我国马铃薯甲虫主要生物学、生态学技术研究进展及监测与防控对策
2014-08-10郭文超吐尔逊程登发谭万忠张正坤李国清姜卫华邓建宇吴家和邓春生刘小侠0吕和平
郭文超,吐尔逊,程登发,谭万忠,张正坤,李国清,姜卫华,邓建宇,吴家和,邓春生,李 晶,刘小侠0 ,吕和平
(1.新疆农业科学院植物保护研究所/农业部西北荒漠绿洲作物有害生物综合治理重点实验室,乌鲁木齐 830091;2.中国农业科学院植物保护研究所,北京 100193;3.西南大学植物保护学院,重庆 400716;4.吉林省农业科学院植物保护研究所,公主岭 136100;5.南京农业大学植物保护学院,南京 210095;6.浙江农林大学,临安 311300;7.中国科学院微生物研究所,北京 100101;8.中国农业科学院农业环境与可持续发展研究所,北京 100081;9.新疆维吾尔自治区植保站,乌鲁木齐 830006;10.中国农业大学,北京 100193;11.甘肃省农业科学院植物保护研究所,兰州 730070)
特约稿件
我国马铃薯甲虫主要生物学、生态学技术研究进展及监测与防控对策
郭文超1*,吐尔逊1,程登发2,谭万忠3,张正坤4,李国清5,姜卫华5,邓建宇6,吴家和7,邓春生8,李 晶9,刘小侠10,吕和平11
(1.新疆农业科学院植物保护研究所/农业部西北荒漠绿洲作物有害生物综合治理重点实验室,乌鲁木齐 830091;2.中国农业科学院植物保护研究所,北京 100193;3.西南大学植物保护学院,重庆 400716;4.吉林省农业科学院植物保护研究所,公主岭 136100;5.南京农业大学植物保护学院,南京 210095;6.浙江农林大学,临安 311300;7.中国科学院微生物研究所,北京 100101;8.中国农业科学院农业环境与可持续发展研究所,北京 100081;9.新疆维吾尔自治区植保站,乌鲁木齐 830006;10.中国农业大学,北京 100193;11.甘肃省农业科学院植物保护研究所,兰州 730070)
本文回顾和分析了马铃薯甲虫在世界及我国的传播扩散情况和趋势,重点介绍了马铃薯甲虫的主要生物学和生态学特性,在我国发生现状、防治技术研究进展与应用等,针对我国马铃薯甲虫扩散趋势和监测防控中面临的问题,提出遏制马铃薯甲虫在我国进一步传播扩散和危害的监测与防控对策。
马铃薯甲虫; 分布与危害; 入侵生物学; 生态适应机制; 防治技术; 监测防控对策
马铃薯甲虫[Leptinotarsadecemlineata(Say)]是国际公认的马铃薯重要毁灭性害虫,隶属于鞘翅目(Coleoptera)叶甲科(Chrysomelidae)。作为我国重要的外来入侵物种之一和对外重大检疫对象,马铃薯甲虫被列入《中华人民共和国进出境植物检疫有害生物名录》[1]和“中国最具危险性的20种外来入侵物种”名单[2]。该虫对马铃薯的危害通常十分严重。据不完全统计,在世界范围内每年因马铃薯甲虫的为害造成马铃薯产量损失超过50亿美元,严重威胁着世界马铃薯生产和生态安全[3]。马铃薯作为世界性重要的粮菜兼用作物和工业原料,在全球主要粮食作物中,仅排在玉米、小麦和水稻之后,位居第四。联合国粮农组织以1999年-2001年的价格为基数测算,马铃薯的价格分别是水稻的1.08倍,玉米的1.09 倍和小麦的2.32倍,马铃薯生产产值和效益高于其他粮食作物。进入21世纪,我国马铃薯生产增长迅速,到2009年马铃薯种植面积508万hm2,面积和总产分别占全世界的27%和22%,以原粮计算,马铃薯占国内粮食总产高达13.3%,中国成为世界最大的马铃薯生产国。2013年我国马铃薯种植面积接近666.7万hm2,面积和总产仅次于水稻、玉米和小麦,为我国第四大粮食作物。由此可见,马铃薯产业已成为我国农村经济发展的一个重要支柱产业[4]。在此背景下,针对我国马铃薯甲虫发生现状和传播趋势,进一步有效控制马铃薯甲虫在我国的危害和蔓延,对于保障马铃薯生产安全和生态安全意义重大。本文在回顾和分析马铃薯甲虫在世界及我国的传播扩散情况,介绍马铃薯甲虫的主要生物学、生态学,在我国发生现状、防治技术研究进展与应用等基础上,通过分析我国马铃薯甲虫扩散蔓延趋势及监测防控面临的问题,提出有效遏制马铃薯甲虫在我国进一步传播、危害的监测和防控对策,以期对未来我国马铃薯监测和防控提供参考。
1 分布与危害
1.1 发生与分布范围
马铃薯甲虫原产于美国落基山山脉东坡,首次在野生杂草刺萼龙葵(SolanumrostratumDunal)上发现,1824年记述为新种。1855年首次报道了马铃薯甲虫作为农作物害虫在美国科罗拉多州马铃薯产区造成严重危害,故此又称该虫“科罗拉多马铃薯甲虫”(Colorado potato beetle, CPB)。此后马铃薯甲虫每年以85 km的速度向东扩散,1875年传播到大西洋沿岸,并向周边国家传播,相继传入加拿大、墨西哥,19世纪70年代通过人为因素传播到欧洲西部的德国、英国和荷兰,但是通过检疫封锁措施,得到有效控制。1918年-1920年第一次世界大战后又经波尔多进入法国,此后分三路向东扩散,不久在捷克、斯洛伐克、克罗地亚、匈牙利、波兰等东欧国家定居。20世纪50年代侵入原苏联边境,60年代传入原苏联欧洲部分,1975年传入里海西岸,20世纪80年代继续向东蔓延至中亚各国,于20世纪90年代初传入我国新疆。在世界范围内,目前马铃薯甲虫分布于美洲大陆15~55 °N之间与欧亚大陆33~60 °N之间的欧洲、亚洲和北美洲,以及非洲北部的 40 多个国家和地区(图1)。
马铃薯甲虫于1993年5-7月在我国伊宁市、察布查尔县和塔城市首次发现。在过去的20年马铃薯甲虫不断沿天山北坡向东扩散蔓延,传播直线距离超过了800 km,每年扩散速度平均约为80 km。2003年传播至昌吉回族自治州木垒哈萨克自治县。截止到2013年,马铃薯甲虫分布扩展至新疆天山以北准噶尔盆地的伊犁河谷地区(察布查尔锡伯族自治县、霍城县、伊宁市、伊宁县、特克斯县、昭苏县、巩留县、尼勒克县、新源县)、塔城地区(塔城市、额敏县、裕民县、托里县、沙湾县、乌苏市)、阿勒泰地区(仅分布于哈巴河县、福海县、吉木乃县、布尔津县、阿勒泰市)、博尔塔拉蒙古自治州(精河县、博乐市、温泉县)、奎屯市、石河子市、五家渠市、昌吉回族自治州(玛纳斯县、呼图壁县、昌吉市、阜康市、奇台县、吉木萨尔县、木垒哈萨克自治县)、乌鲁木齐市(新市区、沙依巴克区、米东区、乌鲁木齐县)、巴音郭楞蒙古自治州(仅分布于和静县林场)等7个地(州、市)的38县(市、区)约30万km2的区域,以及上述区域新疆建设兵团所属团场马铃薯种植区。目前马铃薯甲虫分布的最东端在我国新疆天山以北昌吉回族自治州木垒县博斯坦乡三个泉子村(90°41′15″E,43°44′20″N,海拔1 465 m),距新疆与甘肃省交界处550 km。除新疆天山以北的大部分区域外,2013年7月在我国吉林省延边朝鲜族自治州珲春市春化镇发生马铃薯甲虫疫情。马铃薯甲虫在新疆天山以南喀什地区、和田地区、阿克苏地区、巴音郭楞蒙古自治州(和静县除外)、东疆的吐鲁番地区和哈密地区,以及我国其他省(自治区、直辖市)和台湾地区均未发生(图1)。
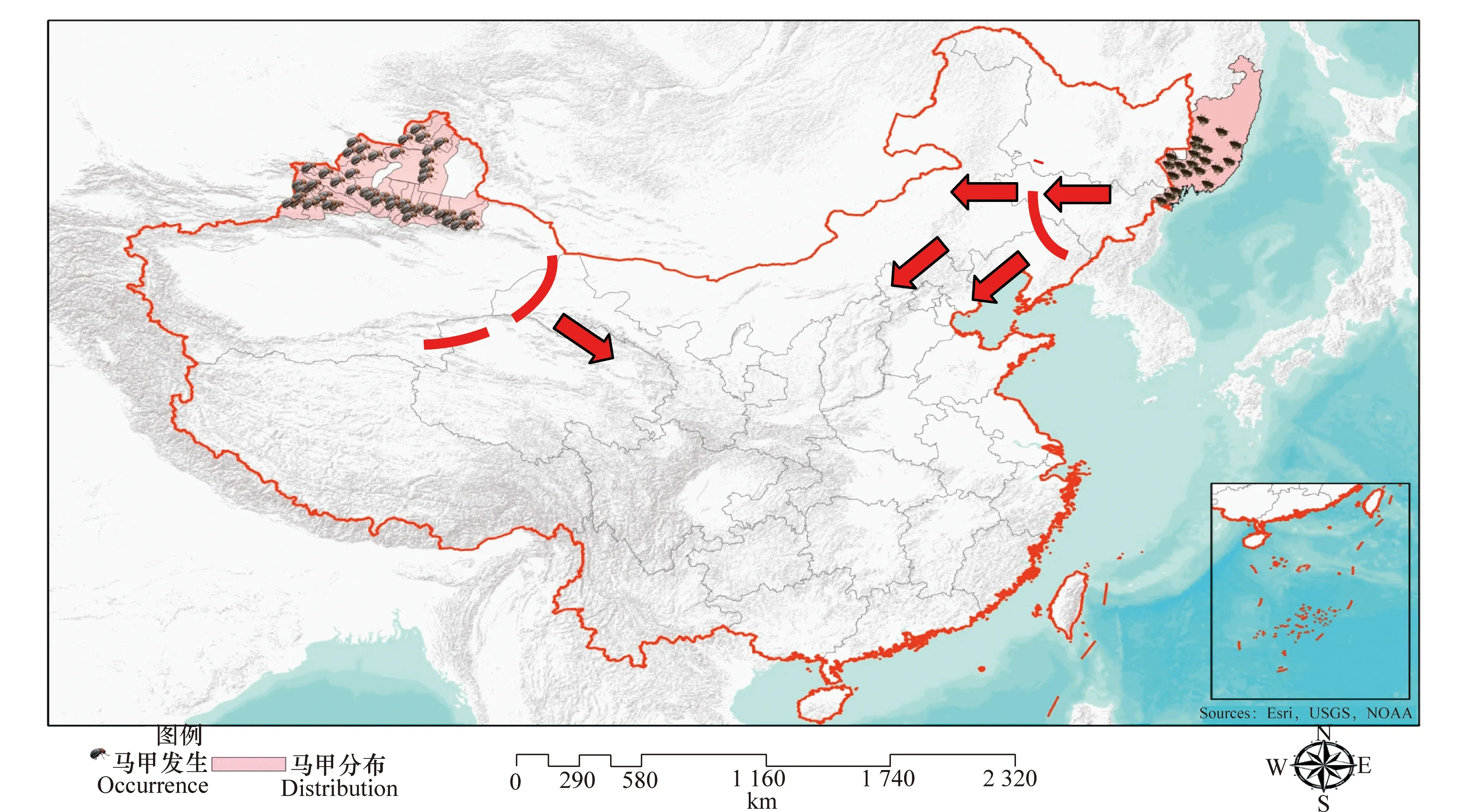
图1 马铃薯甲虫在我国分布区域及可能传播路线图Fig.1 Distribution and possible transmission route of Colorado potato beetle in China
1.2 取食特点及危害
马铃薯甲虫以成虫、幼虫取食为害马铃薯、茄子等寄主作物的叶片和嫩尖。其成虫及3~4龄幼虫取食量较大,在我国发生区,马铃薯甲虫为害初期常使叶片出现大小不等的孔洞或缺刻,其继续取食可将叶肉吃光,留下叶脉和叶柄。成虫和幼虫严重为害时常将叶柄或较细的幼茎咬断,从而引起整个叶片或茎上部分叶片枯死。这种情况通常发生在开花前受害严重的马铃薯植株上,在夏季强烈阳光的照射下,被害马铃薯植株剩余的茎秆会在几天内失水干枯死亡。因此,在马铃薯幼苗、开花至薯块形成期遭受马铃薯甲虫严重为害会对产量造成很大影响。成虫、幼虫喜食幼嫩的中上部叶片,形成“秃顶”后,幼虫向下转移为害,待整株叶片都吃光后,再向邻近植株转移为害。马铃薯甲虫在食物条件匮乏情况下也取食马铃薯块茎,有时还偶食茄子和曼陀罗的果实,以及番茄茎秆韧皮部和白菜等十字花科植物。同时,其危害可传播马铃薯褐斑病、环腐病等多种病害(图2、图3)。

图2 马铃薯植株被害状Fig.2 Damage symptoms of potato plant

图3 茄子植株被害状Fig.3 Damage symptoms of egg plant
马铃薯甲虫是世界范围内马铃薯主要产区最具毁灭性的食叶害虫,在过去的150年里其不断传播扩散,所到之处给当地马铃薯生产造成了严重损失。根据国外相关文献报道和在我国马铃薯甲虫发生区对其为害的调查,如果不进行合理防治,马铃薯甲虫为害一般可造成30%~50% 的产量损失, 严重者减产可达90%,甚至造成绝收。此外,在我国马铃薯甲虫2代发生区研究发现,其为害造成的马铃薯产量损失随虫口密度的增加而逐渐增大。第1代马铃薯甲虫幼虫田间密度与马铃薯产量损失的关系符合模型Y=-2.514 3+2.377 1X(r=0.922 9**)。在第1代马铃薯甲虫低龄幼虫(1~2龄)虫口密度达到5头/株,可造成14.9%的产量损失;当马铃薯甲虫低龄幼虫虫口密度为20头/株,导致的产量损失可达60%以上。据不完全统计,仅美国每年因马铃薯甲虫为害造成的经济损失为41.4亿~69亿美元,而全世界每年因该虫危害损失高达50亿~100亿美元[3,5-8]。
2 入侵生物学及其生态学适应机制
2.1 入侵生物学
2.1.1 生活史
在美洲和欧洲马铃薯甲虫每年发生1~3代。在我国,马铃薯甲虫一年可发生1~2代,以2代为主,个别区域可发生不完全3代。马铃薯甲虫以成虫越冬,其越冬场所为寄主田的土壤中,尤以马铃薯、茄子田为主要越冬场所,与其寄主田邻近的作物田或荒地、林地亦有少数成虫越冬[3,6,9-10](图4)。
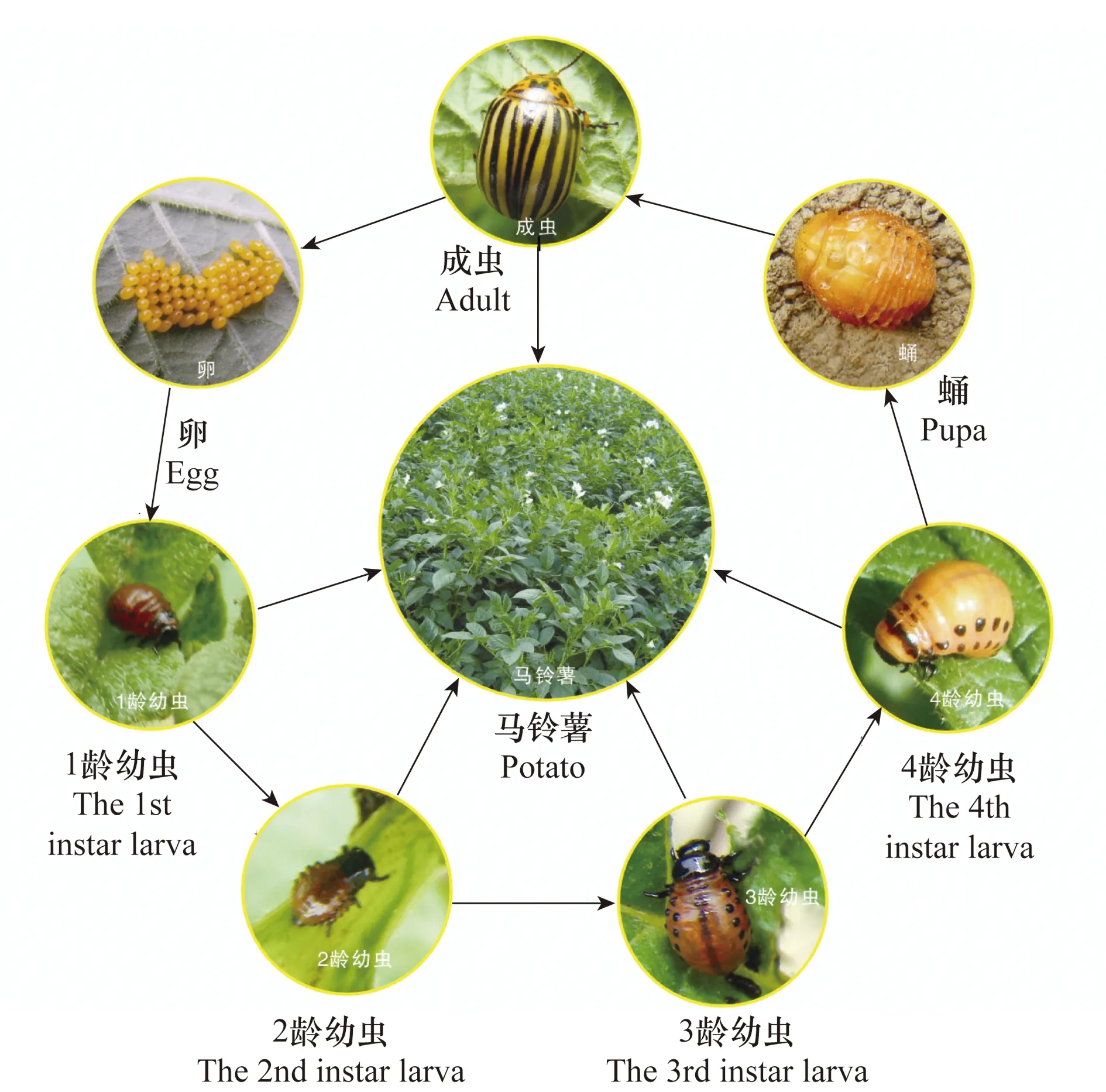
图4 马铃薯甲虫生活史及各虫态Fig.4 Life cycle and every development stage of Colorado potato beetle
2.1.2 寄主范围
马铃薯甲虫的寄主范围相对较窄,属于寡食性昆虫。其寄主主要包括茄科20多个种,多为茄属(SolanumLinn.)的植物,包括马铃薯(SolanumtuberosumLinn.)、茄子(S.melongenaLinn.)等寄主作物,和刺萼龙葵(或称黄花刺茄)(S.rostratumDunal)、欧白英(S.dulcamaraLinn.)、狭叶茄(S.angustifoliumMiller)等茄属野生寄主植物;而茄属的S.carolinense、S.sarrachoides和S.elaeagnifolium等野生植物只是偶尔被取食。马铃薯甲虫寄主还包括菲沃斯属的天仙子(HyoscyamusnigerLinn.);颠茄属的番茄(LycopersiconesculentumMill.)。在我国新疆发生区,马铃薯甲虫寄主有10余种。主要包括马铃薯、茄子、天仙子、刺萼龙葵、番茄、龙葵(S.nigrumLinn.)。此外,马铃薯甲虫偶食曼陀罗属的曼陀罗(DaturastramoniumLinn.)和十字花科的白菜(BrassicapekinensisRupr.)等个别植物,在我国吉林珲春及其以东的区域寄主分布,尤其是野生寄主的分布尚待进一步明确[3,10-12]。
2.1.3 传播扩散
马铃薯甲虫主要以成虫飞行后随气流裹挟进行远距离传播扩散,也可随发生区的马铃薯薯块、蔬菜等相关农副产品以及交通工具等传播,有时可通过水流传播[3,7,13]。
2.1.4 发育起点与有效积温
不同地理种群马铃薯甲虫的发育起点和有效积温有所不同。在我国二代发生区(乌鲁木齐市安宁渠),马铃薯甲虫卵、幼虫、蛹发育起点温度分别为9.14、9.59 和10.23 ℃,有效积温依次为73.26、273.02和100.38日·度[3,14]。
2.1.5 滞育
马铃薯甲虫具有兼性滞育的习性,属于短日照滞育型,成虫感受到临界短日照后进入滞育,环境温度过高和食物质量较差会对滞育产生一定影响。在欧洲和北美洲不同地理纬度的马铃薯甲虫种群临界光周期不同,一般南部地理种群比北部种群所需的光周期短[5]。研究表明我国新疆石河子地区马铃薯甲虫种群的临界光周期为14 h。第1、2代和少数越冬成虫产卵后在短日照(<14 h)时可引起甲虫滞育[3]。
2.2 生态适应机制
大量研究表明,马铃薯甲虫在过去的一个多世纪里持续不断地传播和暴发危害,使其从干旱的亚热带地区扩展到温带的北限区域(接近北纬60°),并成为栽培马铃薯的国际性重大害虫,这与其拥有极强的生理、生态适应能力关系密切[6,12,14-15]。
2.2.1 适应不良环境的生物学机制
马铃薯甲虫具有多种抵御并适应不良环境的生物学特性和近乎完美的生存对策,如高繁殖力、滞育和远距离飞行等。在我国对马铃薯甲虫研究表明,一般单头雌虫一生产卵量为300~3 130粒,平均约1 000粒。越冬代成虫在入土前进行交尾。换言之,只要有一头雌虫在春季传播到新的栖息地,补充营养产卵繁殖后,即可建立一个新的种群。即使在无寄主(马铃薯、茄子、天仙子)存在的情况下,大部分越冬成虫亦可存活超过30 d,这种较强的耐饥饿能力,保证了越冬后的成虫寻找适宜的生存环境所需的时间。此外,马铃薯甲虫具有较强的飞行能力也是其适应不良环境的对策之一,研究表明随气流远距离传播则是马铃薯甲虫主要的传播扩散方式。回顾马铃薯甲虫传播历史,在过去的一个多世纪,马铃薯甲虫从原产地持续不断扩散。例如:马铃薯甲虫在1859年前后美国内布拉斯加州栽培马铃薯上暴发以后,其以每年185 km的速度迅速向东传播,十几年行程3 000 km,于1875年传播到美国东部的大西洋沿岸地区。再例如:1920年马铃薯甲虫在法国波尔多成功登陆,在随后的60年马铃薯甲虫持续向东传播扩散的脚步从来没有停歇过,以每年100~200 km的速度自西向东传播,60年扩展蔓延6 020 km,到1979年蔓延至中国边界地带。1993年我国新疆伊犁河谷地区和塔城地区与哈萨克斯坦接壤的边境区域首次发现马铃薯甲虫以来,该虫持续不断地向东持续传播,到2003年向东沿天山北坡扩散至昌吉回族自治州木垒县博斯坦乡三个泉子村,10年马铃薯甲虫传播扩散了约800 km。因此,马铃薯甲虫不仅可通过高繁殖率应对不良环境,还可通过滞育和远距离飞行避开不良环境。就其生物学特征而言,显示出马铃薯甲虫极强的环境适应性和生态可塑性[3,6,10,16]。
2.2.2 遗传变异性较强
国内外大量研究表明,马铃薯甲虫不同地理种群存在明显的遗传变异性。例如:研究发现新入侵的地区环境条件不利于马铃薯甲虫发育时,该虫可通过滞育等一系列生物学机制适应不良环境,如反复滞育(包括夏滞育)或延长滞育时间。不同地理种群的临界光周期随着纬度变化而变化,纬度越高,诱发滞育的临界光周期越长。这种滞育光周期的多样性与纬度的光周期变化相适应,使得马铃薯甲虫在不同的环境条件下顺利完成生活史。同时,温度通过调节马铃薯甲虫对光周期的反应而影响滞育。高温可降低马铃薯甲虫对光的敏感性,这使得该虫在较热地区更短的光周期条件下也能诱发滞育[3,12,17-18]。
2.2.3 极易产生抗药性
马铃薯甲虫的抗药性也是反映其生态适应性的重要指标之一。国外马铃薯甲虫抗药性的研究显示,马铃薯甲虫对化学农药的抗药性发生极快,对于新注册的化学农药往往只需2~4年就可产生抗药性,抗药性的严重发生始终困扰着该虫防治[6,21]。在过去近20年里我国马铃薯甲虫的抗药性发展很快,大部分种群对拟除虫菊酯类(氯氟氰菊酯、溴氰菊酯)杀虫剂普遍产生中到极高水平的抗性;对氨基甲酸酯类药剂(丁硫克百威、克百威)为中到高水平抗性;对新烟碱类药剂初现抗性,部分种群对噻虫嗪产生了低到中等水平抗性。另据在我国发生区对马铃薯甲虫抗药机制的研究发现,钠离子通道基因的点突变导致了对高效氟氯氰菊酯的高水平抗性,而乙酰胆碱酯酶基因的点突变引起对克百威的抗性[20-23]。因此,马铃薯甲虫具有这种超强的抗药性适应机制与其遗传变异性有着十分密切的关系。
2.2.4 自然控制作用较弱
虽然国外报道捕食性天敌种类多达200余种,包括白僵菌、绿僵菌等昆虫病原性真菌,以及个别昆虫病原性细菌和线虫,但总体而言,自然控制作用不强[13,24-27]。而在我国发生区,研究发现目前马铃薯甲虫捕食性天敌仅有54种,昆虫病原真菌和细菌分别有4种和2种。天敌资源种类相对有限,且缺乏专一性的优势天敌,自然天敌控制效应较弱[3,28]。这些因素为马铃薯甲虫适应环境和暴发成灾提供了可能。
3 监测与综合防控技术
从2008年以来,在我国马铃薯甲虫疫区(新疆天山以北的区域)和高风险区(东北地区黑龙江省、吉林省和辽宁省),以及西南地区的重庆市、贵州省、广西壮族自治区和四川省、云南省,以及西北地区的甘肃省、青海省和宁夏回族自治区、内蒙古自治区等开展大规模监测,累计监测面积超过213.3万 hm2,为有效地监控马铃薯发生提供了有力的保障。同时,在借鉴国外相关先进技术成果的基础上,结合我国马铃薯生产实际,在研究、试验和示范基础上,通过对研究的技术进一步优化和集成,我国科技工作者研究提出了适宜我国马铃薯甲虫发生区生产实际、操作性强的由与环境相容的化学防治、生物防治、物理防治、生态调控和防虫栽培等关键技术组成的马铃薯甲虫持续防控和应急防控技术。这一技术成果在新疆马铃薯甲虫发生区伊犁河谷地区、塔城地区和阿尔泰地区、昌吉回族自治州西部等地累计推广超过13.3万 hm2,防效达到90.0%以上。而且,新疆植保工作者利用自然地理优势条件开展了科学防控,在马铃薯甲虫发生前沿建立了“马铃薯甲虫阻截带”等。通过上述工作,20年来成功地将马铃薯甲虫控制在天山以北、昌吉回族自治州木垒哈萨克自治县博斯坦乡三个泉子村以西的局部区域,有效遏制了马铃薯甲虫持续向东传播蔓延的脚步,有力保障了我国马铃薯生产的安全,成为我国有效控制农业重大外来入侵的成功案例[3,7,26]。
3.1 监测技术
在寄主植物生长期,在马铃薯、茄子等寄主作物和刺萼龙葵、天仙子等马铃薯甲虫野生寄主植物分布区,以及农产品运输、储藏、加工场所和周围地区,无论发生区和非发生区,每年定点调查2次,第一次在越冬代成虫出土后(5月-6月),第二次在越冬代成虫入土前(7月-8月)。监测区采取对角线式或棋盘式取样方法取样。监测区内4 hm2以下地块取10个调查点,每个点调查10株;4 hm2以上地块取20个调查点,每个点调查5株。记录每株植物上马铃薯甲虫卵、幼虫和成虫数量(卵记录卵块数量),未发生区与发生区监测点数量按寄主植物分布区,以县级行政区域为单位设立4~10个监测点。在未发生区若发现疑似虫体,立即做好标记,记录调查情况,扩大调查范围(半径10 km),将疫情及时上报,并采用应急、扑灭和封锁技术,有效控制马铃薯甲虫的危害和传播扩散[3,29-30]。
3.2 植物检疫措施
植物检疫是控制和预防马铃薯甲虫的有效手段之一。英国1877年颁布了世界上第一部植物检疫法规《毁灭性害虫法》,其主要针对马铃薯甲虫。1933年正式颁布了《马铃薯甲虫法》,并于1955年进行了修订,这也是世界上唯一针对马铃薯甲虫制定的检疫法规,提出了加强进口相关农产品的入境检疫措施,建立了一整套切合实际的马铃薯产区的疫情监测、疫区划定和封锁防控工作流程和与此有关的组织协调机构,通过努力,成功铲除了马铃薯甲虫的多次入侵,马铃薯甲虫始终未能在英国建立种群并定居[11]。根据我国国情和马铃薯甲虫发生特点,提出了具体的植物检疫措施。主要包括一旦局部地区发生马铃薯甲虫疫情,应在当地行政主管部门的监督和指导下将该区域划为疫区;制定行之有效的防控预案;采取积极的检疫、封锁和应急扑灭措施等响应机制;防止和杜绝马铃薯甲虫的进一步传播等内容[3,29]。
近年来,在我国马铃薯甲虫可能传入的风险区域,在植物检疫部门的协助下,对来源于疫情发生地区或国家的相关农产品开展了马铃薯甲虫货运检疫工作。2006年以来,在新疆维吾尔自治区与甘肃省接壤的312国道距哈密市200 km处星星峡联合检疫检查站,对新疆运往我国其他省份的相关农产品实施检疫检验总计184 964批次,300 239 t,多次截获马铃薯甲虫,先后销毁或退回14批次感染或疑似感染马铃薯甲虫的货物。2008年以来,在黑龙江、吉林和内蒙古自治区的主要口岸都设立了检疫监测点,分别对相关进出口货物1 130批次、460批次和500批次进行了检疫检验。西南地区在重庆、贵州、云南和广西对重要机场、港口、车站和集市等处实施货运检疫,共检疫检验货物4 769批次,27.4万t。上述检疫工作对有效阻止和杜绝马铃薯甲虫的传播和扩散,确保我国西北、东北和西南等马铃薯种植区的生产和生态安全发挥了重要作用[3]。
3.3 生态调控技术
国内外相关研究表明,秋耕冬灌、与小麦等禾本科或豆科作物轮作是控制马铃薯甲虫的有效措施,可恶化马铃薯甲虫的生存环境,有效压低越冬基数,减轻越冬代马铃薯甲虫危害。在我国发生区,马铃薯田经秋翻马铃薯甲虫越冬死亡率33.3%~76.36%,秋翻田越冬死亡率比未翻田平均提高约50%。轮作田越冬代马铃薯甲虫种群密度比连作田低29.4%~84.3%,平均达到65.4%[5,26]。
3.4 生物防治技术
国外已经报道的天敌包括昆虫、两栖动物、蜘蛛、线虫和真菌等200多种,以捕食性天敌为主。其中二点益蝽[Perillusbioculatus(Fab.)]和斑腹刺益蝽[Podisusmaculiventris(Say)]等天敌资源应用最为广泛。在病原微生物方面,球孢白僵菌[Beauveriabassiana(Bals.)Vuill]和苏云金杆菌(Bacillusthuringiensis)应用最多[13,24-25,27]。目前我国发生区马铃薯甲虫天敌有54种。其中中华通草蛉[Chrysoperlasinica(Tjeder)]、蓝蝽(ZicronacaeruleaLinnaeus)、多异瓢虫[Adoniavariegata(Goeze)]、蠋蝽[Armacustos(Fabricius)]等天敌捕食效应相对较强,可充分保护自然天敌发挥其控害作用。此外,我国科技工作者开发的300亿孢子/g球孢白僵菌可湿性粉剂和100亿孢子/g油悬浮剂在生产上应用也取得了较好的防治效果。即在1~2龄幼虫期,使用高效白僵菌可湿性粉剂1 500~3 000 g/hm2,喷雾防治2~3次,间隔7 d;或低量喷雾100亿孢子/g油悬浮剂15 000 mL/hm2,共施2次,间隔7 d。该项技术产品在我国马铃薯甲虫发生区应用面积累计达213.3 hm2,平均防效可达72%~91%[3,28,31]。在昆虫细菌杀虫剂的开发利用方面,目前我国科研工作者利用分子生物工程技术手段对Bt杀虫蛋白基因进行了修饰,构建了针对鞘翅目甲虫的高效毒蛋白Bt基因,研制出马铃薯甲虫的工程菌制剂[32],该菌剂喷施80 g/667 m2药后10 d田间应用效果达99.30%[26,28]。上述研制的白僵菌剂和Bt工程菌剂已经进入农药登记程序,显示了良好的应用前景。
3.5 环境友好型化学防治技术
目前国内外马铃薯甲虫的防治主要采用化学防治技术[5,19,26]。一般而言,越冬代成虫和第1代低龄幼虫发生期是防治关键时期,第1代马铃薯甲虫越冬成虫和第一代低龄幼虫(1~2龄)防治经济域值分别为24头/百株和106头/百株,当田间虫口密度达到或超过经济阈值时,可选择高效、低毒和低污染或无污染环境友好型杀虫剂喷雾防治。如使用12.5%高效氯氰菊酯1 500倍液、5%噻虫嗪水分散粒剂90 g/hm2、70%吡虫啉水分散粒剂30 mL/hm2、3%啶虫脒乳油225 mL/hm2、20%啶虫脒可溶性液剂150 g/hm2、3%甲氨基阿维菌素苯甲酸盐微乳剂900 g/hm2、20%氯虫苯甲酰胺悬浮剂150 g/hm2、40%氯虫·噻虫嗪水分散粒剂150 g/hm2、14%氯虫·高氯氟微囊悬浮-悬浮剂150 g/hm2、22%噻虫·高氯氟微囊悬浮-悬浮剂150 g/hm2和60 g/mL乙基多杀菌素悬浮剂750 g/hm2等杀虫剂15 d防效可达90%以上。目前,由于我国对马铃薯甲虫防治仍然主要采取化学防治技术,针对马铃薯甲虫极易产生抗药性这一特点,在实际防治中,应重视合理用药,采用有机磷类、烟碱类、菊酯类、生物源类、昆虫生长调节剂类等不同杀虫机制农药品种的交替使用防治马铃薯甲虫,可有效避免或延缓抗药性的产生和发展。同时,针对马铃薯甲虫防治,我国科技工作者研制出了新型薯块专用种衣剂3.2%甲·噻悬浮种衣剂(获得国家专利:ZL.200710145394.X)在马铃薯播种期,按药种比1∶80包衣种薯,试验示范表明60 d防效可达70%以上,可有效控制越冬代成虫和第1代幼虫的危害[3,26,33]。
3.6 防虫栽培技术
合理的施肥和管理可有效提高马铃薯的耐害性。在我国马铃薯甲虫发生区,研究表明:一般中等肥力土壤投入各种肥料总量分别为氮肥25 kg/667 m2+磷肥15 kg/667 m2+钾肥15 kg/667 m2,其中60%氮肥、60%磷肥和30%钾肥作基肥,播种前施用;40%氮肥、40%磷肥和70%钾肥在开花前期作追肥。该施肥技术可显著提高马铃薯的耐害性,降低马铃薯甲虫为害造成的产量损失[34]。该技术在新疆伊犁河谷的特克斯县示范,与常规防治田相比,马铃薯甲虫为害率平均降低16%,产量平均提高17.7%,肥料利用率提高12.5%。此外,采用覆膜有效提高了春季地温,促进马铃薯生育进程,同时覆膜能够有效降低无效蒸腾,提高了马铃薯对水肥的利用效率。该技术在新疆伊犁河谷特克斯县累计应用面积达到69.3 hm2,虫口减退率达62%,增产24.8%。其防虫机理在于覆膜影响马铃薯甲虫幼虫化蛹,减少第2代马铃薯甲虫田间种群数量,减轻了危害,是一种重要的物理防治措施之一。而且,研究发现,在覆膜条件下大量元素配施适宜的Zn(硫酸锌)、B(硼砂)等微量元素可以显著提高马铃薯的耐害性[3,34]。
3.7 种植诱集带
国内外研究表明:利用马铃薯、茄子等寄主作物对马铃薯甲虫有明显的引诱作用,提早种植马铃薯诱集带(面积不低于种植面积1%),随后集中杀灭越冬成虫[3,26],可有效控制越冬代成虫的危害。此项技术作为一项重要的防控技术,通常应用在发生的前沿地带。
3.8 人工捕捉
利用马铃薯甲虫成虫具有“假死性”的习性,于越冬成虫出土期,在田间定期(1~2次/周)捕捉越冬成虫,摘除卵块,带出田外集中销毁,可有效压低虫口基数,减轻危害[3,24,26]。虽然这一方法较为传统,但是目前仍然广泛用于马铃薯甲虫的防治。
3.9 新型化学防治替代技术的应用
近年来,针对马铃薯甲虫抗药性治理这一世界性难题,国内外相继开发了多种新型化学防治替代技术。如开发了抗虫转基因(cry3A、cry1B3)马铃薯品系[3,35-38],研制了基于昆虫化学信息生态学的聚集素引诱剂和植物引诱剂等,该项技术在我国马铃薯甲虫发生区和风险区推广应用面积达66.7 hm2[3,39-43]。此外,我国科技工作者在国内外首次发现了与马铃薯甲虫保幼激素合成相关的致死基因,克隆了保幼激素合成和代谢相关基因片段,并构建双链RNA表达载体(pET-2P-X),转入大肠杆菌成功表达,其中含腺苷高半胱氨酸水解酶dsRNA的菌液表现出明显致死作用,室内12 d后幼虫死亡率可达70%以上。这一研究成果为进一步应用RNA干扰技术开发出具有高效、低毒、专一性强的“基因农药”奠定了基础[22,44]。
那木偶没多大事,只脸上被新漆过的铁架子剐蹭了一些漆渍上去。王爷的脸却变成了猪肝色,一边嘴里咕咕叨叨骂起来,一边从口袋里掏出平日里擦嘴用的手帕,沾了些唾沫,使劲在那木偶脸上擦。见擦不干净,又从口袋里掏出一个小刀(不知道还能掏出啥来),把着力度轻轻地刮,竟然那漆渍就被刮掉了。
4 我国马铃薯甲虫的监测和防控对策
4.1 我国马铃薯甲虫的监测防控形势
作为世界公认的马铃薯毁灭性害虫,马铃薯甲虫传入我国新疆20年来,虽然一直被控制在天山以北昌吉回族自治州木垒县博斯坦乡3个泉子村以西的局部区域,但自该虫入侵以来,其由西向东传播,疫区面积扩大了数百倍,马铃薯甲虫进一步向东越过新疆东部传播扩散至我国内蒙古和甘肃的风险依然存在。
在我国东北区域,马铃薯甲虫现已蔓延到与俄罗斯的滨海边区西南部毗邻的我国吉林省延边朝鲜族自治州边境地区的珲春市。2013年7-8月份吉林珲春春化镇首次发现马铃薯甲虫疫情,发生地为春化镇桦树村、下帽子村、上帽子村和分水岭村等地,距边境仅约10 km。据调查,疫情点马铃薯被害株既有成虫、也有幼虫和卵,分布于偏僻的林间和农户院落种植的马铃薯地、而且呈零星分布。通过以往的监测和对突发疫情分析,据此判断马铃薯甲虫传入时间应该在2013年5-6月,我们初步得出马铃薯甲虫通过自然扩散传入的可能性较大,目前属于疫情的初发和入侵早期阶段。此外,据悉俄罗斯滨海区已经解除了对马铃薯甲虫的检疫,表明在俄罗斯滨海区马铃薯甲虫已普遍发生。根据马铃薯甲虫发生规律和生物学特性,对目前我国在东北区域马铃薯甲虫发突发疫情进行综合分析,可能与俄方对马铃薯甲虫防控工作不利,在马铃薯生长季节,距离我国边境地区俄方一侧马铃薯甲虫虫口密度较高(造成寄主作物马铃薯受害较重),因食物匮乏引起马铃薯甲虫成虫随即扩散迁飞所致。这也可能与近年来全球气候变暖使得该区域马铃薯甲虫越冬、种群扩张和严重发生并不断扩散有密切关系,这一点尚需进一步研究和验证。因此,未来马铃薯甲虫从俄罗斯滨海区持续传入我国东北边境地区的风险大幅度增加。
近年来我国检疫部门在边境口岸检疫中截获马铃薯甲虫的频率在逐步增加,这充分表明该害虫通过交通工具和相关农副产品传入我国的风险也在加剧,可能对我国马铃薯生产构成威胁。
综上所述,未来我国马铃薯甲虫的监测和防控形势依然十分严峻。
4.2 未来我国马铃薯甲虫的监测和防控对策
首先,未来应继续加强对目前我国疫区的马铃薯甲虫有效监测和防控。其目的是有效遏制马铃薯甲虫从西向东由新疆传入我国西北马铃薯主产区,进而传入我国中部和西南马铃薯主产区。消除或减轻马铃薯甲虫从新疆发生区向甘肃省、青海省、内蒙古自治区等我国西北马铃薯主产区扩散蔓延的风险,仍然是我国马铃薯甲虫监测和防控的重点(图1)。过去20年来,在我国马铃薯甲虫被成功地狙击在新疆现有的发生区,其继续向东扩散的脚步被有效阻止充分证明了未来遏制其从新疆向东传入我国其他马铃薯产区是完全可行的。
其次,应高度重视对我国东北新发现疫情的全面普查和封锁防控工作,并将其常态化,及时准确了解马铃薯甲虫发生情况,并在该区域建立“马铃薯甲虫封锁和防控阻截带”,防止马铃薯甲虫从俄罗斯滨海区自西向东传入我国黑龙江省、吉林省和内蒙古东部,进而南下通过山东省、河北省、河南省等华北地区传入安徽省、江西省、湖北省、湖南省、广西壮族自治区、贵州省、云南省、四川省和重庆市等我国中部和西南马铃薯产区,从而化解或降低马铃薯甲虫从俄罗斯滨海区通过东北传入我国其他马铃薯主产区的风险。为此,在吉林省珲春市、黑龙江省东宁市和绥芬河市等沿俄罗斯滨海区的边境口岸区域应重点加强定期监测,该区域纬度相对较低,热量资源较为丰富(>10 ℃的有效积温超过2 500日·度,马铃薯甲虫可发生2代),有利于马铃薯甲虫的种群发生、增长和扩散,传入的风险较大。建议在上述区域每年5-8月开展马铃薯甲虫监测1~2次。从毗邻我国吉林和黑龙江俄方区域的气候特点分析,由于黑龙江、吉林等我国东北地区农作物生长季节(春末-夏季-秋初)多盛行西北风,而俄方马铃薯甲虫发生区处于珲春市的东侧,马铃薯甲虫大规模从俄方边境地区起飞后逆主要气流方向进行高空远距离传入我国吉林省和黑龙江的可能性极小。从生态地理条件和寄主分布情况分析,珲春市、绥芬河市和东宁市一带多丘陵地带,初步观察和调查吉林珲春市和东宁市以东马铃薯和茄子等寄主作物仅零星种植,面积很小。而俄罗斯滨海区毗邻我国的边境区域马铃薯种面积也十分有限,且多集中在居民住户的房前屋后私家菜地,不具备大范围入侵扩散和危害的条件,仅可能形成随机(和风向无关)、小规模、局部和低空短距离(几百米至十几公里距离)的入侵传播。因此,只要在该区域采取科学有效的措施,全面及时掌握疫情动态,对发现疫情进行及时的铲除等,建立“马铃薯甲虫阻截带”,遏制马铃薯甲虫从俄罗斯滨海区持续传入我国,防止其定殖和进一步扩散是完全可能的(图1)。
此外,应加强宣传和培训,提高疫情发生区和高风险区社会公众对马铃薯甲虫危害性的认识,鼓励当地群众积极参与疫情发现、封锁和铲除工作;对疫情区已经确定疫情点进行定位,通过种植寄主作物马铃薯进行诱集,监控疫情发生情况,并及时铲除疫情。加强外检、内检联合,采取统一联动和信息交流与共享机制,提高马铃薯甲虫监测、疫情处置和口岸检疫的实施效果。而且,应进一步做好吉林省、黑龙江省和内蒙古自治区马铃薯主产区疫情的监测工作,防患于未然。
在现有研究技术成果和防控监测工作经验的基础上,应整合国内现有相关研究力量,进一步增加持续的科技投入,加大马铃薯甲虫监测、防控新技术和产品研究力度,为我国马铃薯甲虫监测防控提供持续有效的技术支撑。
综上所述,对于未来我国马铃薯甲虫监测和防控而言,必须采取有效措施和科学对策,加强我国西北和东北疫情发生区前沿地带的监测和封锁防控工作,继续在东北和西北两个“高风险区”建立“马铃薯甲虫阻截带”。同时进一步开展我国马铃薯甲虫监测和防控新技术研究与应用,并强化相关部门之间的协作和配合,重视我国北方边境地区(高风险区)马铃薯甲虫监测和封锁防控工作的实施,以及我国马铃薯主产区的陆路、航空口岸的检疫工作。通过采取上述对策和措施,在较长一段时期有效遏制马铃薯甲虫在我国进一步的传播和危害,甚至彻底将马铃薯甲虫控制在我国现有发生区,保障我国马铃薯生产安全和生态安全不仅可能,而且完全可行。
[1] 万方浩, 谢丙炎, 褚栋.生物入侵: 管理篇[M].北京:科学出版社, 2008: 297-314.
[2] 万方浩, 郭建英, 张峰.中国生物入侵研究[M].北京: 科学出版社, 2009: 29-100.
[3] 郭文超,谭万忠,张青文.重大外来入侵害虫马铃薯甲虫生物学、生态学与综合防控[M].北京:科学出版社,2013: 2-460.
[4] 谢从华.马铃薯产业的现状与发展[J].华中农业大学学报 (社会科学版), 2012,97(1):1-4.
[5] Casagrande R A.The Colorado potato beetle: 125 years of mismanagement[J].Bulletin of the ESA,1987,33(3):142-150.
[6] Hare J D.Ecology and management of the Colorado potato beetle[J].Annual Review of Entomology,1990,35:81-100.
[7] 郭文超, 吐尔逊, 许建军,等.马铃薯甲虫识别及其在新疆的分布、传播和危害[J].新疆农业科学, 2010, 47(5): 906-909.
[8] 张生芳.马铃薯甲虫[J].植物检疫, 1988, 2(1): 59-68.
[9] Dwyer J D, Dill J F, Carter S.Colorado potato beetle[M].IPM Potato Fact Sheet 101,Cooperative Extension Service,University of Maine at Orono, 2001.
[10]吐尔逊·阿合买提,许建军,郭文超,等.马铃薯甲虫主要生物学特性及发生规律研究[J].新疆农业科学, 2010, 47(6):1147-1151.
[11]Bartlett P W.Preventing the establishment of Colorado beetle in England and Wales[M]∥Ebbels D L,King J E.Plant health, the scientific basis for administrative control of plant diseases and pests, 1979:247-257.
[12]De-Wilde J, Hsiao T.Geographic diversity of the Colorado potato beetle and its infestation[M]∥Lashomb J H,Casagrande H,eds.Advances in potato pest management.Hutchinson Ross, Stroudsburg, 1981:47-48.
[13]张生芳.欧洲的马铃薯甲虫食虫天敌[J].植物检疫, 1994(5): 276-278.
[14]周昭旭,罗进仓,吕和平,等.温度对马铃薯甲虫生长发育的影响[J].昆虫学报,2010, 53(8): 926-931.
[15]张云慧,张智,何江,等.马铃薯甲虫自然种群抗寒能力测定[J].植物保护,2012,38(5):64-67.
[16]Caprio M A,Grafius E J.Effects of light, temperature and feeding status on flight initiation in post diapause Colorado potato beetles (Coleoptera: Chrysomelidae)[J].Environmental Entomology, 1990, 19(27): 281-285.
[17]Tauber M J, Tauber C A, Obrycki J J, et al.Geographical variation in responses to photoperiod and temperature byLeptinotarsadecemlineata(Coleoptera: Chrysomelidae) during and after dormancy[J].Annals of Entomology Society of America, 1988,81(5):764-773.
[18]梁忆冰, 林伟, 王跃进, 等.生态因子在马铃薯甲虫地理分布中的作用[J].植物检疫, 1999, 13(5): 257-262.
[19]Alyokhin A.Colorado potato beetle management on potatoes: current challenges and future prospects[J].Fruit Vegetable Cereal Science and Biotechnology, 2009,3:10-19.
[20]Jiang Weihua, Wang Zhitian, Xiong Manhui, et al.Insecticide resistance status of Colorado potato beetle adults in Northern Xinjiang Uygur Autonomous Region[J].Journal of Economic Entomology, 2010, 103(4):1365-1371.
[21]Jiang Weihua, Xiong Manhui, Wang Zhitian, et al.Incidence and synergism of resistance to conventional insecticides in 4th instar larvae of Colorado potato beetle,Leptinotarsadecemlineata, in northern Xinjiang Uygur Autonomous Region[J].Acta Entomologica Sinica, 2010, 53(12):1352-1359.
[22]卢伟平.几种新型杀虫剂对马铃薯甲虫的毒力及两类靶标的分子克隆[D].南京:南京农业大学,2011:1-111.
[23]王志田, 姜卫华, 李国清.新疆北疆马铃薯甲虫成虫抗药性水平监测[J].农药, 2010, 49(3): 206-208.
[24]Logan P A, Casagrande R A, Hsiao T H,et al.Collections of natural enemies ofLeptinotarsadecemlineata[Coleoptera:Chrysomelidae] in Mexico,1980-1985[J].Entomophaga, 1987, 32: 249-254.
[25]O’Neil R J, Canas L A.Foreign exploration for natural enemies of the Colorado potato beetle in Central and South America[J].Biological Control, 2005, 33(1): 1-8.
[26]郭文超, 吐尔逊, 许咏梅,等.马铃薯甲虫持续防控技术研究与应用[J].新疆农业科学, 2011, 48(2): 197-203.
[27]张生芳.由美洲引入欧洲的马铃薯甲虫天敌[J].植物检疫, 1994(6): 342-344.
[28]郭文超, 邓春生, 谭万忠, 等.我国CPB生物防治研究进展[J].新疆农业科学, 2011, 48(12): 2217-2222.
[29]郭文超.马铃薯甲虫应急防控技术指南[M]∥张国良.农业重大外来入侵生物的应急防控技术指南.北京:科学出版社,2010:422-442.
[30]新疆维吾尔自治区标准计量局.马铃薯甲虫应急扑灭和封锁防控技术规程(DB65/T3398-2012)[S].乌鲁木齐, 2012.
[31]付文君, 何江,米慧,等.6种新型药剂防治马铃薯甲虫田间药效初探[J].新疆农业科学, 2013, 50(8): 1482-1487.
[32]张杰, 宋福平, 黄大昉,等.对鞘翅目害虫高毒力Bt基因cry3Aa7的分离克隆及表达研究[J].中国农业科学, 2002, 35(6):650-653.
[33]郭建国,刘永刚,张海英,等.70%噻虫嗪种子处理可分散粉剂和10%吡虫啉可湿性粉剂拌种对马铃薯甲虫的防效[J].植物保护, 2010,36(6):151-154.
[34]许咏梅,郭文超,谢香文,等.新疆马铃薯甲虫对不同施肥及品种的响应[J].西北农业学报,2011,20(4):179-185.
[35]Adang M J, Brody M S, Cardineau G, et al.The reconstruction & expression of aBacillusthuringiensiscryⅢAgene in protoplasts and potato plants[J].Plant Molecular Biology, 1993, 21: 1131-1145.
[36]Zhou Zhaoxu, Pang Jinhuan, Guo Wenchao,et al.Evaluation of the resistance of transgenic potato plants expressing various levels of Cry3A against the Colorado potato beetle (LeptinotarsadecemlineataSay) in laboratory and field[J].Pest Management Science,2012:1-24.
[37]罗晓丽,杨欣,郭文超,等.转cry3A 基因马铃薯外源蛋白表达和对CPB抗性分析[J].山西农业科学,2010,38(11):3-5.
[38]周壮志, 周永刚, 何朝族,等.cry3A和vhb基因在转基因马铃薯中的表达[J].生物化学与生物物理进展, 2004,31(8):741-751.
[39]Dickens J C.Orientation of Colorado potato beetle to natural and synthetic blends of volatiles emitted by potato plants[J].Agricultural and Forest Entomology, 2000, 2:167-172.
[40]Dickens J C, Oliver J E, Hollister B, et al.Breaking a paradigm: male-produced aggregation pheromone for the Colorado potato beetle[J].The Journal of Experimental Biology, 2002, 205: 1925-1933.
[41]Dickens J C.Plant volatiles moderate response to aggregation pheromone in Colorado potato beetle[J].Journal of Applied Entomology, 2006, 130(1): 26-31.
[42]李源, 赵珮, 尹春艳,等.多种植物挥发物及CPB聚集素对CPB的引诱作用[J].昆虫学报, 2010, 53(7): 734-740.
[43]赵珮.马铃薯甲虫对植物挥发物和聚集素的触角电位反应[J].安徽农业科学, 2011,39(18): 10926-10928.
[44]吕东.马铃薯甲虫脯氨酸脱氢酶基因的克隆、表达分析及其dsRNA的发酵生产[D].南京:南京农业大学,2010:1-88.
致谢
以下各位专家承担了《植物保护》2013年度稿件
审理工作,特此向他们表示衷心感谢!
安玉兴 曹坳程 曹克强 曹学仁 曹雅忠 陈巨莲 陈 君 陈 林 程登发 程兆榜 褚 栋 崔海兰
董金皋 樊美珍 范在丰 封洪强 冯俊涛 傅俊范 傅 强 高书晶 高微微 高希武 高增贵 高智谋
宫亚军 郭建英 郭青云 郭荣君 韩宝瑜 何康来 何月秋 何自福 贺 伟 侯茂林 胡白石 胡东维
华红霞 黄凤宽 黄贵修 黄红娟 黄 建 黄世文 黄文坤 黄应昆 简桂良 简 恒 江幸福 蒋红云
蒋细良 孔垂华 雷仲仁 李宝聚 李保华 李 波 李定旭 李健强 李克斌 李明福 李世东 李世访
李湘民 李向东 李兴红 李志红 梁革梅 梁宗锁 蔺瑞明 刘 钢 刘开启 刘太国 刘西莉 刘晓辉
刘新刚 刘 勇 刘玉娣 卢向阳 陆宴辉 马春森 马占鸿 马忠华 倪汉文 倪汉祥 农向群 彭埃天
彭德良 漆艳香 秦玉川 芮昌辉 盛承发 施大钊 石 洁 束长龙 宋福平 宋玉立 孙广宇 田呈明
田国忠 汪来发 王保通 王殿轩 王恩东 王广君 王海鸿 王 琦 王 伟 王锡锋 王小艺 王晓鸣
王 勇 王源超 王振营 王中康 王忠跃 魏守辉 魏艳敏 问锦曾 吴佳教 吴钜文 吴立峰 吴茂森
吴青君 吴学宏 武予清 向 红 肖 强 肖悦岩 谢丙炎 谢 明 徐秉良 徐汉虹 徐 军 徐 梅
徐世昌 徐学农 许木启 杨艳丽 杨宇红 杨忠岐 叶 非 印丽萍 游春平 虞国跃 袁会珠 袁庆华
翟保平 张朝贤 张春霞 张 东 张广民 张桂芬 张国珍 张 杰 张 兰 张礼生 张力群 张 龙
张青叶 张星耀 张永军 张友军 赵廷昌 赵伟全 赵文军 赵文生 赵晓军 曾洪梅 郑经武 周常勇
周而勋 周国辉 周 洁 周卫川 周益林 朱荷琴 朱杰华 朱振东 竺晓平 祝增荣 (以汉语拼音排序)
Mainprogressonbiology&ecologyofColoradopotatobeetleandcountermeasuresofitsmonitoringandcontrollinginChina
Guo Wenchao1,Tuerxun1,Cheng Dengfa2,Tan Wanzhong3,Zhang Zhengkun4,Li Guoqing5,Jiang Weihua5,Deng Jianyu6,Wu Jiahe7,Deng Chunsheng8,Li Jing9,Liu Xiaoxia10Lü Heping11
(1.InstituteofPlantProtection,XinjiangAcademyofAgriculturalSciences/KeyLaboratoryofIntegratedPestManagementofCropofNorthwesternOasis,MinistryofAgriculture,P.R.China,Urumqi830091,China;2.InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences,Beijing100193,China;3.CollegeofPlantProtection,SouthwestUniversity,Chongqing400716,China; 4.InstituteofPlantProtection,JilinAcademyofAgriculturalSciences,Gongzhuling136100,China; 5.CollegeofPlantProtection,NanjingAgriculturalUniversity,Nanjing210095,China; 6.ZhejiangA&FUniversity,Lin’an311300,China;7.InstituteofMicrobiology,ChineseAcademyofSciences,Beijing100101,China; 8.InstituteofEnvironmentandSustainableDevelopmentinAgriculture,ChineseAcademyofAgriculturalSciences,Beijing100081,China;9.StationofPlantProtectionTechniquesExtensioninXinjiang,Urumqi830006,China;10.ChinaAgricultureUniversity,Beijing100193,China; 11.InstituteofPlantProtection,GansuAcademyofAgriculturalSciences,Lanzhou730070,China)
This paper reviews and analyzes the transmission situation and trend of Colorado potato beetle in all around world and China, and emphasizes the research progress about main biological and ecological characteristics of Colorado potato beetle, its occurrence status in China and integrated management.Aimed at the control situation and transmission trend of Colorado potato beetle in China, the countermeasures of monitoring and controlling have also been put forward.
Leptinotarsadecemlineata; distribution and damage; invasive biology; ecological adaptation mechanisms; integrated pest control; monitoring and control countermeasures
2013-11-06
: 2013-11-02
公益性行业(农业)科研专项(200803024,201103026)
S 435.32
: ADOI: 10.3969/j.issn.0529-1542.2014.01.001
* 通信作者6 E-mail:gwc66@163.com
