盐害指数=(对照发芽率-处理发芽率)/对照发芽率×100%[11]。使用SPSS 19.0统计软件对所测数据进行统计分析,测定结果用平均值±标准差表示,分别对同一盐分浓度不同温度处理、同一温度不同盐分浓度处理进行单因素方差分析,并对温度和盐分浓度进行双因素分析,用Duncan法和LSD法对各测定数据进行多重比较;采用Excel 2007进行制图。
2 结果与分析
2.1 盐分和温度对万寿菊种子发芽率和盐害指数的影响
由双因素方差分析得知,盐分浓度、温度以及盐分浓度与温度的交互作用都对万寿菊种子的发芽率有极显著影响(P<0.01)(表1)。
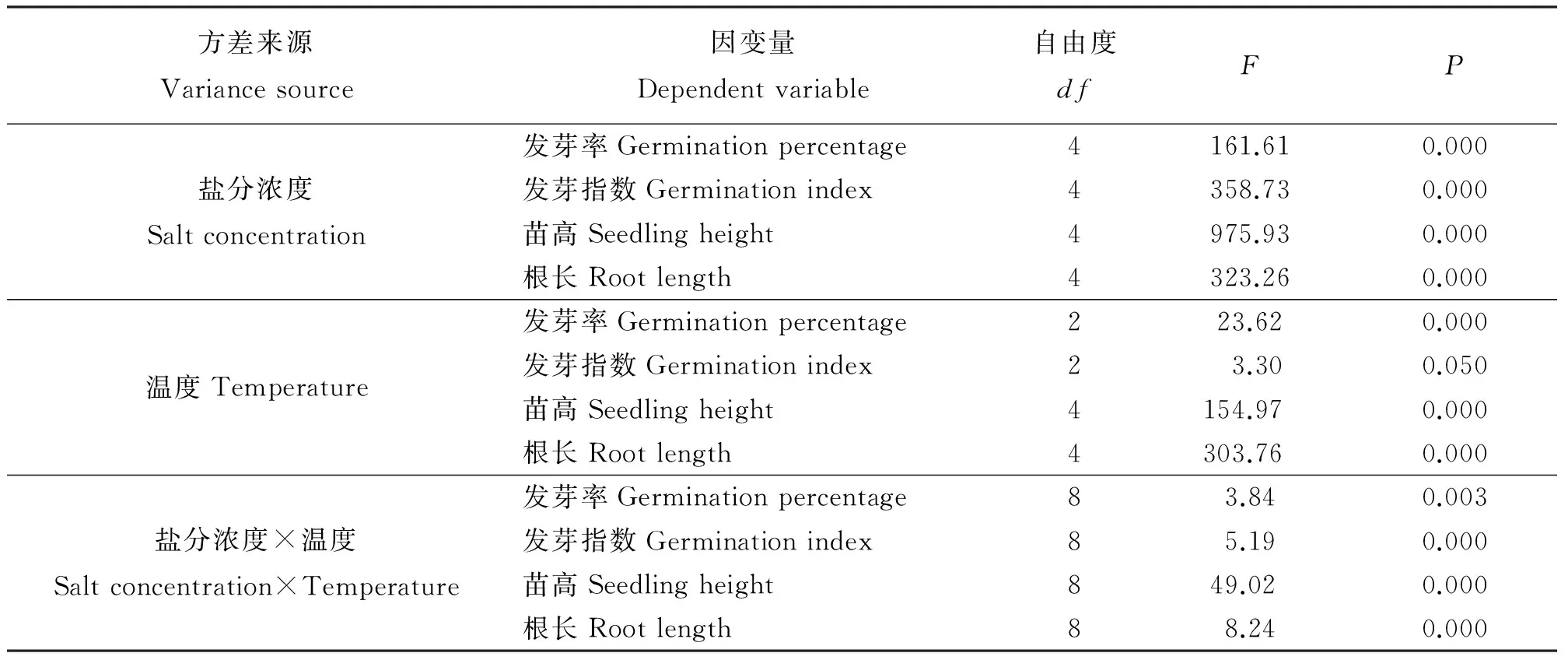
表1 不同因素影响下万寿菊种子发芽及幼苗生长的组间效应分析
随着盐分浓度的增高,在各温度处理下万寿菊种子的发芽率总体呈下降趋势(图1),而其盐害指数呈上升趋势(图2),特别是在高盐分浓度环境中变化非常显著(P<0.05),反映出高浓度盐分严重抑制了万寿菊种子的发芽。
盐分浓度与温度对种子发芽的交互作用比较明显,表现在以下两个方面。第一,温度对种子发芽率和盐害指数的影响程度受盐分浓度的影响比较明显。在无盐及2 g·L-1低盐分浓度下,3个温度组的发芽率未表现出显著差异(P>0.05)。随着盐分浓度的增大,25/35 ℃的高温组万寿菊种子的发芽率显著低于另外两个温度组(P<0.05),表明高温严重抑制了中等盐分环境下万寿菊种子的发芽率。然而,在8 g·L-1高盐分浓度下,虽然高温组的发芽率低于其他两个温度组,但3个温度组的发芽率差异不显著(图1)。说明在高盐分环境下,盐分对万寿菊种子发芽的抑制作用比高温的抑制作用更明显。第二,盐分浓度对种子发芽率的影响程度受温度的影响也比较明显。在15/25 ℃与20/30 ℃的温度条件下,≤4 g·L-1的低盐分浓度对种子发芽的抑制作用不显著,盐害指数低,甚至在2 g·L-1时的发芽率是最高,盐害指数为负数,但与对照差异不显著,只有盐分浓度大于6 g·L-1时种子发芽率才极显著降低,盐害指数急剧增高。而在25/35 ℃的高温环境中,盐分浓度≥4 g·L-1时种子发芽率就极显著下降,盐害指数明显升高(图2)。
2.2 盐分和温度对万寿菊种子发芽指数的影响
发芽指数是衡量种子活力的重要指标,反映了种子发芽速度和发芽整齐度。发芽指数高,种子活力强,在自然条件下能快速出苗[12]。盐分浓度、温度以及盐分浓度与温度的交互作用都显著影响(P≤0.05)万寿菊种子的发芽指数(表1)。3个温度处理下万寿菊种子的发芽指数都随着盐分浓度的增加而迅速降低(图3)。在最高盐分浓度下,3个温度处理的发芽指数分别为对照组的17.64%、17.77%和10.41%,说明盐胁迫对万寿菊种子活力的抑制作用很明显。与发芽率不同的是,在15/25 ℃温度条件下,盐分浓度≥4 g·L-1时种子发芽指数极显著下降(P<0.01),在20/30 ℃和25/35 ℃温度条件下,盐分浓度≥2 g·L-1时种子发芽指数显著下降。 无盐胁迫时,25/35 ℃高温组的发芽指数最高,20/30 ℃组次之,15/25 ℃组最低,主要是由于高温环境中的前2 d万寿菊种子发芽速度率明显高于其他两组,后期趋于平缓,说明短时间的高温对种子发芽起促进作用。而在6 g·L-1盐分浓度时,高温组的发芽指数显著低于其他两个温度组(P<0.05),说明较高盐分浓度下高温反而抑制种子的发芽活力。
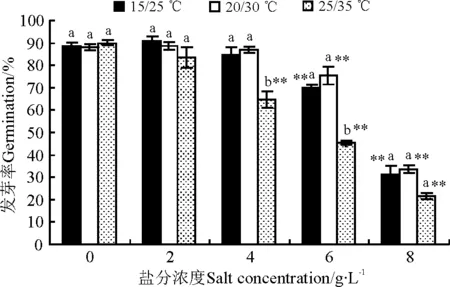
图1 盐分和温度对万寿菊种子发芽率的影响
注:不同小写字母表示同一盐分浓度不同温度处理间差异显著(P<0.05);*和**分别表示同一温度下不同盐分浓度处理组与无盐处理组间的差异显著(P<0.05)和极显著(P<0.01)。下同。
Note: Different lower case letters for the same concentration indicate significant difference between different temperature treatments under the same salt concentration at 0.05 level; * and ** indicate significant difference between different salt treatments under the same temperature at 0.05 and 0.01 level, respectively.The same blow.
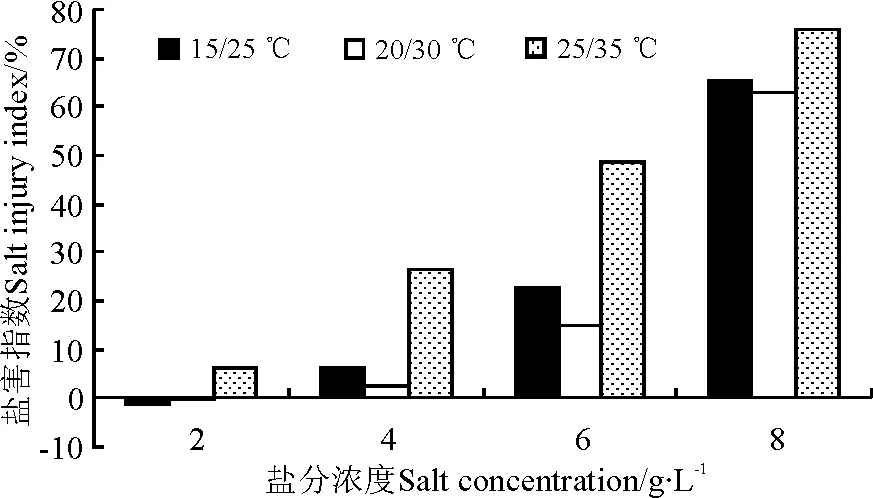
图2 盐分和温度对万寿菊种子发芽的盐害指数
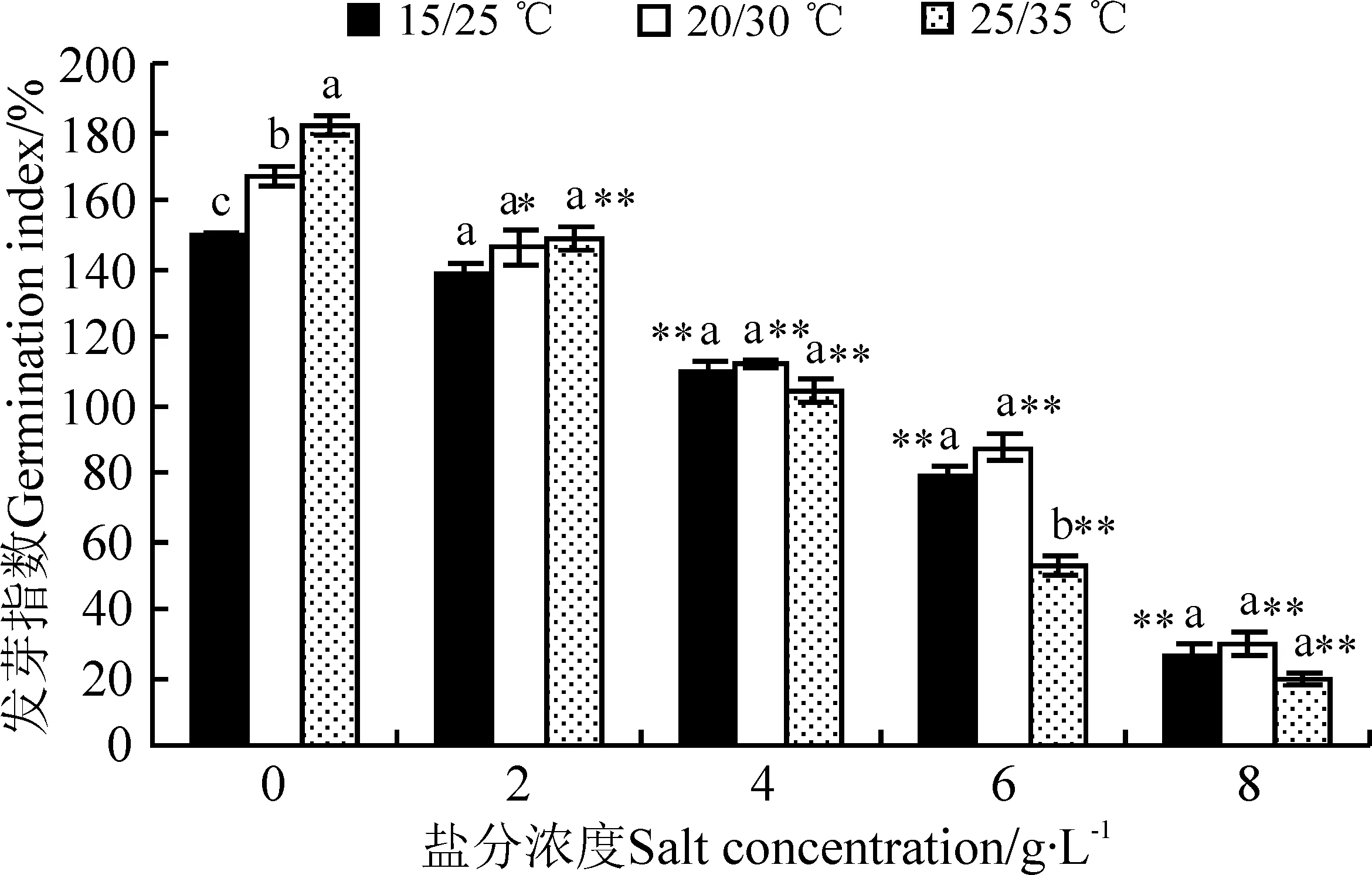
图3 盐分和温度对万寿菊种子发芽指数的影响
2.3 盐分和温度对万寿菊幼苗生长的影响
方差分析(表1)显示,盐分浓度、温度以及盐分浓度与温度的交互作用都对万寿菊幼苗的苗高和根长有极显著影响(P<0.01)。万寿菊幼苗苗高和根长受盐分的影响很相似,在同一温度下,苗高和根长都是随着盐分浓度的增加而呈下降趋势,除2 g·L-1盐分浓度组与无盐组差异不显著(P>0.05)外,其余各盐分组都与对照组差异显著(P<0.05)(图4、5)。万寿菊幼苗苗高和根长受温度的影响略有差异,但温度与盐分之间的交互作用对它们的影响都很明显。无盐和2g·L-1低盐环境中,25/35 ℃高温组的苗高显著高于其他两个温度组;而其他盐分中25/35 ℃高温组的苗高显著低于其他两个温度组。根长受温度的影响与苗高略有不同,所有盐分浓度下25/35 ℃高温组的根长都显著低于其他两个温度组。
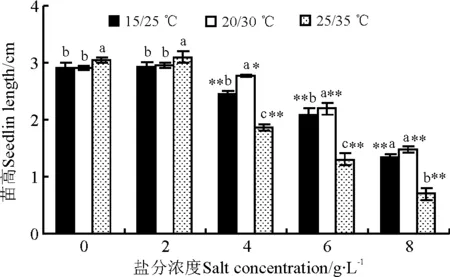
图4 盐分和温度对万寿菊幼苗苗高的影响

图5 盐分和温度对万寿菊幼苗根长的影响
3 讨论与结论
本研究表明,万寿菊种子能适应低浓度的盐胁迫,3个温度水平都表现为2 g·L-1盐分浓度组与对照组的发芽率和幼苗生长无明显差异,但中、高浓度盐胁迫对种子发芽和幼苗的生长产生明显的抑制作用,高浓度盐分抑制作用最显著(P<0.05),种子发芽情况与张桂玲[8]和刘淑贤[9]对万寿菊种子的研究结果相似,也与张立磊和王少平[13]对同属的孔雀草(Tagetespatula)种子发芽的研究相似;幼苗生长情况与刘敏等[14]对万寿菊幼苗的研究结果和林伟等[11]对孔雀草的研究结果一致。高浓度盐分抑制万寿菊种子发芽和幼苗生长的作用机制,可能是NaCl毒害的结果[15]、离子渗透效应[16],或者是NaCl毒害和离子渗透综合作用的结果[17-18]。
在无盐及低盐(2 g·L-1)胁迫下,3个温度处理之间的种子发芽率差异不显著(P>0.05),说明万寿菊的种子发芽在无盐和低浓度盐胁迫下能够适应较宽的温度范围。这与蔺吉祥等[19]对小麦(Triticumaestivum)种子发芽的研究结果一致,但吴志刚等[7]认为,万寿菊种子在25 ℃下发芽率最高,其次为20、30 ℃,35 ℃时最低,高温会抑制万寿菊种子的发芽。可能是由于本研究采用的是有10 ℃日夜温差的变温条件,虽然20/30 ℃组和25/35 ℃组白天高温时(09:00-19:00,10 h)温度分别为30和35 ℃的高温,但傍晚至凌晨(19:00-09:00,14 h)相对凉爽,只有20和25 ℃,不至于使种子一直遭受高温胁迫。15/25 ℃组虽然白天高温时(09:00-19:00,10 h)温度为25 ℃,是吴志刚等[7]研究的万寿菊种子最适发芽温度,但傍晚至凌晨(19:00-09:00,14 h)温度只有15 ℃,温度较低,会影响种子发芽。高温组夜间的低温缓解了白天高温对种子发芽的抑制作用,常温组夜间的低温抑制了种子发芽,因此3个温度组的发芽率差异不显著。应该说变温情况更接近室外自然环境,本研究对室外植物管理来说更具有应用价值。
在无盐及低盐(2 g·L-1)环境中,25/35 ℃高温组的万寿菊幼苗有徒长的现象,虽然其苗高比其他两个温度组高,但其径较细,植株比较弱。随着盐分浓度的增大,高温环境中幼苗高度和根长都急剧下降,显著低于其他两个温度组。
综合种子发芽率、盐害指数、发芽指数和幼苗生长情况可知,盐胁迫与温度对万寿菊种子发芽的交互作用明显,这与Al-Helal等[10]、刘寅哲和张义[3]、高瑞如[20]、蔺吉祥等[21]的结果一致,体现出盐胁迫下的发芽率与温度高低也有一定的关系,但不同物种的具体表现有所差别。本研究显示,盐分浓度会影响温度对万寿菊种子发芽和幼苗生长的作用。在无盐及低盐分浓度(2 g·L-1)下,3个温度组的发芽率未表现出显著差异(P>0.05),但在4~6 g·L-1盐分浓度下,25/35 ℃的高温组万寿菊种子的发芽率显著低于另外两个温度组(P<0.05)。说明在中高盐分浓度时,万寿菊种子对温度的适应范围变窄,不能忍受高温环境。幼苗生长的数据同样显示,在高盐环境中高温严重抑制了苗高和根长的生长,植株虽然能发芽,但幼苗弱小,抗性弱,观赏价值低。另外,温度也会影响盐分浓度对万寿菊种子发芽的作用。在中低温度(15/25 ℃与20/30 ℃)环境下,只有较高盐分浓度(≥6 g·L-1)时种子发芽率才显著降低,说明中低温度环境下其耐盐范围较宽;而在25/35 ℃的高温环境中,盐分浓度≥4 g·L-1时,种子发芽率、苗高和根长就下降显著,高温组下降的速率明显高于另外两个温度组,说明高温下种子的耐盐范围下降明显。高温环境中盐分胁迫会使种子酶活性紊乱,影响正常的生理代谢活动,高温还会加速水分蒸发,进而增加盐分浓度,此环境下造成的伤害往往是不可逆的[21-22]。
通过盐分与温度的互作效应可预测,在无盐或低盐(2 g·L-1)的环境中,气候变暖对万寿菊种子的发芽和幼苗生长影响不大;但在中等及较高盐分浓度环境中,气候变暖温度达到高温(25/35 ℃)时盐和高温共同作用将会严重抑制万寿菊种子的发芽和幼苗生长,影响万寿菊在盐渍地区的种植。在高盐分浓度(≥8 g·L-1)地区,无论什么温度环境下,万寿菊种子的发芽率都很低,幼苗生长状况也很差,不适宜万寿菊的种植。
万寿菊种子的发芽指数从低盐分浓度开始就明显低于对照组,相比于种子发芽率,种子发芽指数较发芽率对NaCl胁迫的反应更加敏感,与宋彬等[23]的研究结果一致,说明盐胁迫下万寿菊种子发芽的整齐程度首先受到影响,在引种栽培中将会影响到出苗品质。
[1] 伍林涛,杜才富,邵明波.植物盐胁迫耐受性研究进展[J].吉林农业,2010(9):51-52.
[2] Grime J P,Campbell B D.Growth rate,habitat productivity,and plant strategy as predictors of stress response[A].Winner W E,Pell E J,Roy J.Response of Plants to Multiple Stresses[M].Burlington:Academic Press Incorporated,1991:143-159.
[3] 刘寅哲,张义.不同温度下复合盐胁迫及恢复对紫花苜蓿种子发芽特性的影响[J].种子,2009,28(11):22-25.
[4] 范小美.模拟酸雨对一串红、万寿菊种子萌芽的影响[J].安徽农业科学,2007,35(6):1595-1596.
[5] 李芳,张俊民.融雪剂对园林植物种子萌发的影响[J].环境科技,2009,22(5):22-24.
[6] 袁大刚,刘成,蒲光兰,吴德勇.沼液浸种对万寿菊种子发芽及幼苗生长的影响[J].中国中药杂志,2011,36(7):817-822.
[7] 吴志刚,王平,吕双双,赵景云,李娜,张玉静,崔玥晗.万寿菊种子发芽实验[J].中国种业,2007(11):54-55.
[8] 张桂玲.盐胁迫下8种草花种子萌发特性的比较[J].作物杂志,2011(6):110-112.
[9] 刘淑贤.几种露地花卉种子萌发期耐盐性研究[J].内蒙古民族大学学报(自然科学版),2002,17(3):228-230.
[10] Al-Helal A A,Al-Farraj M M,El-Desoki R A.Germination response ofCassiasennaL.seeds to sodium salts and temperature[J].Journal of the University of Kuwait(Science),1989,16(2):281-288.
[11] 林伟,李璟,蔡仕珍.几种钠盐对孔雀草萌发期胁迫效应的比较[J].种子,2013,32(5):96-99.
[12] 颜宏,矫爽,赵伟,闫修民,周道玮,姜百川.不同大小碱地肤种子的萌发耐盐性比较[J].草业学报,2008,17(2):26-32.
[13] 张立磊,王少平.盐胁迫对万寿菊种子发芽的影响[J].北方园艺,2012(23):86-83.
[14] 刘敏,厉悦,梁艳,田立广.不同浓度氯化钠胁迫对万寿菊幼苗生长及生理特性的影响[J].北方园艺,2013(24):63-66.
[15] Sosa L,Llanes A,Reinoso H,Reginato M,Reginato M,Luna V.Osmotic and specific ion effects on the germination of Prosopis strombulifera[J].Annals of Botany,2005,96:261-267.
[16] Pujol J A,Calvo J F, Ramírez-díaz L.Recovery of germination from different osmotic conditions by four halophytes from southeastern Spain[J].Annals of Botany,2000,85:279-286.
[17] Song J,Feng G,Tian C,Zhang F S.Strategies for adaptation ofSuaedahysophora,HaloxylonammodendronandHaloxylonpersicumto a saline environment during seed-germination stage[J].Annals of Botany,2005,96:399-405.
[18] Williamj K,Irwina U,Johnp M.Effect of salinity on germination and seedling growth of twoAtriplexsepecies(Chenopodiaceae)[J].Annals of Botany,1998,82:167-175.
[19] 蔺吉祥,李晓宇,唐佳红,王俊锋,李卓琳,张兆军,穆春生.温度与盐、碱胁迫交互作用对小麦种子萌发的影响[J].作物杂志,2011(6):113-116.
[20] 高瑞如,赵瑞华,张双风,邸丽珍,黄培祐.盐分和温度对盐节木种子萌发的影响[J].西北植物学报,2007,27(11):2281-2285.
[21] 蔺吉祥,李晓宇,张兆军,王俊锋,李卓琳,穆春生.温度与盐碱胁迫交互作用对羊草种子萌发与幼苗生长的影响[J].草地学报,2011,19(6):1005-1009.
[22] Khan M A,Ungar I A.Alleviation of seed dormancy in the desert forbZygophyllumsimplexL.from Pakistan[J].Annals of Botany,1996,80:395-400.
[23] 宋彬,朱小虎,韩娜,李芳,黄俊华.温度及盐胁迫对新疆两种锦鸡儿种子萌发的影响[J].新疆农业大学学报,2011,4(2):119-124.
(责任编辑 武艳培)
Effects of salt and temperature onTageteserectaseed germination
LIU Min1,2, LI Yue2, MAO Zi-jun1, LIU Long2
(1.Key Laboratory of Forest Plant Ecology, Ministry of Education,Northeast Forestry University, Harbin 150040, China;2.College of Life Science and Agriculture Forestry, Qiqihar University, Qiqihar 101006, China)
The two-factor interaction design was employed to explore the effects of different salt concentrations and temperatures onTageteserectaseed germination. The results showed thatT.erectaseed germinated well at saltless and low salt concentration (2 g·L-1) salinity environment which adapt to a wide temperature range. However,T.erectaseed germination was significantly (P<0.05) inhibited at high salt concentrations. There was significant interaction (P<0.05) between salt stress and temperature. The salt concentration affected the impacts of temperature on the germination and seedling growth. There was no significant difference between the germination of three temperature treatments at saltless and low salt concentration environment. The germination percentages, seedling height and root length of high temperature treatment were significantly lower (P<0.05) than those of the other two temperature treatments when the salt concentration were 4 and 6 g·L-1. The temperature also affected the impacts of salt concentration on the germination and seedling growth. At 15/25 ℃ and 20/30 ℃ treatments, the germination percentage significantly decreased (P<0.05) when the salt concentration was greater than or equal to 6 g·L-1. While at 25/35 ℃ treatment, it was significantly decreased (P<0.05) when the salt concentration was greater than or equal to 4 g·L-1. The germination index was more sensitive to salinity stress than the germination percentage.
Tageteserecta; seed germination; seedling growth; interaction; salt injury index
MAO Zi-jun E-mail:zijunm@126.com
10.11829j.issn.1001-0629.2013-0440
2013-08-02 接受日期:2014-03-06
齐齐哈尔市生态园林城市建设中的城市生态环境建设研究(11531427)
刘敏(1979-),女,湖南长沙人,副教授,在读博士生,主要从事植物生态研究。E-mail:liuminaa168@126.com
毛子军(1958-),女,黑龙江齐齐哈尔人,教授,博士,主要从事植物学及生态学研究。E-mail: zijunm@126.com
Q945.35
A
1001-0629(2014)06-1104-06*1
